植物雌激素对骨组织的影响及作用机制的研究进展
2015-12-16 07:44:26张晓梅施雪妮何学令
四川解剖学杂志 2015年4期
张晓梅 彭 旭 施雪妮 何学令 刘 艳△
1(四川大学 实验动物中心,成都610041)
2(四川大学 生命科学院,成都610041)
雌激素在骨生长、矿化和细胞的分化及功能活性中发挥着重要作用。因此,在很长一段时间里,对于绝经后骨质疏松症的治疗,临床上推荐补充外源性雌激素的方法,即传统的雌激素替代疗(estrogen replacement treatment,ERT)。但是大量临床实验的结果表明[1-3],雌激素替代疗法有使乳腺癌和子宫内膜癌的发生概率上升的趋势,增加静脉血栓的发生率等。近年来,临床上广泛采用选择性雌激素受体调节剂(selective estrogen regulate mediator,SERM),如他莫西芬和雷洛西芬,虽然在治疗骨质疏松的过程中达到了预期的治疗效果[5,6],但显示均有潜在副作用,他莫西芬由于其对子宫的雌激素效应可能会使子宫内膜癌的发病风险增加,而雷洛西芬近期也有报道其可能增加热潮红、深静脉血栓、肺栓塞等类似使用ERT后带来的副作用[4-7]。因此,寻找更加安全的抗骨质疏松的药物成为了被广泛关注的课题。已有许多研究等发现[8-10]某些植物雌激素具有防治骨质疏松的作用。植物雌激素是一类天然存在于植物中具有杂环多酚类的结构的生物活性物质。至上世纪四十年代首次[11]在绵羊体内观察到某些化合物产生雌激素样效应以来,至今已经发现植物雌激素百余种。这些具有杂环多酚结构的植物雌激素大都存在于各种谷物、豆类中。近年来在一些升麻科植物,苜蓿类植物以及真菌类中也提取出植物雌激素。目前,研究的植物雌激素主要为4类:(1)异黄酮类(Isoflavones):包 括 染 料 木 黄 酮 (genistein)、大 豆 黄 酮(daidzein)、黄豆黄素(glycitein);(2)木酚素类(Lignan);(3)香豆素类(Coumestans);(4)真菌雌激素类:如玉米赤霉烯酮(Zearalenone)。
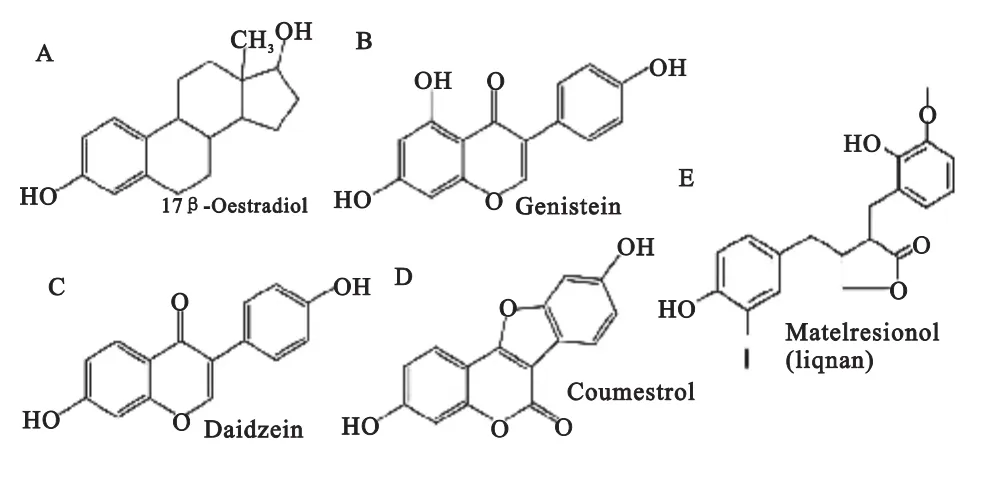
图1 17β-雌二醇与植物雌激素结构示意图
1 植物雌激素的作用机制
雌激素的经典作用机制认为,雌激素首先与雌激素受体(estrogenreceptor,ER)上的配体结合区(ligand binding domain,LBD)结合并使受体二聚化,进一步雌激素受体的DNA结合区(DNA binding domain,DBD)与靶基因启动子上的雌激素应答元件(estrogen response element,ERE)结合,从而启动靶基因的转录。植物雌激素与内源性雌激素结构相似,均具有酚环结构。(如图1所示)因此与雌激素的两型受体均可结合,但与受体的亲和力相对内源性雌激素较弱[12,13]。一些植物雌激素既能模拟内源性雌激素的作用而发挥雌激素样的效应,同时又可以阻断与雌激素受体的结合产生雌激素拮抗的作用。Kurzer[14]研究发现植物雌激素能够与ER结合,刺激表达ER的乳腺癌细胞生长,诱导生成特殊的雌激素反应基因。雌激素与ERα和ERβ亲和力相似,而一些植物雌激素尤其是异黄酮类对ERβ的亲和力远高于ERα[15]。目前对其作用机制还未完全阐明。植物雌激素也可以通过不与ER作用的其他机制来作用于组织器官,如:诱导癌细胞的分化、抑制酪氨酸激酶和DNA拓扑异构酶的活性等。植物雌激素还在细胞和分子水平影响生物合成以及激素相关的信号转导等。同时由于雌激素受体在机体内组织分布不同,分布的比例也不同等,使得植物雌激素对不同组织的作用机制以及产生的生物效应都不竟相同。
2 异黄酮类植物雌激素
异黄酮类是目前研究得较多的一类植物雌激素,因为其分子结构与雌激素相似,能激活ER,能发挥对骨组织的保护作用,但其较雌激素具有更高的ERβ亲和性,与ERα的结合时间长度及亲和性则不及雌激素,因此在保护骨组织的同时,异黄酮对子宫和乳腺的影响较小,减少患子宫内膜癌和乳腺癌的风险。目前已有产品应用于保健和临床。它主要包括染料木黄酮(genistein)、大豆黄酮(daidzein)和大豆黄素(glycitein)以及其相应的葡萄糖配糖体六类。目前在研究异黄酮类对防治骨质疏松的作用的研究中,genistein和daidzein是研究得较为深入的两种植物雌激素。在利用骨质疏松动物疾病模型进行研究发现异黄酮类对去卵巢骨质疏松大鼠产生显著的骨保护的作用,它不仅能抑制由于雌激素缺乏后引起的高骨转换,而且能够影响骨微结构[16],而在使用高剂量的异黄酮时能增加去卵巢大鼠松质骨和皮质骨的骨矿物质密度(bone mineral density,BMD)[17]。利用体外培养细胞的研究发现异黄酮能使成骨样细胞增殖,ALP活性和胶原合成增强(如下图2),同时也能影响破骨细胞的磷酸化[18]。近来更有研究发现[19],在核因子NF-кB活化因子受体配体(RANKL)诱导的破骨细胞分化中,genistein能减少破骨细胞的多核化及多核细胞核的数量,并能抑制细胞周期的S期,增加G0,G1和G2,M期。这提示异黄酮类植物雌激素可能直接影响成骨细胞的分化和功能活性,而抑制破骨细胞的形成和功能活性。在流行病学的研究中发现在亚洲妇女中,食用高含量大豆蛋白食物的妇女具有较高的BMD,反之其BMD则较低:在42~52岁绝经期间的妇女中,日本妇女genistein较中国妇女的摄入量大;对于日本妇女绝经前期BMD与genistein的摄入量呈剂量性的正性关系,而对于绝经后的中国妇女和日本妇女,BMD与genistein的摄入量无相关性[20]。Lydeking等[21]给177名绝经后妇女每日服用500毫升豆奶(含有异黄酮76毫克),两年后其腰椎的骨密度和骨矿物质含量(bone mineral content,BMC)各增长1.1%和2.0% ,而每日异黄酮摄入量小于1毫克对照组的妇女其椎体的BMD和BMC各下降4.2%和4.3%。在该试验的中期则不能观测到椎体的BMD和BMC改变。还有部分的研究显示[22]服用异黄酮60mg可改变体内骨转化标志物浓度而并不影响BMD和BMC。在对异黄酮类植物雌激素的研究中,由于用药时间、用药剂量的不同,因此观测到的对BMD的影响并不一致,也很难确定其在防治骨质疏松疗效方面的有效信息,需要进一步的长期临床研究证明异黄酮类植物雌激素的抗骨质疏松的作用。

图2 异黄酮对成骨样细胞增殖、ALP分泌和胶原合成的影响Fig.2 The effect of isoflavones on osteoblast-like cell、ALP and collagen
在成骨细胞、破骨细胞以及骨髓基质细胞等上均存在有ERα和ERβ,而且它们在松质骨与皮质骨上的分布比例不一。虽然异黄酮类植物雌激素较ERα来说,与ERβ具有更高的亲和性,但目前异黄酮类的植物雌激素如genistein、daidzein对骨组织保护作用的ER介导机制还并未完全的清楚。而异黄酮类影响成骨细胞和破骨细胞的分化以及功能活性的影响除了通过相似于内源性雌激素能够于ER结合的途径外,还可通过局部细胞因子和旁分泌的激素的介导作用。TNF-α、IL-1β和IL-6是成骨细胞或骨髓基质细胞分泌的细胞因子,TNF-α能刺激成骨细胞分泌其他因子IL-1β和IL-6等,对调节破骨细胞的增殖。分化等功能活动产生重要的影响。genistein、daidzein等可调节一些炎症因子如TNF-α或IL-1β的水平影响破骨细胞的活性同时抑制TNF-α引起的细胞调亡[23-25]。genistein可以调节骨代谢相关基因如骨基质蛋白,细胞生长因子等的表达[26]。异黄酮对减少骨吸收、促进骨形成的机制是多方面的,对与其保护骨组织的作用机制仍需更深入的研究。
3 木酚类植物雌激素
木酚素(Lignan)是一类主要通过p-羟基苯乙烯单体的氧化耦合而成的,在数百种植物的木质部、根、叶、花和果实中均发现有此类物质,同时在动物的体液(血清、血浆和尿液)中也发现有该类物质,它们主要是植物木酚素在肠道细菌的作用下的代谢产物。因此根据它们的来源就分为植物木酚素和哺乳动物木酚素。当内源性的雌激素减少时某些哺乳动物木酚素呈现雌激素样效应,反之则显现抗雌激素样效应[27]。因此木酚素在对雌激素缺乏后产生的相关疾病可产生一定的作用。木酚素种类繁多,虽然对其在骨组织影响的研究相对异黄酮类较少,但仍然发现Sauchinone(木酚素类的一种)能够影响RANK/RANKL途径抑制破骨细胞的生成[28]。异紫杉脂素 (Isotaxiresinol)能够使去卵巢大鼠BMD和BMC增加,并能影响骨转换标志物,提示其可能对防治绝经后骨质疏松产生一定的作用[29]。木酚素类化合物在植物的成分中含量较少,但却广泛存在很多植物中,尤其使亚麻籽。目前,我们已经发现木酚素类植物雌激素还具有一定的抗氧化活性和抗癌活性,但是人们对木酚素的了解和关注远远不如异黄酮类,对其安全性问题、体内作用剂量和作用机制等仍需要更深入的研究和探讨。
4 香豆素类植物雌激素
香豆素类植物雌激素(Coumestans)是另一类具有雌激素样效应的植物化合物,它最初是从三叶草中提取出来的。Coumestans被证实和genistein一样与ERβ具有较高的亲和性,而且coumestans对子宫不产生明显的雌激素效应。因此coumestans被考虑用于防治骨质疏松的研究。Tsutusmi等[30]报道,体外实验发现拟雌内酯对骨吸收激动剂的骨吸收作用具有抑制作用,能加强鸡胚股骨骨质的矿化。因此,拟雌内酯是一种独特的,既能抑制骨吸收,又能刺激骨质矿化,而对大鼠子宫的雌激素样作用很弱,Drpaer等[31]研究了拟雌内酯对卵巢切除骨质疏松大鼠骨丢失的情况,结果发现拟雌内酯组与雌二醇组在抑制股骨骨密度降低的作用上无统计学差别。同时还发现尿钙的变化与骨密度的变化并不一致,拟雌内酯能降低尿钙/肌酐比值,降低尿钙排泄率,由于尿钙仅占骨钙丢失的一小部分,可能拟雌内酯会影响其他钙代谢的平衡。Morris等[32]发现在拟雌内酯处理的雌激素缺乏大鼠,尿中吡啉啶和脱氧吡啉啶含量减少,说明拟雌内酯可抑制骨的吸收。以上研究表明拟雌内酯能刺激骨质矿化,抑制骨吸收,它还可通过其他机制影响骨钙的代谢,从而抑制骨的吸收。但其对骨组织的影响机制仍很不了解,可能不通过ER结合而发挥作用,而是通过ERK1/2通道RANKL降低诱导的TRAP的活性来影响破骨细胞功能,并在抑制破骨细胞的分化起者重要作用。
5 真菌类植物雌激素
1962年Stob等[33]首先报道自玉米赤霉的培养基中分离出具有雌激素样效应的物质,因此这种物质被称为玉米赤霉烯酮(Zearalenone,ZEN)。1966年 Urry等[34,35]利用经典化学、核磁共振及质谱技术,确定此物质的化学结构属于二羟基苯酸内酯类,是自然界罕见的一类化合物。由于ZEN也能促进某些真菌形成子实体,因此也被认为是真菌的性激素。ZEN的衍生物主要有以下五种α-zearalenol(α-ZEA),β-zearalenol (β-ZEA),α-zearalanol (α-ZAL),β-zearalanol(β-ZAL),zearalanone(ZAN)(如图3)。由于ZEN存在如小麦、高粱等很多粮食谷物中,国外的研究认为ZEN是动物吃了大量发霉玉米引起雌性化疾病的病因,所以在种子和食品的检疫工作中,也将其作为污染的指标,而ZEN的还原产物在霉变的一些谷物或者育种中也同样被发现[36.37]。因此对ZEN的研究主要是探讨其对人体和肉用牛羊等产生的生殖系统的毒性、免疫毒性及遗传毒性等有害影响。而另一方面英美等国科学家已做了大量实验证明ZEN及其衍生物具有二羟基苯酸内酯类的结构,使其能够通过改变构象与哺乳动物体内的ER结合,并可促进肉用牛羊增重而无可察觉的副作用,尤其是其还原产物α-ZAL,可促进肉用牛羊的蛋白质合成因而增加瘦肉重量,与动物雌激素相比,作为动物生长促进剂,二者所用剂量相似,安全无毒,体内无积蓄,不引起组织增生,优于雌激素[38,39]目前,美国、加拿大等国已作为动物生长促进剂用作动物饲料,商品名为Ralgro。1980年起我国学者证实ZEN为冬小麦春化的关键物质,为植物中普遍存在的内源激素,而且在许多植物如大豆、冬小麦等发现ZEN且随着生长发育的不同阶段发生动态变化,这些植物是人们经常食用的,并非“污染物”。同样在冬性植物的越冬茎尖与花菜内也发现了ZEN的同系物,其中就有α-ZAL,这提示了它的安全性,而且研究显示[40]。

图3 ZEA及其衍生物的化学结构Fig.3 Chemical structures of ZEA and its derivatives
α-ZAL的生物活性是ZEN的18倍。这就提示了我们对α-ZAL应用前景的探讨。戴顺龄等人通过大量的实验发现α-ZAL有明显抗动脉粥样硬化作用,可明显的抑制主动脉粥样斑块形成,可调节血管活性因子和抗氧化损伤等发挥保护心血管的作用,而副作用比雌激素明显减少,并不引起组织的增生,对正常乳腺细胞不产生增殖的影响[38,39,41-43]。我们看到α-ZAL已经在心血管方面初步显现其有利的影响,而目前在α-ZAL对骨组织的影响方面的研究相对较少。仅有少数的实验报道发现α-ZAL可以使去卵巢大鼠的BMD和BMC增加[31,44];可以增加去势雄性羊羔的BMC[45]。这些研究提示α-ZAL有一定的骨保护的作用,可能对防治绝经后骨质疏松产生潜在的临床意义。
6 问题与展望
综上所述,由于植物雌激素广泛存在于人类饮食中,具有相似于内源性雌激素的效应,因此,植物雌激素被认为在研究抗骨质疏松药物上是很有应用前景的,也使得越来越多的学者关注它。现在已有越来越多的植物雌激素被发现,如在葛根中提取的葛根素,在刺桐、亚麻籽、蛇麻花等植物中均能提取出植物雌激素,而且研究也发现它们中的一些如葛根素能改善去卵巢大鼠的BMD,影响其骨微结构和骨转换指标的变化,具有一定的骨保护功能[46]。大蒜被认为在降低血中胆固醇有很好的治疗效果,而最近有研究者认为其中也含有植物雌激素,并认为其可能通过增加钙的吸收而减少骨矿物质量的丢失[45]。而且,已经从大量的流行病学的调查、动物实验中发现植物雌激素确实对绝经后骨质疏松具有一定的防治作用,同时不产生ERT引起的对子宫等的副作用。虽然已经有一些植物雌激素的产品应用于保健和临床,但由于植物雌激素种类很多,其结构和功能的差异很大,目前对植物雌激素的作用机制还不是很清楚,在评价其安全性、对植物雌激素反应的剂量差异等问题上仍然需要深入的研究。
[1] Writing Group for the Women’s Health Initiative Investigators.Risksand benefits of estrogen plus progestin in healthy postmenopasual women:principal results from the Women’s Health Initiative randomized controlled trial[J].JAMA ,2002,288(3):321-333.
[2] Lacey JV,Mink PJ,Lubin JH,et al.Menopausal hormone replacement therapy and the risk of ovarian cancer[J].JAMA,2002,288(4):334-341.
[3] Wuttke W,Jarry H,Westphalen S,et al.Phytoestrogens for hormone replacement therapy?[J].Steroid Biochem Mol Biol,2002,83:133-147.
[4] Jordan VC.The past,present,and future of selective estrogen receptor modulation[J].Ann NY Acad Sci,2001,949:72-79.
[5] Pukkala E,Kyyrönen P,Sankila R,et al.Tamoxifen and toremifene treatment of breast cancer and risk of subsequent endometrial cancer:apopulation-based case-control study.Int J Cancer,2002,100:337-341.
[6] Arshad M,Sengupta S,Sharma S,etal.In vitro anti-resorptive activity and prevention of ovariectomy-induced osteoporosis in female Sprague-Dawley rats by ormeloxifene,a selective estrogen receptor modulator[J].Journal of Steroid Biochemistry & Molecular Biology,2004,91:67-78.
[7] Clemett D,Spencer CM.Raloxifene and post-menopausal osteoporosis[J].Drugs,2000,60:380-409.
[8] Arjmandi BH,Alekel L,Hollis BW,et al.Dietary soybean protein prevents bone loss in an ovariectomized rat model ofosteoporosis[J].J Nutr,1996,126:161-167.
[9] Draper CR,Edel MJ,Dick IM,etal.Phytoestrogens reduce bone loss and bone resorption in oophorectomized rats[J].J Nutr,1997,127,1795-1799.
[10] Morabito N,Crisafulli A,Vergara C,et al.Effectsof genistein and hormone-replacement therapy on bone loss in early postmenopausal women:a randomized double-blindplacebo-controlled study[J].J Bone Miner Res,2002,17:1904-1912.
[11] Bennetts HW,Underwood EJ,Shier FL.A specific breeding problem of sheep on subterranean clover pastures in Western Australia[J].Aust Vet J,1946,22:2-12.
[12] Martin PM,Horwitz KB,Ryan DS,et al.Phytoestrogens interaction with estrogen receptors in human breast cancer cells[J].Endocrinology,1978,103:1860-1867.
[13] Le Bail JC,Varnat F,Nicolas JC,Habrioux G.Estrogenic and antiproliferative activities on MCF-7humanbreast cancer cells by flavonoids[J].Cancer Lett,1998,130:209-216.
[14] Kurzer MS.Hormonal effects of soy in premenopausal women and men[J].J Nutr,2002,132:570S-573S.
[15] Kuiper GG,Lemmen JG,Carlsson B,et a l.Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta[J].Endocrinology ,1998,139:4252-4263.
[16] Latha Devareddy,Dania A.Khalil,Brenda J.Smith,et al.Arjmandi Soy moderately improves microstructural properties without affecting bone mass in an ovariectomized rat model of osteoporosis[J],Bone,2006,38:686-693.
[17] Chang Sun Hwang,Ho Seok Kwak,Hwa Jae Lim,et al.Isoflavone metabolites and their in vitro dual functions:They can act as an estrogenic agonist or antagonist depending on the estrogen concentration[J].Journal of Steroid Biochemistry &Molecular Biology,2006,101:246-253.
[18] Choi EM,Suh KS,Kim YS,et al.Soybean ethanol extract increases the function of osteoblastic MC3T3-E1cells[J].Phytochemistry 2001,56:733-739.
[19] Garcia PalaciosV,Robinson LJ,Borysenko CW,et al.Negative reg ulation of RANKL-induced osteoclastie differentiation in RAW264.7cells by estrogen and phytoestrogen[J].J Biol Chem,2005,280(14):13720-13727.
[20] Lorraine A.Fitzpatrick.Soy isoflavones:hope or hype?[J]Maturitas,2003,1:S21-S29.
[21] Lydeking E,Jensen JBE,Setchell KDR,et al.Soymilk orp rogesterone for prevents of bone loss a 2year randomized,placebo-controlled trial[J].Eur J Nutr,2004,43(4):246-257.
[22] Arjmandi BH,Lucas EA,Khalil DA,et al.One year soy proteinsupp lementation has positive effects on bone formation markers but not bone density in postmenopausal women[J].Nutr J,2005,4(1):8.
[23] P.Fanti,M.C.Monier-Faugere,Z.Geng,J.Schmidt,P.E.Morris,D.Cohen and H.H.Malluche.The Phytoestrogen Genistein Reduces Bone Loss in Short-Term Ovariectomized Rats[J].Osteoporos Int,1998,8:274-281.
[24] Sik Suh K,Koh G,Young Park C,et al.Soybean isoflavones inhibit tumor necrosis factor-Alpha-induced apoptosis and the production of interleukin-6and prostaglandin E(2)in osteoblastic cells[J].Phytochemistry,2003,63:209-215.
[25] Kwang Sik Suha,Gwanpyo Koha,Cheol Young Parka,et al.Soybean isoflavones inhibit tumor necrosis factor-a-inducedapoptosis and the production of interleukin-6and prostaglandin E2in osteoblastic cells[J].Phytochemistry,2003,(63):209-215.
[26] Jae-Eun Piea,Jin-Hee Parkb,Yoon-Hee Parkb,et al.Effect of genistein on the expression of bone metabolism genes in ovariectomized mice using a cDNA microarray[J].Journal of Nu-tritional Biochemistry,2006,17:157-164.
[27] 李欣,袁建平,刘昕,王江海.木脂素--一类重要的天然植物雌激素[J].中国中药杂志,2006,24(31):2021-2025.
[28] Kyoung-Youn Han,Daum Yang,Eun-Ju Chang,et al.Inhibition of osteoclast differentiation and bone resorption by sauchinone[J].biochemical pharmacology,2007,74:911-923.
[29] Yina J,Tezukaa Y,Subehana L,et al.In vivo anti-osteoporotic activity of isotaxiresinol,alignan from wood of Taxus yunnanensis[J].Phytomedicine,2006,13:37-42.
[30] Tsutsumi N,Effect of coumestrol on bone metabolism in organ culture.Biol Pharm Bull,1995;18(7):1012-1015.
[31] Draper CR,Edel MJ,Dick IM,et al.Phytoestrogens reduce bone loss and bone resorption in oophorectomized rats[J].J Nutr,1997,127(9):1795-1799.
[32] Morris HA,O’Loughlin PD,Mason RA,et al.The effect of oophorectmy on calcium homeostasis[J].Bone,1995;17:169S-174S.
[33] Stob M,Boldwin RS,Tuite J,et al.Isolation of an anabolic uterotrophic compound from corn infected with Gibbezella Zeae[J].Nature,1962,196(1):318.
[34] Chriotense CM,Nelson GH,Mirocha CJ.Effect on the white rat uterus of a toxsis substance from Fusarium [J].Microbiol Appl,1966,13:653-659.
[35] Urry WH,Wehrmeister HL,Hodge EB,et al.The structure of Zearalenone[J].Tetrahedon letter,1966,27(3):109-114.
[36] Bottalico A,Visconti A,Logrieco A,et al.Occurrence of zearalenols(diastereomeric mixture)in corn stalkrot and their production by associated Fusarium species[J].Appl Environ Microbiol,1985,49,547-551.
[37] Richardson KE, Hagler WM, Mirocha CJ.Production ofzearalenone a-and b-zearalenol and a-and b-zearalanol by Fusarium spp.in rice culture[J].J Agric Food Chem,1985,33:862-866.
[38] 王雯,杨慧,王红霞,张立克.17β-雌二醇及植物雌激素α-玉米赤霉醇致去卵巢大鼠子宫增大作用的比较[J].首都医科大学学报,2005,1(26):51-54.
[39] 邓文慧,戴顺龄,等.植物雌激素α-玉米赤霉醇对人正常乳腺的影响[J].中国医学科学院学报,2004,26(5):566-570.
[40] Shiera WT,Shiera AC,Xieb CJ.Structure activity relationships for human estrogenic activity in zearalenone[J].Mycotoxins Toxicon,2001,(39):1435-1438.
[41] 戴顺龄,段金虹,陆媛,等.新的植物雌激素α-玉米赤霉醇明显抑制实验性动脉粥样硬化发病进程[J].中国动脉硬化杂志,2003,5(11)385-390.
[42] 戴顺龄,段金虹,等.植物雌激素α-玉米赤霉醇与17β-雌二醇、戊酸雌二醇对兔食饵性动脉粥样硬化的作用[J].微循环学杂志,2006,16(4):15-18.
[43] 徐海珊,段金虹,戴顺龄,等.植物雌激素α-ZAL对oxLDL诱导的人内皮细胞分泌NO和ET.1的影响及其机制研究[J].中国药理学通报,2005,21(6):692-769.
[44] Jeffrey AD,Andrew LG,David EM,etal.Short and Henry U.BryantEnvironmental Estrogens:Effects on Cholesterol Lowering and Bone in the Ovariectomized Rat[J].Steroid Biochem Molec BioL,1996,59(2):155-161.
[45] Chanetsa F,Hillman LS,Thomas MG,Keisler DH.Estrogen Agonist(Zeranol)Treatment in a Castrated Male Lamb Model:Effects on Growth and Bone Mineral Accretion[J].Journal of Bone And Mineral Research,2000,15(7):1361-1367.
[46] Wong R,Rabie B.Effect of puerarin on bone formation[J].OsteoArthritis and Cartilage,2007,15:894-899.
[47] Maitrayee Mukherjee,Asankur Sekhar Das,Dolan Das,et al.Role of Oil Extract of Garlic(Allium sativum Linn.)on Intestinal Transference of Calcium and Its Possible Correlation with Preservation of Skeletal Health in an Ovariectomized Rat Model of Osteoporosis[J].Phytotherapy Research,2006,20:408-415.
猜你喜欢
中国骨质疏松杂志(2024年2期)2024-03-19 09:30:14
中国骨质疏松杂志(2021年9期)2021-10-08 10:07:40
天津中医药(2020年5期)2020-06-01 12:16:14
中国临床医学(2019年3期)2019-01-04 09:12:32
天然产物研究与开发(2018年5期)2018-06-13 03:23:42
中成药(2017年6期)2017-06-13 07:30:35
中成药(2017年4期)2017-05-17 06:09:27
中国民族医药杂志(2016年6期)2016-05-09 08:52:57
应用化工(2014年5期)2014-08-08 13:10:58
中医研究(2014年4期)2014-03-11 20:28:42