
不同水分胁迫下罗甸小米核桃生长特性研究
2015-12-20 02:51:49刘济明
中南林业调查规划 2015年1期
金 祺,刘济明
(1.贵州省林业调查规划院,贵阳 550001;2.贵州大学林学院,贵阳 550025)
不同水分胁迫下罗甸小米核桃生长特性研究
金 祺1,刘济明2
(1.贵州省林业调查规划院,贵阳 550001;2.贵州大学林学院,贵阳 550025)
以罗甸小米核桃苗木为试验材料,采用称取质量控制土壤含水量的方法,设置对照(CK)、轻度水分胁迫(W1)、中度水分胁迫(W2)和重度水分胁迫(W3)4个不同处理,研究罗甸小米核桃幼苗的生长特性。结果表明:水分胁迫下,罗甸小米核桃的生长受到抑制,苗高、地径、叶面积、生物量增长量都呈现出减少的趋势,并且随着水分胁迫强度和胁迫周期的增加而下降。
罗甸小米核桃;水分胁迫;生长特性
前言
核桃为胡桃科(Juglandaceae)胡桃属(JuglansL.)植物,在我国栽培种植已有2000多年的历史[1],原产于我国的有五个种,即普通核桃核(J.regia)、铁核桃(J.sigillata)、核桃楸(J.manshurica)、野核桃(J.cathayensis)和河北核桃(J.hopeiensis)[2]。中国是世界上最大的核桃生产国和消费国,核桃种植面积居世界首位,全国20多个省(包括自治区、直辖市)均有核桃分布[3],作为我国核桃主产区之一的贵州省地处云贵高原东南部,核桃种植面积已达3万hm2,境内核桃资源丰富,主要集中在黔西北的赫章、威宁等地[4]。
罗甸小米核桃(JuglansregiaL. f.luodianenseLiu et Xu)主要分布于贵州省罗甸县海拔1 000 m以上的北部喀斯特地区,具有缝合线浅、壳薄(手捏即开)、能取整米、果仁饱满、颜色浅、无涩味、风味香、口感好、营养丰富、出仁率和出油率高的特点。罗甸小米核桃资源量少,产量低、管理粗放,主要为庭院栽种或野生,未形成规模种植。由于其独特的经济价值,受到了当地人们的普遍欢迎,作为西南喀斯特地区适生经济树种进行栽培。本文设计主要通过测定不同程度的水分胁迫对小米核桃苗高、地径、叶面积、各部分生物量及根冠比的影响,可以为其物种资源保护、扩大种群数量及培育优质壮苗提供一定的理论与技术支持。
1 材料与方法
1.1 试验材料的准备
1.1.1 材料与工具
罗甸小米核桃种子,肥料,花盆及相关的播种工具。
1.1.2 方法
第一阶段,采用清水浸泡种子,浸泡7d后,将种子暴晒2h~4h,直到一半以上种子沿腹缝线开裂。选取缝合线开裂无病虫的种子进行苗圃地播种。
第二阶段,对播种后的小米核桃进行苗圃地管理(除草、施肥、间苗等)。
第三阶段,选取长势较好的核桃苗移栽到花盆中,移栽96盆,并进行编号,作为以后的试验用苗。然后再对这96株苗进行4个水分处理:CK正常供水 (对照);Wl轻度水分胁迫,使土壤含水量保持在FC(田间持水量)的(80±5)%;W2中度胁迫,含水量保持在FC的(60±5)%;W3重度胁迫,含水量保持在FC的(40±5)%。
在整个实验过程中通过人工称重控制土壤含水量,每天下午18:00使用电子天平称其质量并补差,保证CK,W1,W2和W3的试验梯度。
1.2 生长特性研究
1.2.1 材料与仪器
苗圃地中的试验用苗,卷尺、游标卡尺、电子天平等。
1.2.2 方法
1) 苗高、地径、生物量的测定
以10d为1周期,利用卷尺测出核桃苗木的苗高,用游标卡尺测定植株的地径(茎底部离土面5cm处茎粗),测量各部分生物量及根冠比时,需将苗木连同根系取出,洗净,用吸水纸吸干水分后将每株苗木的茎、复叶、根分别用电子天平称其鲜质量,再用烘箱烘干后称其干质量;
2) 叶面积(M)的测定
将不同处理下的植物叶片摘下分类,随机抽出6片,在A3纸上画出每片叶子的轮廓,用剪刀剪下,再将所有叶片烘干(烘干时注意将每株的6片叶子和其他叶子分开),用电子天平称出一张A3纸的质量(M1)、剪下纸张的质量(M2),烘干后的6片叶子的质量M3,烘干所有叶子的质量M总,算出6片叶子的面积S2,根据公式(1)算出6片叶片的面积,再根据公式(2)算出叶子的总面积。
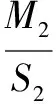
(1)
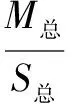
(2)
1.3 数据处理
用Excel软件和SPSS软件对数据进行处理和绘图,对所测得的原始数据进行整理分析,得出不同水分胁迫下罗甸小米核桃苗高、地径、叶面积、各部分生物量及根冠比的表格数据及曲线图,分析讨论。
2 结果与分析
2.1 水分胁迫对苗高的影响
苗高是反映植物生长状况的一项重要指标,由图1可以看出:不同水分处理对苗高产生明显影响,随着土壤持水量的减少抑制了苗木的生长,苗高的相对增长量呈现下降的趋势:CK>W1>W2>W3,苗木高度增长量最小值比对照组减少了58.28%。长期控水处理下,试验组苗高增长量与对照组苗高增长量之间的比值随着胁迫程度的加重,而逐渐减小。在水分胁迫循环周期中,随着胁迫周期的增加,苗高生长的增量下降,且下降的速度与水分胁迫强度有关。这主要是由于干旱胁迫导致细胞脱水,原生质向内收敛,细胞的延伸生长受到抑制,从而影响苗木的生长高度。
苗高的生长量在图1中呈现出两个阶段的生长,8月20日为分界线,前一阶段苗木的生长处于旺盛时期,生长量占据了整体生长量的80%,而且在CK下苗高生长量最大,说明在正常供水条件下对苗木的苗高生长有利;随着干旱胁迫的影响,苗高生长受抑制的现象越来越明显;后一阶段苗木的生长随着时间的推移逐渐减少,苗木在这个阶段生长缓慢,苗木的净光合作用的量在不断减少,说明苗木在干旱胁迫的长期作用下,生长受抑制的影响越来越严重。
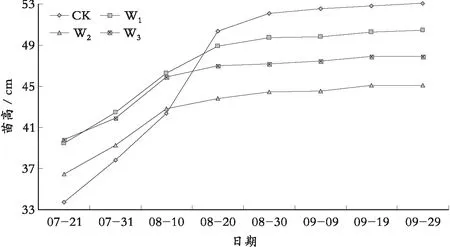
图1 不同水分胁迫下罗甸小米核桃苗高生长量
由图2得知,罗甸小米核桃苗高净增量按照不同水分胁迫梯度的不同,可以分为2段:
1) 苗木旺盛生长时期。从苗木发芽(5月份)到9月5日植物苗高净增量呈有规律的变化,这段时期,苗木净增量先增后减,构成一个生长波峰,充分体现出苗木在各个水分梯度下出现最适生长的时期;而且,苗木净增量在CK下与各个水分胁迫下存在着显著差异,在各个水分胁迫下彼此之间的差异并不明显,苗木在CK下出现最适生长峰值要比水分胁迫下出现的峰值晚。说明在苗木旺盛生长时期,苗木在充足水环境中营养生殖的最高峰值会向后推迟,水分胁迫对苗木生长的影响效果显著。
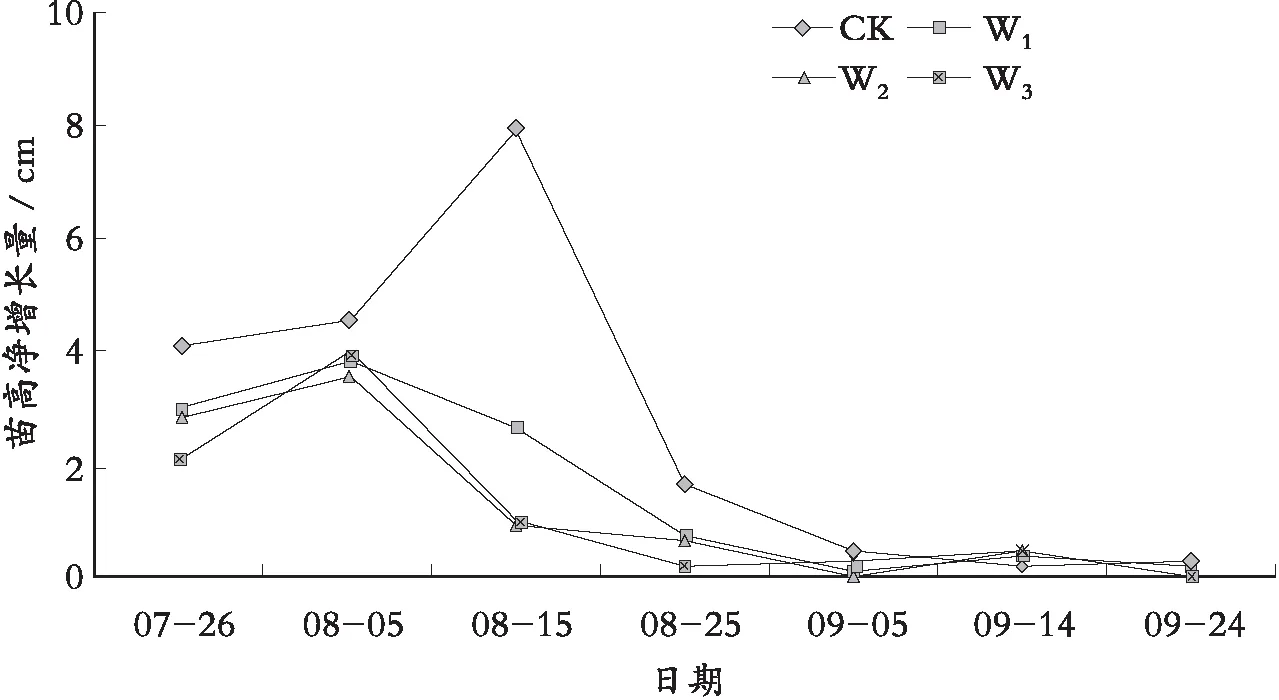
图2 水分胁迫下罗甸小米核桃苗高净增量
2) 苗木稳定生长时期。9月5日以后苗木净增量在各个水分胁迫下变化不明显,说明该时期苗木的生长已经趋于稳定。这段时期,无论是在CK下还是在各个水分胁迫下,苗木的生长都已经稳定下来,说明苗木在这段时期受到水分胁迫的影响比较小,水分胁迫不再是苗木生长抑制的主导因子,主要是因为苗木在这段时期中对水分的需求大量减少,水分胁迫下的土壤含水量小于了苗木的需求量。
2.2 水分胁迫对地径的影响
由图3可以看出,随着水分胁迫程度的增加,地径的增长量逐渐减小,茎生长速度明显减慢。CK下幼苗地径增长总量最大,W3下最小。与CK相比,W1的地径增长总量减少20.03%, W2的地径增长总量减少45.53%,W3的地径增长总量减少61.45%,在4个水分梯度下苗木的地径增长量之间的差值按照20%的梯度在减小,这恰好与试验设置的水分梯度之间的差距相一致。通过进一步探讨,可以了解苗木地径的生长量与水分胁迫梯度之间的是否存在一定的相关性。
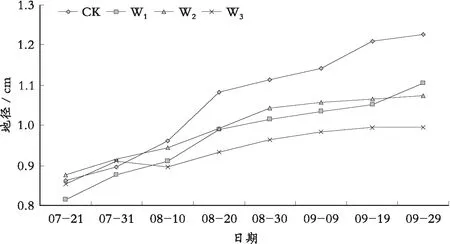
图3 不同水分胁迫下罗甸小米核桃地径生长量
对图4分析得知,随着水分胁迫程度的增加,苗木净增量之间存在如下关系:CK>W1>W2>W3。即苗木净增量的大小与苗木受水分胁迫的程度是成正比的,当胁迫越大时,地径净增量越小,在CK下地径净增量相应地表现出增量的最大值。而且在试验阶段,苗木的地径净增量呈“双峰型”增长。

图4 不同水分胁迫下罗甸小米核桃地径净增量
2.3 水分胁迫对叶面积的影响
由图5可以看出:水分亏缺处理下的罗甸小米核桃幼苗叶面积均表现为下降趋势,与CK组相比,W1下降了9.71%,W2下降了40.92%,W3下降了47.92%,下降幅度与胁迫强度大小有关,水分胁迫程度越强,叶面积减小的数值也就越大,这主要是因为水分胁迫下,叶片是植物外部形态中反应最敏感的器官,叶片的主要变化有利于保水和提高水分利用率,随胁迫程度的加强,细胞的扩大和分裂受限制,细胞增大比细胞分裂对水分缺乏更为敏感。一旦缺水,首先受抑制的是细胞增大,单叶叶面积比正常条件下形成的要小,从而减少水分向大气散失。植物通过保持一个较小的总叶面积来使单位叶面积的光合作用维持在一定水平是植物适应土壤水分减少的一种重要方式。
2.4 水分胁迫对各部分生长量及根冠比的影响
生物量是植物获取能量能力的主要体现,对植物的发育形成具有十分重要的影响。众多研究表明:生物量的大小,综合反映了外界因子对林木生长的影响以及林木对外界环境的适应能力。在生长发育中,植物总是不断调整其生长和生物量的分配策略来适应环境变化。在土壤水分胁迫条件下,植物通过调整生物量分配将逆境伤害降低到最小来适应环境胁迫。
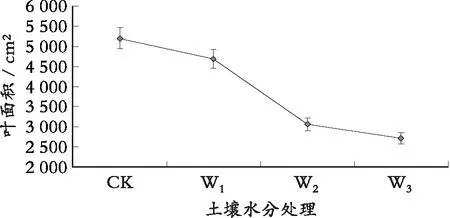
图5 不同水分胁迫下罗甸小米核桃的叶面积
2.4.1 对地上部分生物量的影响
在土壤水分亏缺时,树木根系的水分和养分吸收受到影响,使各个器官的生长发育都受到限制。首先受到抑制的是细胞增大,造成光合面积减小;其次是细胞增殖,进而造成植物生物量的减少。在土壤水分胁迫条件下,植物通过减少茎、叶干的质量,使地上部分生物量减少从而避免有限水分在地上部分的消耗,以适应在水分胁迫环境下的生存。
2.4.2 对地下部分生物量的影响
根干质量是分析根系生长最常用的指标之一,它反映根系生长和环境的关系。不同的水分胁迫对幼苗生长地下生物量都有不同程度的影响,这是因为植物在受到水分胁迫时,会更多地把生物量分配到根系,以便更好地吸收土壤里的水分和养分。但重度水分胁迫会使根系的生长减慢,可能是由于在重度水分胁迫条件下,植物各个器官的生长都受到了抑制,干物质量积累减小,地下生物量也随之下降。
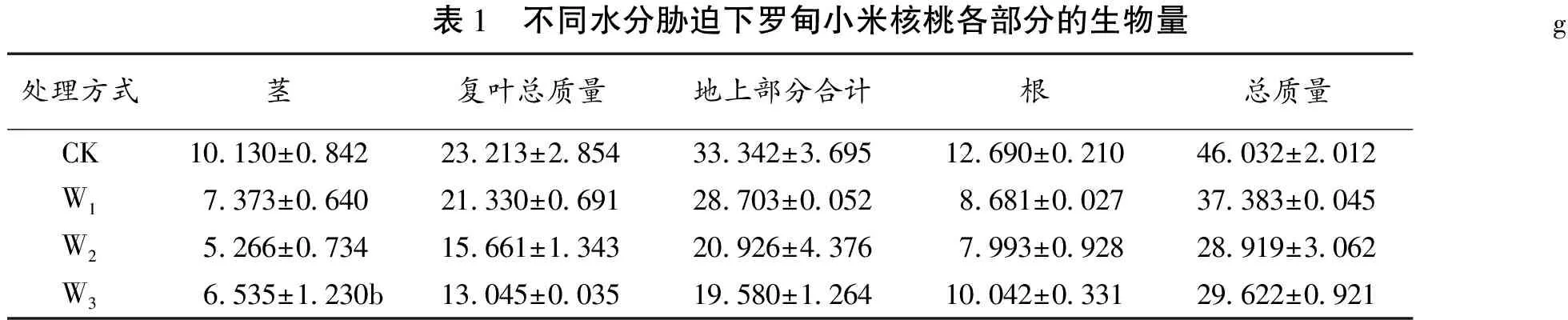
表1 不同水分胁迫下罗甸小米核桃各部分的生物量g处理方式茎复叶总质量地上部分合计根总质量CK10 130±0 84223 213±2 85433 342±3 69512 690±0 21046 032±2 012W17 373±0 64021 330±0 69128 703±0 0528 681±0 02737 383±0 045W25 266±0 73415 661±1 34320 926±4 3767 993±0 92828 919±3 062W36 535±1 230b13 045±0 03519 580±1 26410 042±0 33129 622±0 921
研究表明,对罗甸小米核桃不同水分胁迫下幼苗生物量分析,苗木生物量对干旱胁迫的反应基本趋势一致,即苗木的生物量随土壤含水量的降低而下降。
据表1可知:①在茎的生物量中,CK与W1,W2,W3之间差异极显著;而W1与W3,W2与W3之间差异不显著;W1与W2差异显著。②地上部分生物量:CK与W1,W2与W3之间差异不显著,CK与W2和W3差异都极显著,W1与W2之间差异显著,W1与W3之间差异极显著。③根的生物量中,CK与W1,W2,W3之间差异极显著,W1与W2差异不显著,W2与W3差异极显著。
从器官生物量分配看,表1显示不同的水分胁迫强度苗木根、茎、叶分别占总生物量的比例上都有很大影响。从器官生物量分配看,图显示不同的水分胁迫强度罗甸小米核桃苗木根、茎、叶分别占总生物量的比例上都有很大影响。并且可以明显看出植物在水分胁迫加强的情况下把更多的能量用于地下部分的生长,水分胁迫抑制了罗甸小米核桃苗木总生物量,促进了根生物量的分配比例。这说明在水分亏缺的条件下,幼苗把更多的能量用于地下部分生长,获取更多的土壤水分和养料以适应土壤水分的不足。
在正常水分条件下,植物地上部分与地下部分生长比例基本相似[5],但在胁迫逆境条件下,植物生物量分配的改变有助于树木适应环境的变化,根系的生物量是植物适应干旱的重要标志[6]。根系是树木生长最重要的地下营养器官,当植物受到干旱胁迫时,根系生物量在整个植物生物量中所占比重提高,有助于植物从干旱的土壤中吸收水分,保持体内水分平衡。因此,在水分胁迫下,根冠比是鉴定植物抗旱性的重要指标[7]。根冠比不仅反映植物对光合产物的分配对策,也反映了植物对水分环境的适应对策。
根系在植物的生理过程中,有着非常重要的地位,有吸收水分、矿质营养、固持植株等作用。适应干旱的植物根系发达而深扎,根冠比大能更有效的利用土壤水分特别是土壤深处的水分,并能保持水分平衡。当土壤含水量降低时,会增加根的相对重量,根冠比值增高。潘瑞炽等[8]对几个农作物的抗旱性研究发现,根冠比越大越抗旱,否则不抗旱。
从图6中可以看出:CK,W1,W2的根冠比比值差异都不太明显,但重度水分胁迫组根冠比比值与其它组有显著的差异,说明当水分胁迫达到重度时,对幼苗生物量的分配才有影响,且影响明显。生物量的分配比例在一定程度上能反映植物在受到水分胁迫时的生存对策,植物在地上部分生物量降低的同时,较多地提高根的比重,即地下生物量,这有利于缓解植物在水分胁迫下水分、养分的供求矛盾。所以这种变化也可以认为是罗甸小米核桃幼苗在重度水分胁迫下的一种响应对策。

图6 不同水分胁迫下罗甸小米核桃根冠比
3 结论
通过对水分胁迫下罗甸小米核桃生长特性研究,以及对数据的分析处理,得出不同水分处理对罗甸小米核桃生长和生理都产生了显著的影响:
1) 不同水分处理对苗高、地径产生明显影响,在不同的水分胁迫下,苗高、地径的相对增长量之间存在如下关系:CK>W1>W2>W3,随着胁迫强度的增加对苗高、地径的影响更加显著。说明在正常供水条件下对苗木的苗高生长有利,随着干旱胁迫的影响加剧和影响时间的延续,苗高生长受抑制的表征现象更明显;同时,在不同水分胁迫下苗木的苗高、地径的增长量都保持着一致的生长,水分对苗木生长特性各指标的影响是整体性的,水分含量下降会直接导致苗木生长量的下降。
2) 在旺盛生长时期,苗高净增量在CK下与各个水分胁迫下存在着显著差异,但在各个水分胁迫下彼此之间的差异并不明显,苗木在CK下出现最适生长峰值要比水分胁迫下出现的峰值晚;苗木在稳定生长器水分胁迫不再是抑制苗木生长的主导因子;在地径净增量的大小与苗木受水分胁迫的程度是成反比的,当胁迫越大时,地径净增量越小,在CK下地径净增量相应地表现出增量的最大值,地径净增量呈“双峰型”增长。
3) 水分亏缺处理下的罗甸小米核桃幼苗叶面积均表现为下降趋势,下降幅度与胁迫强度大小有关,水分胁迫程度越强,叶面积减小的数值也就越大。苗木在水分胁迫下,由于土壤含水量不高,苗木根系所能吸收利用于蒸腾作用的水分不多,为了减少苗木的蒸腾作用,生长过程中,苗木叶片面积减小,但是同时也就降低了苗木的有效光合叶面积,影响了苗木整体的生长量,使得植株地上部分生长矮小。
4) 水分胁迫抑制了罗甸小米核桃苗木总生物量,促进了根生物量的分配比例。在水分亏缺的条件下,幼苗把更多的能量用于地下部分生长,有利于从土壤中获取更多的土壤水分和养料以满足植物地上部分蒸腾作用所需水分和植物生长所需要的能量和物质,加大苗木对逆境生长的适应。
5) 罗甸小米核桃苗木的生物量随土壤含水量的降低而下降。水分胁迫对根冠比产生一定的影响,在一定程度范围内,对苗木各部分生长量的影响不明显,当达到W3(重度)水分胁迫时,根冠比的比值越大,苗木各部分生长量之间存在着显著差异,而且不利于苗木的正常生长,随着时间的推移,该水分胁迫不能够及时解除,一旦超过了苗木自身的耐性范围,苗木开始发生病变或者死亡。
[1]甘霖,吴正琴. 核桃的发展及其应注重的问题[J]. 四川果树,1995(1):30-32.
[2]郗荣庭,张毅萍. 中国果树志:核桃卷[M]. 北京:中国林业出版社,1996.
[3]魏贞伟,沈茂盛. 核桃开发利用研究[J]. 现代农业科技,2009,20:149-150.
[4]潘学军,张文娥,刘伟,等. 贵州核桃种仁脂肪酸和氨基酸含量分析[J]. 西南农业学报,2010(2):407-501.
[5]Wilson J B. Shoot Competition and Root Competition[J]. J Appl Ecol,1988,25(2):279-296.
[6]Theodore C. Hsiao,Liu Kangxu. Sensitivity of growth of roots versus leaves to water stress: biophysical analysis and relation to water transport[J]. Journal of Experimental Botany ,2000,51(350) :1595-1616.
[7]杨敏生,梁海永,王进茂,等. 水分胁迫下白杨双交杂种无性系苗木生长研究[J]. 河北农业大学学报,2002,25(4):2-6.
[8]潘瑞炽. 植物生理学:第六版[M]. 北京:高等教育出版社,2008.
[9]汤章城. 植物对水分胁迫的反应和适应性:I抗逆性的一般概念和植物的抗涝性[J]. 植物生理学通讯,1983,19(3):24-29.
[10]汤章城.植物对水分胁迫的反应和适应性:Ⅱ植物对干旱的反应和适应性[J].植物生理学通讯,1983,19(4):1-7.
StudyonGrowthCharacteristicsofJuglansregiaL.f.luodianenseLiuetXuunderDifferentWaterStress
JIN Qi1, LIU Jiming2
(1.Guizhou Forestry Survey Institute,Guiyang 550001,Guizhou,China;2.College of Foresty,Guizhou University,Guiyang 550025,Guizhou,China)
Took the seedlings ofJuglansregiaL. f.luodianenseLiu et Xu as experiment materials and weighing method was uesd to control the soil moisture content. The indexes of growth characteristics ofJuglansregiaL. f.luodianenseLiu et Xu seedlings were analyzed under different water stress treatments by setting the control(CK), mild water stress (W1), moderate water stress (W2) and severe water stress (W3), 4 different treatment total. The result showed that: The growth ofJ.regiaL. f.luodianenseLiu et Xu were inhibited under the water stress, Seedling height, ground diameter, leaf area and biomass growth presented a decrease trend, and those figures reduced with the increase of water stress strength and stress cycle decreation.
JuglansregiaL. f.luodianenseLiu et Xu;water stress;growth characteristics
2014—12—01
2014—12—10
贵州省林业厅重大项目“喀斯特地区特色经济林树种培育与示范”(黔林科合[2010]重大04号)。
金祺(1974—),男,工程师,主要从事林业调查与规划工作。
刘济明(1963—),男,博士,教授,主要从事植物生态学研究。
S 664.1
A
1003—6075(2015)01—0048—06
10.16166/j.cnki.cn43—1095.2015.01.013
猜你喜欢
大众科学(2021年9期)2021-12-23 01:38:42
今日农业(2021年19期)2021-11-27 00:45:49
农技服务(2020年1期)2020-12-17 08:15:01
山东农业科学(2019年11期)2019-12-24 01:11:27
现代农业科技(2019年1期)2019-07-11 01:36:25
基层中医药(2018年2期)2018-05-31 08:45:17
现代农业科技(2017年24期)2018-01-22 21:35:06
乡村地理(2017年4期)2017-09-18 02:54:20
贵州地质(2016年4期)2016-02-20 01:47:04
当代贵州(2015年22期)2015-11-29 02:58:37
