
单宁酸体外抑制痘苗病毒感染活性的效果分析
2015-11-29 08:31:50刘璇杨益李瑾慧孙走南唐玥苏文莉刘京梅于雅琴常国辉
生物技术通讯 2015年6期
刘璇 ,杨益,李瑾慧,孙走南,唐玥,苏文莉,刘京梅,于雅琴,常国辉
1.吉林大学 公共卫生学院,吉林 长春 130021;2.军事医学科学院 疾病预防控制所,北京 100071
植物多酚是一类种类繁多的高分子多元酚化合物,为植物的次级代谢产物,广泛存在于植物体的叶、果肉及树皮中,包括药材、粮食、水果和蔬菜等,并且有很强的生物学活性,能与蛋白质、核酸和金属离子等相互作用[1-3]。单宁酸(tannic acid)是简单的水解类植物多酚,其抗肿瘤、抗氧化及抗感染等特性已多有报道[4]。近年来,单宁酸作为广谱低毒的病毒抑制剂而备受研究者关注[5]。单宁酸的生物学效应可能与其和细胞膜相互作用的能力密切相关[6]。膜荧光各向异性和跨膜电位是细胞膜的重要生物物理学特性,有研究表明,病毒侵入可改变宿主细胞膜的流动性和膜电位[7-8]。因此,我们以痘苗病毒(vaccinia virus,VACV)WR(Western Reserve)株为研究对象,测定病毒感染对细胞膜流动性和膜电位的影响及单宁酸的干预作用,为揭示单宁酸抑制病毒感染的作用机制提供实验依据。
1 材料与方法
1.1 材料
Vero E6 细胞由本实验室细胞库保存;VACV WR株由英国布里斯托大学Siddell教授惠赠。
胎牛血清(Sigma,美国);DMEM、胰蛋白酶(Gibco,美国);荧光探针DPH(用四氢呋喃配成2.0×10-3mol/L 母液,避光保存于-20℃,临用前用PBS 稀释)(Aldrich,美国);荧光探针DiBAC4(3)(用无水乙醇配成1 mg/mL 母液,避光保存于-20℃,临用前用PBS 稀释)(Molecular Probe,美国);单宁酸(浙江省温州市瓯海精细化工有限公司);中性红(国药集团化学试剂有限公司);SpectraMax M5 多功能酶标仪(美谷分子仪器公司,美国);FACSCalibur 流式细胞仪(BD公司,美国)。
1.2 实验分组
实验分为正常对照组、病毒组、单宁酸组、病毒加单宁酸组(病毒+单宁酸组)。病毒+单宁酸组分为先加入单宁酸后感染病毒(吸附前组)、先感染病毒后加入单宁酸(吸附后组)和感染病毒的同时加入单宁酸(吸附同时组)。
1.3 单宁酸细胞毒性测定
取对数生长期的Vero E6 细胞,按8×104/mL 浓度每孔200 μL接种于96孔板,37℃、5% CO2培养箱中培养,待长成单层细胞后,移去培养液上清,分别加入浓度为1、2、4、8、16、32、64、128 μg/mL 的含单宁酸维持液,每孔100 μL;培养24 h 后,采用中性红比色法,在酶标仪上测定D492nm值,按下式计算细胞存活率,以Probit 回归方法计算单宁酸半数中毒浓度(TC50)。各组均设4个复孔,实验重复3次。
细胞存活率(%)=(试验孔D492nm/对照孔D492nm)×100%
1.4 单宁酸抑制VACV活性测定
制备已长满单层的96孔细胞培养板,使用单宁酸1~16 μg/mL,分别于病毒吸附前、吸附后及吸附同时干扰病毒感染细胞:①吸附前:单宁酸溶液100 μL/孔孵育1 h,弃液,PBS 洗涤,100 TCID50病毒液100 μL/孔吸附1 h;②吸附后:100 TCID50病毒液100 μL/孔吸附1 h,弃液,PBS 洗涤,加单宁酸溶液100 μL/孔孵育1 h;③吸附同时:将等体积的100 TCID50病毒液与单宁酸溶液混合后,于细胞培养板加混合液100 μL/孔,37℃孵育1 h。
将上述处理的细胞培养板内液体吸弃,PBS 洗涤后加入维持培养基。设正常细胞对照孔和病毒对照孔,每天于倒置显微镜下观察细胞变化。置37℃、5% CO2培养箱中培养72 h 后,用中性红比色法检测。各组均设4个复孔,实验重复3次。
以Probit 回归方法计算半数抑制浓度(IC50);按下式计算病毒抑制率:
病毒抑制率(%)=(药物处理组D492nm-病毒对照组D492nm)/(细胞对照组D492nm-病毒对照组D492nm)×100%
按下式计算治疗指数(TI):
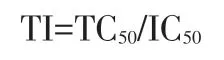
1.5 荧光偏振法测定细胞膜流动性
在Vero E6 细胞(密度为2×106/mL)中加入100 TCID50的病毒液或16 μg/mL 单宁酸溶液进行孵育。正常对照组加入1 mL细胞维持液,病毒组加入1 mL病毒液,单宁酸组加入1 mL单宁酸溶液,吸附同时组加入1 mL病毒和单宁酸的等体积混合液,以上各组均于37℃孵育1 h。吸附前组先加1 mL 单宁酸溶液,37℃孵育1 h,PBS离洗后再加1 mL病毒液,37℃孵育1 h;吸附后组先加1 mL 病毒液,37℃孵育1 h,PBS洗涤后再加1 mL单宁酸溶液,37℃孵育1 h;孵育时须每15 min摇动混匀一次。
上述各组细胞悬液于1000 r/min离心5 min,弃上清,加入2×10-6μmol/mL DPH 荧光探针溶液1 mL,37℃孵育并不断振荡30 min,再用PBS 液洗一次,于激发波长365 nm、发射波长430 nm测定荧光各向异性(r)。r值越大,表示细胞膜流动性越小;反之,流动性越大。
1.6 流式细胞术测定细胞膜电位
细胞处理同1.5,细胞悬液1000 r/min 离心5 min,弃上清,加入0.1 μg/mL DiBAC4(3)荧光探针溶液1 mL,37℃孵育30 min 后上机测定,用未经荧光物质孵育的Vero E6 细胞调零。荧光增强,表示细胞膜电位绝对值减小,出现去极化变化;反之,荧光减弱,细胞膜电位绝对值增大,出现超极化变化。
1.7 统计学分析
应用SPSS17.0 统计软件对结果进行分析,实验数据以x±s表示。组间比较采用方差分析,P<0.05认为差异有统计学意义。
2 结果
2.1 单宁酸对Vero E6细胞的毒性作用
单宁酸对Vero E6细胞的毒性作用表现为细胞增殖缓慢、颗粒较多、折光性强。单宁酸对细胞的毒性作用随着浓度的降低而逐渐减弱。中性红比色法测得单宁酸TC50为81.0 μg/mL,TC0为16.0 μg/mL。
2.2 单宁酸抑制VACV感染效果
实验结果见图1,单宁酸对病毒的抑制作用随着浓度的增加而增强。病毒吸附后加入单宁酸,病毒抑制率为18.7%~69.3%(IC50为8.7 μg/mL,TI 为9.3);病毒吸附同时加入单宁酸,病毒抑制率为11.7%~60.0%(IC50为13.1 μg/mL,TI为6.2)。
2.3 单宁酸对VACV 感染后细胞膜流动性变化的干预作用
VACV感染后宿主细胞膜荧光各向异性显著低于正常细胞对照组(P<0.05),说明病毒感染后宿主细胞膜流动性升高。单宁酸处理Vero E6细胞后荧光各向异性与正常对照组接近,说明单宁酸对正常细胞膜流动性无明显影响。3 种加入方式中,单宁酸在病毒吸附前和病毒吸附后作用,均可使荧光各向异性升高,其中吸附后组与正常对照组比较有显著性差异(P<0.01),说明单宁酸在VACV 吸附后作用可降低宿主细胞膜流动性(表1)。
2.4 单宁酸对VACV 感染后细胞膜电位变化的干预作用
VACV感染Vero E6细胞后,荧光强度显著低于正常细胞对照组(P<0.01),说明病毒感染后宿主细胞膜处于超极化状态。单宁酸作用于Vero E6细胞后,荧光强度显著低于正常细胞对照组(P<0.01),说明单宁酸作用同样使细胞膜处于超极化状态。单宁酸3种处理方式下,VACV吸附各组的荧光强度值与病毒对照组比较无显著性差异,说明单宁酸不同处理方式对VACV感染各组中膜电位的变化无明显干预作用(表2)。
3 讨论
单宁酸是相对分子质量为500~3000 的天然酚类化合物,在自然界分布广泛,因其具有多酚羟基结构而表现出独特的化学特性和生理活性。VACV是只在靶细胞的胞浆中复制的双链DNA(dsDNA)病毒。成熟的病毒粒子呈椭圆形或砖形,外层有较厚的囊膜包裹,其囊膜含有多种病毒蛋白。病毒感染和复制的过程包括3个步骤:①病毒的吸附、侵入和脱壳;②基因组的转录和复制;③病毒的组装、成熟和释放。本实验中,单宁酸以不同加入方式干扰病毒感染细胞,结果显示,单宁酸以病毒吸附后及病毒吸附同时2种方式加入时,可有效抑制VACV 感染,于吸附前孵育细胞对VACV 侵入细胞无阻断作用,提示单宁酸抑制VACV感染的机制可能存在于病毒吸附同时和吸附后。

图1 单宁酸不同加入方式抑制VACV感染效果(,n=3)
病毒的感染首先依赖于病毒吸附蛋白对细胞表面受体的吸附,随后发生的侵入也同病毒感染成功与否密切相关,这些环节均涉及病毒与细胞膜的相互作用。细胞膜脂质和脂蛋白质都有一定的流动性,细胞膜流动性的改变可影响病毒吸附和侵入细胞[9-10]。如吩噻嗪通过增强细胞膜流动性从而达到抑制丙型肝炎病毒(HCV)与细胞膜融合的效果[11];而Harada 等发现,在病毒吸附同时和吸附后2 个阶段,细胞膜流动性的升高都可以增加人免疫缺陷病毒(HIV)感染率[12]。有研究表明,植物多酚可嵌入细胞脂质双层膜的表层,影响细胞膜生物学特性[13]。本研究结果显示,单宁酸对正常细胞膜流动性无明显影响,而VACV 感染后宿主细胞膜流动性显著升高。3 种不同加入方式中,只有在感染后加入单宁酸,可以干预病毒感染对细胞膜的影响,细胞膜流动性显著低于正常对照组。提示单宁酸抑制病毒感染可能存在以下机制:VACV 感染后细胞膜流动性的增加易于单宁酸嵌入脂质双层膜,致使细胞膜流动性显著降低,膜流动性的降低干扰了VACV 入胞过程,从而抑制病毒感染。
细胞膜是病毒侵入细胞的第一道重要防线,膜电位是细胞膜重要的生物物理特性。从病毒吸附蛋白与细胞受体特异性结合到内吞入胞,病毒侵入细胞是一个能量依赖的过程。研究表明,脊髓灰质炎病毒和人鼻病毒侵入宿主细胞过程涉及细胞膜电位变化,细胞膜跨膜电位差是促使病毒内化的能态之一[14-15]。本实验结果表明,VACV 感染Vero E6 细胞后,可使细胞膜发生超极化,形成局部电流,跨膜电位差利于病毒穿入靶细胞内。单宁酸亦使细胞膜处于超极化状态,并不能有效降低病毒感染引起的细胞超极化现象,说明对病毒借助跨膜电位差进入宿主细胞这一过程无显著干预作用,由此我们推测单宁酸抑制病毒感染的作用机制可能与此环节无关。
表1 单宁酸对VACV感染后细胞膜流动性变化的干预作用(,n=3)

表1 单宁酸对VACV感染后细胞膜流动性变化的干预作用(,n=3)
与正常对照组比较,aP<0.05,bP<0.01
表2 单宁酸对VACV感染后细胞膜电位变化的干预作用(,n=3)
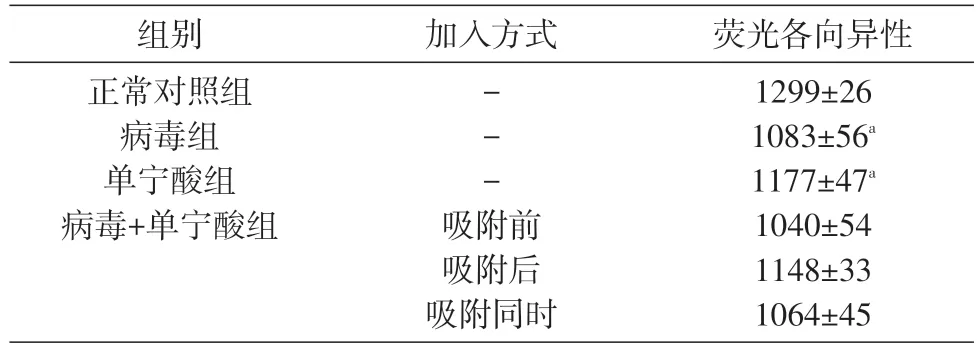
表2 单宁酸对VACV感染后细胞膜电位变化的干预作用(,n=3)
与正常对照组比较,aP<0.01
综上所述,单宁酸在体外有较强的抑制痘苗病毒感染的作用,其作用机制可能与干预细胞膜流动性有关,具体作用靶点还有待进一步研究。
[1]Haslam E.Vegetable tannins — lessons of a phytochemical lifetime[J].Phytochemistry,2007,68(22-24):2713-2721.
[2]Frazier R A,Deaville E R,Green R J,et al.Interactions of tea tannins and condensed tannins with proteins[J].J Pharm Biomed Anal,2010,51(2):490-495.
[3]Li M,Hagerman A E.Interactions between plasma proteins and naturally occurring polyphenols[J].Curr Drug Metab,2013,14(4):432-445.
[4]Cho H S,Lee J H,Cho M H,et al.Red wines and flavonoids diminish Staphylococcus aureus virulence with anti-biofilm and anti-hemolytic activities[J].Biofouling,2015,31(1):1-11.
[5]Kanwar J,Taskeen M,Mohammad I,et al.Recent advances on tea polyphenols[J].Front Biosci,2012,4(4):111-131.
[6]Abram V,Berlec B,Ota A,et al.Effect of flavonoid structure on the fluidity of model lipid membranes[J].Food Chem,2013,139(1-4):804-813.
[7]Matsuda K,Hattori S,Kariya R,et al.Inhibition of HIV-1 entry by the tricyclic coumarin GUT-70 through the modification of membrane fluidity[J].Biochem Biophys Res Commun,2015,457(3):288-294.
[8]Kuo Y C,Lin Y L,Liu C P,et al.Herpes simplex virus type 1 propagation in HeLa cells interrupted by Nelumbo nucifera[J].J Biomed Sci,2005,12(6):1021-1034.
[9]Franquelim H G,Gaspar D,Veiga A S,et al.Decoding distinct membrane interactions of HIV-1 fusion inhibitors using a combined atomic force and fluorescence microscopy approach[J].Biochim Biophys Acta,2013,1828(8):1777-1785.
[10]Amaral L,Viveiros M,Molnar J.Antimicrobial activity of phenothiazines[J].In Vivo,2004,18(6):725-731.
[11]Chamoun-Emanuelli A M,Pecheur E I,Simeon R L,et al.Phenothiazines inhibit hepatitis C virus entry,likely by increasing the fluidity of cholesterol-rich membranes[J].Antimicrob Agents Chemother,2013,57(6):2571-2581.
[12]Harada S,Yusa K,Monde K,et al.Influence of membrane fluidity on human immunodeficiency virus type 1 entry[J].Biochem Biophys Res Commun,2005,329(2):480-486.
[13]Verstraeten S V,Jaggers G K,Fraga C G,et al.Procyanidins can interact with Caco-2 cell membrane lipid rafts:involvement of cholesterol[J].Biochim Biophys Acta,2013,1828(11):2646-2653.
[14]Irurzun A,Carrasco L.Entry of poliovirus into cells is blocked by valinomycin and concanamycin A[J].Biochemistry,2001,40(12):3589-3600.
[15]Berka U,Khan A,Blaas D,et al.Human rhinovirus type 2 uncoating at the plasma membrane is not affected by a PH gradient but is affected by the membrane potential[J].J Virol,2009,83(8):3778-3787.
猜你喜欢
食品工业(2024年3期)2024-04-13 11:03:58
湖南饲料(2023年2期)2023-06-20 02:39:28
中老年保健(2022年1期)2022-08-17 06:14:30
中国生物医学工程学报(2019年6期)2019-07-16 07:52:58
兽医导刊(2019年1期)2019-02-21 01:14:20
猪业科学(2018年8期)2018-09-28 01:27:52
天然产物研究与开发(2018年4期)2018-05-07 06:48:00
中国医疗美容(2015年4期)2015-04-27 02:24:11
科学中国人(2015年13期)2015-02-28 09:13:26
食品工业科技(2014年23期)2014-03-11 18:18:53
