
甘肃河西地区西门塔尔杂交类群NGB基因第3外显子的遗传变异研究
2015-10-26 08:51梁春花刘霞孙雪婧罗玉柱高雪晋杜晓华
生物技术通报 2015年2期
梁春花刘霞孙雪婧罗玉柱高雪晋杜晓华
(1. 甘肃农业大学动物医学院,兰州 730070;2. 甘肃农业大学生命科学技术学院,兰州 730070;3. 甘肃省草食动物生物技术重点实验室,兰州 730070)
甘肃河西地区西门塔尔杂交类群NGB基因第3外显子的遗传变异研究
梁春花1,3刘霞2,3孙雪婧1,3罗玉柱3高雪晋2杜晓华1,3
(1. 甘肃农业大学动物医学院,兰州 730070;2. 甘肃农业大学生命科学技术学院,兰州 730070;3. 甘肃省草食动物生物技术重点实验室,兰州 730070)
旨在对甘肃河西的临泽、甘州、武威、金昌、高台5个地区283头西门塔尔杂交类群NGB基因第3外显子的遗传多态性及变异特征进行系统分析,采用PCR-SSCP方法检测了283头西门塔尔杂交类群NGB基因第3外显子和部分内含子的多态性,且对群体内各等位基因进行了测序。结果显示,5个地区西门塔尔杂交类群共检测出5个等位基因(A、B、C、D、E),表现为5种基因型(AA、AB、AC、AD、AE)。其中甘州、武威、金昌西门塔尔杂交类群NGB基因均只检测到AA、AB 2种基因型,高台西门塔尔杂交类群检测到AA、AE 2种基因型,临泽西门塔尔杂交类群检测到AA、AB、AC、AD 4种基因型。A等位基因和AA基因型的频率在5个群体中最高,为优势基因和优势基因型。对不同SSCP带型的对应片段进行测序分析,共发现6个核苷酸突变位点(75 bp C→T,78 bp C→G,128 bp G→A,214 bp G→A,232 bp C→T,233 bp G→A),其中第75 bp和第78 bp处的突变位点位于内含子区域,其余4处突变位点均位于外显子区域。第214 bp处的核苷酸突变导致甘氨酸(Gly)突变为丝氨酸(Ser),第232 bp处核苷酸突变导致精氨酸(Arg)突变为色氨酸(Trp),第233 bp处核苷酸突变导致精氨酸(Arg)突变为谷氨酰胺(Gln),经χ2检验结果显示,5个地区的西门塔尔杂交类群在此3个突变位点上都处于Hardy-Weinberg平衡状态(P>0.05)。群体遗传学分析结果表明,临泽、甘州、武威、金昌、高台西门塔尔杂交类群的多态信息含量(PIC)分别为0.0582、0.0196、0.0196 、0.0161、0.0159,均属于低度多态(PIC<0.25)。
西门塔尔杂交类群;NGB基因;外显子;遗传变异
2000年德国科学家Burmester等[1]首次在Nature上报道人和小鼠脑内存在第3种携氧球蛋白——脑红蛋白(neuroglobin,NGB)。此后,科学家们逐渐发现NGB存在于各种哺乳动物、鸟类、爬虫类、两栖类、鱼类等。NGB主要在脑组织中表达,能够可逆地结合氧,从而特异性地向脑组织供氧。缺氧能够诱导NGB的表达,而高表达的NGB则能够保护神经元免受缺氧损伤,从而在神经系统缺氧/缺血损伤中具有重要的神经保护功能,已成为国内外学者研究的热点[2-5],但有关NGB作用的生理机制仍不明确。
西门塔尔牛原产于瑞士西门山谷地带(Simon Valley),其属大型乳、肉、役兼用品种,是世界第二大牛种。甘肃河西地区自1980年开始引用西门塔尔牛冻精进行杂交改良本地黄牛的产肉产奶性能[6],通过30多年持续不断地级进改良和导入选育,基本形成了适应甘肃河西地区的西门塔尔杂交类群。目前,国内外对NGB基因的研究多见于人和大鼠,对牦牛、藏羚羊、高原兔、藏鸡等也都有相关的研究[7-13],但有关西门塔尔牛NGB基因的研究尚未见报道。本试验采用PCR-SSCP方法,对甘肃河西地区西门塔尔杂交类群NGB基因第3外显子的遗传变异规律进行了研究,其结果能够为进一步了解西门塔尔杂交类群NGB基因的遗传特性提供有价值的理论依据,对于探讨NGB基因在缺氧/缺血状态下的保护性生理机制也具有一定的理论与实践意义。
1 材料与方法
1.1 材料
1.1.1 试验动物血样 随机选取甘肃河西5个地区共283头西门塔尔杂交类群。其中,临泽地区50头(临泽县位于甘肃省河西走廊中部,海拔1380-2 278 m)、甘州地区50头(甘州区位于甘肃省河西走廊中部,平均海拔1 474 m)、武威地区60头(武威市位于河西走廊东端,海拔1 440-3 263 m)、金昌地区61头(金昌市位于甘肃省河西走廊东端,海拔1 500-4 442 m)、高台地区62头(高台县位于甘肃省河西走廊中部,黑河终段下游,海拔1 260-3 140 m),颈静脉采血10 mL,ACD(酸性柠檬酸葡萄糖)法抗凝,-70℃冻存。
1.1.2 试剂与仪器 蛋白酶K、Tris平衡酚购自大连宝生物工程有限公司;硼酸、乙二胺四乙酸(EDTA)购自Amersco公司;过硫酸铵购自烟台市双双化工有限公司;去离子甲酰胺、TEMED购自Sigma公司;丙烯酰胺、甲叉双丙稀酰胺购自上海生工生物工程股份有限公司;2×Power Taq PCR Master Mix购自北京百泰克生物技术公司;Accurate Run 100 bp-I DNA ladder购自上海捷瑞生物工程有限公司;其他常规试剂均为进口或国产分析纯级产品。
移液枪购自德国Eppendorf公司;梯度PCR仪购自美国ABI公司;DYY-11电泳仪、DYCP-31DN水平电泳槽购自北京六一仪器厂;A-6030凝胶成像分析系统购自韩国BIONEER公司;PROTEAN Ⅱ xi Cell双垂直电泳槽、042BR10095 PowerPac Universal电泳仪购自美国BIO-RAD公司;F12加热/制冷循环仪购自德国JULABO公司。
1.2 方法
1.2.1 基因组DNA提取 用常规酚-氯仿抽提法从冻存血样中提取基因组DNA,-20℃保存备用。
1.2.2 引物设计与PCR扩增 参考GenBank中普通牛NGB基因序列(登录号:AC_000167.1),用Primer 3.0引物设计软件设计西门塔尔杂交类群NGB基因第3外显子的引物,上游引物:5'-ACACAGGCTGCCTTTTGTCT-3'、下游引物:5'-GGGAGAGATGAACAGGTGGA-3',预扩增片段大小约为356 bp,包括两端的部分内含子区域,引物由北京六合华大基因科技股份有限公司合成。
PCR反应体系为20 μL:ddH2O 7.4 μL,上游和下游引物各0.4 μL,2×Power Taq PCR Master Mix 11 μL,模板DNA 0.8 μL。PCR反应程序为:94℃预变性3 min;94℃变性30 s,60℃退火30 s,72℃延伸30 s,共35个循环;72℃终延伸7 min,-4℃保存。PCR产物用10 g/L的琼脂糖凝胶电泳检测。
1.2.3 PCR-SSCP分析 取2 μL的PCR产物加入8 μL的上样缓冲液(980 mL/L去离子甲酰胺,0.25 g/L溴 酚 蓝,0.25 g/L二 甲 苯 青,pH8.0、0.01 mol/L EDTA),105℃变性10 min,迅速冰浴5 min后上样于浓度为14%的100 g/L(Acr∶Bis的质量比为39∶1)的非变性聚丙烯酰胺凝胶,18℃、260 V电泳21 h,银染法显色后拍照。
观察电泳结果,选取条带不同的PCR产物送往北京六合华大基因科技股份有限公司进行测序。
1.2.4 统计分析 运用Popgene 1.32软件计算基因型频率(Genotype frequency)、基因频率(Allele frequency)、基因杂合度(Heterozygosity,He)、有效等位基因数(Effective number of alleles,Ne)以及进行Hardy-Weinberg平衡的χ2检验等;用PIC_Calc 0.6软件计算多态信息含量(Polymorphism information content,PIC);使用Lasergene7.1软件包中的Meg-Align软件进行同源性分析及构建系统进化树[14]。
2 结果
2.1 NGB基因第3外显子的PCR扩增
甘肃河西5个地区西门塔尔杂交类群NGB基因第3外显子的扩增产物用10 g/L琼脂糖凝胶电泳检测,结果(图1)与预扩增片段大小一致且条带清晰明亮,可直接用于SSCP分析。
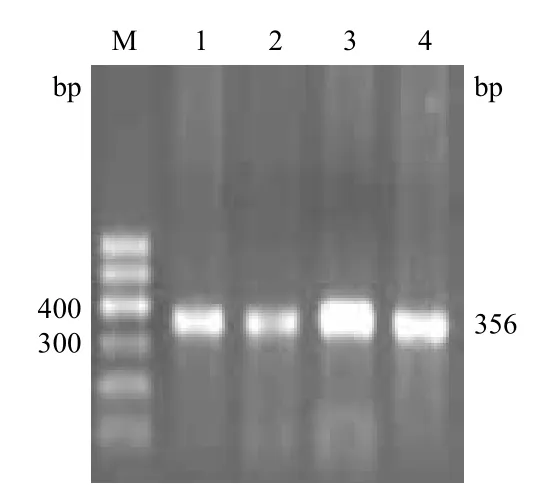
图1 西门塔尔杂交类群NGB基因第3外显子的PCR扩增
2.2 SSCP检测
SSCP检测结果(图2)显示,甘肃河西5个地区的283头西门塔尔杂交类群中共发现AA、AB、AC、AD、AE 5种基因型,受A、B、C、D、E 5个等位基因控制。

图2 西门塔尔杂交类群NGB基因第3外显子PCR-SSCP检测
对具有多态性的PCR产物进行测序,得到西门塔尔杂交类群NGB基因第3外显子356 bp的核苷酸序列,其中A等位基因序列与普通牛(登录号:AC_000167.1)的相应序列一致。将不同等位基因的测序结果进行比对(图3),发现与A等位基因相比,B等位基因在第128 bp处发生了G→A突变,但未引起氨基酸序列的变化,为同义突变;C等位基因在第214 bp处发生了G→A突变,导致甘氨酸(Gly)突变为丝氨酸(Ser);D等位基因在第232 bp处发生了C→T突变,导致精氨酸(Arg)突变为色氨酸(Trp);E等位基因在第233 bp处发生了G→A突变,导致精氨酸(Arg)突变为谷氨酰胺(Gln),在第75 bp和第78 bp处(即内含子区域)分别发生了C→T和C→G突变。
2.3 遗传多样性分析
5个地区西门塔尔杂交类群NGB基因第3外显子等位基因的基因型频率和基因频率分布,见表1。群体中发现AA、AB、AC、AD、AE 5种基因型,受A、B、C、D、E 5个等位基因控制。甘州、武威、金昌西门塔尔杂交类群中均只检测到AA、AB两种基因型,高台西门塔尔杂交类群检测到AA、AE两种基因型,临泽西门塔尔杂交类群检测到AA、AB、AC、AD 4种基因型,5个地区均只发现AA一种纯合基因型,其余均为杂合型。A等位基因和AA基因型的频率在5个群体中最高,为优势基因和优势基因型。5个地区西门塔尔杂交类群NGB基因第3外显子多态性群体遗传结构分布,见表2。遗传杂合度(He)、有效等位基因数(Ne)和多态信息含量(PIC)是评价群体遗传变异的重要指标,不同的遗传参数体现各群体的遗传差异性。由表2可知,武威地区西门塔尔杂交类群的群体杂合度和有效等位基因数最大,表示其遗传多样性最丰富,遗传变异程度最高,具有较大的遗传选择潜力;甘州西门塔尔杂交类群的群体杂合度和有效等位基因数最低,其遗传变异程度最小。多态信息含量(PIC)是衡量标记多态性较好的指标,不同地区西门塔尔杂交类群的PIC值在0.015 9-0.058 2之间,甘肃河西5个地区西门塔尔杂交类群均表现为低度多态(PIC<0.25)。Hardy-Weinberg平衡检验结果显示,5个地区的西门塔尔杂交类群均处于平衡状态(P>0.05)。

图3 西门塔尔杂交类群NGB基因第3外显子等位基因的核苷酸序列比对

表1 5个地区西门塔尔杂交类群NGB基因第3外显子的基因频率和基因型频率

表2 NGB基因第3外显子多态性群体遗传结构
2.4 西门塔尔牛杂交类群NGB基因编码区核苷酸序列同源性及系统发育分析
西门塔尔牛杂交类群与8个物种NGB基因编码区核苷酸序列同源性,见表3。西门塔尔杂交类群与牦牛和绵羊的同源性最高,分别为98.3%、99.2%,与野猪、猴、人、黑猩猩、兔、鸡同源性依次降低,分别为92.5%、93.3%、92.5%、91.7%、91.7%、72.5%。9个物种NGB基因编码区核苷酸序列的系统进化树,见图4。西门塔尔杂交类群与牦牛和绵羊亲缘关系最近,而与野猪、猴、人、黑猩猩、兔、鸡亲缘关系依次渐远,其NGB基因编码区核苷酸序列的系统发育树与生物进化的物种树以及动物分类学观点也基本一致。
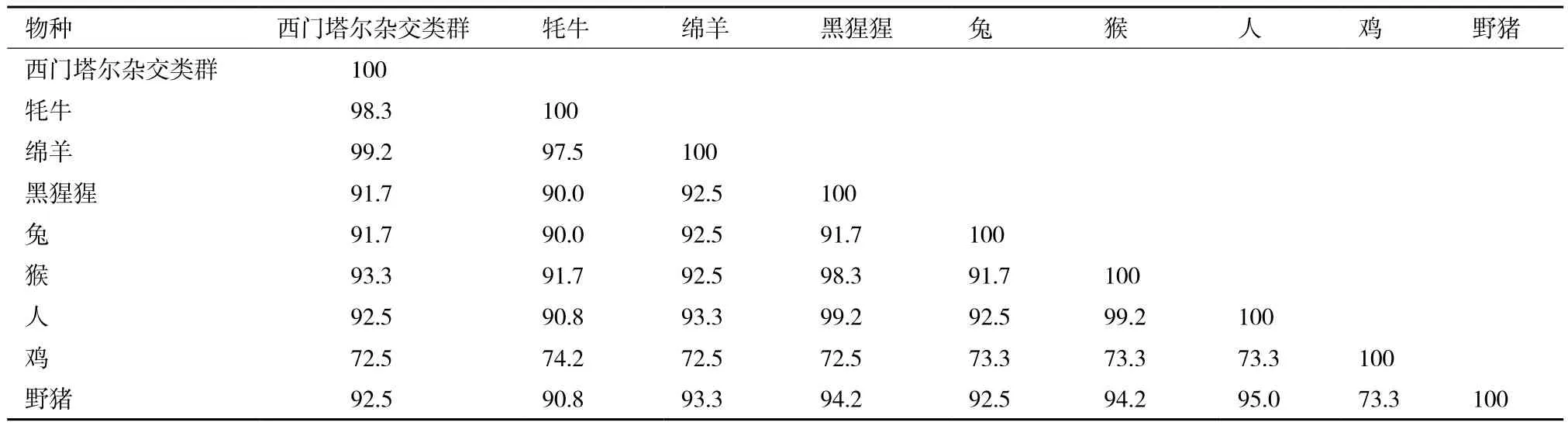
表3 西门塔尔杂交类群与8个物种NGB基因编码区核苷酸序列同源性(%)比较
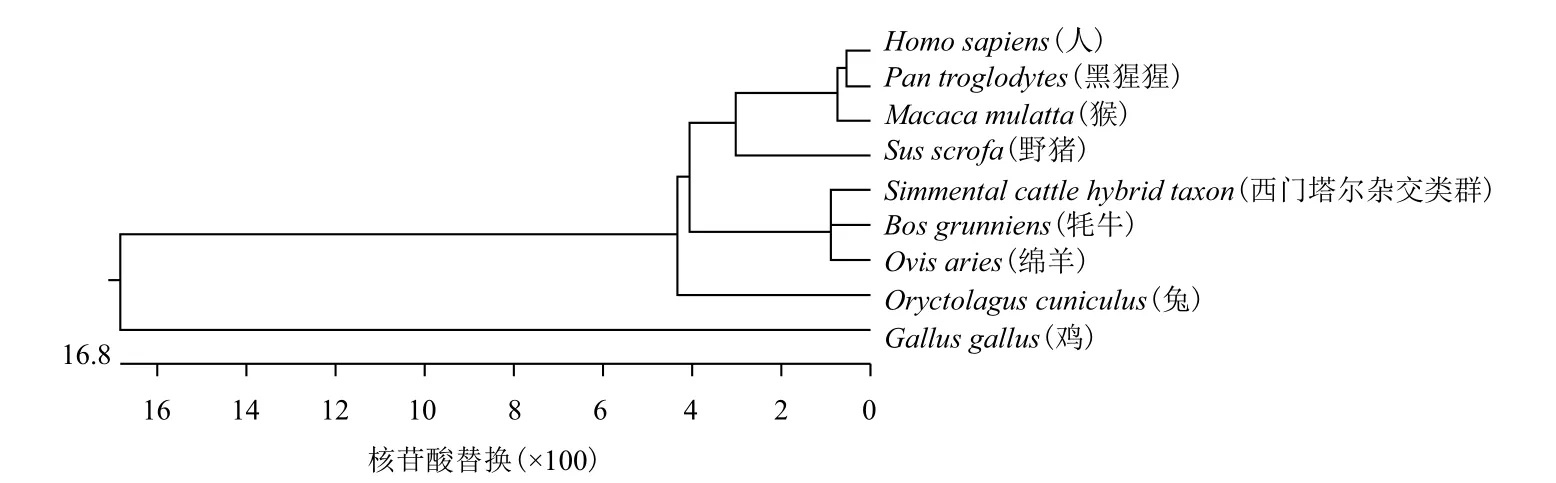
图4 九个物种NGB基因编码区核苷酸序列的系统进化树
3 讨论
本研究采用PCR-SSCP方法分析了甘肃河西5个地区283头西门塔尔杂交类群NGB基因第3外显子的遗传多态性,研究结果显示,河西不同地区西门塔尔杂交类群NGB基因第3外显子具有一定的遗传多样性。共发现6个核苷酸突变位点(75 bp C→ T,78 bp C→ G,128 bp G→ A,214 bp G→A,232 bp C→T,233 bp G→A),其中第75 bp和第78 bp处突变位点落在内含子区域,可能会对NGB基因的表达调控产生一定的影响。其余4处突变位点均落在外显子区域,第128 bp处的核苷酸突变未导致氨基酸的改变,为同义突变。第214 bp处的核苷酸突变导致甘氨酸(Gly)突变为丝氨酸(Ser),第232 bp处的核苷酸突变导致精氨酸(Arg)突变为色氨酸(Trp),第233 bp处的核苷酸突变导致精氨酸(Arg)突变为谷氨酰胺(Gln),这些氨基酸突变形成的原因可能与河西各地区的生态环境、地理条件和选育方法等不同有关。其中第232 bp处的核苷酸突变导致精氨酸(Arg)突变为色氨酸(Trp),其侧链上增加了苯环结构,可能引起蛋白质的空间位阻效应,导致其结构发生改变。这些氨基酸的突变是否会对整个NGB的功能及其在群体中的遗传效应产生影响,还有待更进一步的研究证实。
根据核苷酸突变位点的不同,形成了AA、AB、AC、AD、AE 5种基因型,受A、B、C、D、E 5个等位基因控制。甘州、武威、金昌西门塔尔杂交类群发现了AA、AB 2种基因型,高台西门塔尔杂交类群发现了AA、AE 2种基因型,临泽西门塔尔杂交类群发现了AA、AB、AC、AD 4种基因型。在第233 bp位点只有高台西门塔尔杂交类群发现了E等位基因,第214 bp和第232 bp位点仅临泽西门塔尔杂交类群发现了C、D等位基因,可能与这两个地区西门塔尔杂交类群的育成历史有关,高台西门塔尔杂交类群中的E等位基因和临泽西门塔尔杂交类群中的C、D等位基因是杂交培育过程中固定下来的,而其它地区的西门塔尔杂交类群中并未检测到C、D、E等位基因,说明高台和临泽西门塔尔杂交类群与其它地区西门塔尔杂交类群之间的基因交流很少,属于封闭群体。甘肃河西5个地区西门塔尔杂交类群中仅发现一种AA型纯合体,其基因型频率远高于AB、AC、AD和AE基因型,可能是由于等位基因B、C、D和E在进化及群体遗传过程中受到一定的选择压力,其纯合个体因某种原因被自然选择或人工选择所淘汰,导致其纯合型在群体遗传过程中被稀释或消失,闫永红等[15]推测该过程可能是一种完全淘汰隐性基因的选择效应,而A等位基因在长期的人工选育过程中则受到正向选择,处于优势地位。A等位基因的基因频率范围为0.966 7-0.991 9,属于优势等位基因。其余4个等位基因的基因频率范围为0.008 1-0.033 3,其等位基因频率极低,原因可能是甘肃河西地区西门塔尔杂交类群不断地进行杂交改良,随着代数的增加而导致等位基因B、C、D、E逐渐被稀释。在甘肃河西地区西门塔尔杂交类群NGB基因第3外显子区域发现了第128 bp处G/A的转换、第214 bp 处G/A的转换、第232 bp处 C/T的转换、第233 bp处G/A的转换,这4处突变类型均为转换,说明其突变类型比较保守,有利于保持其群体的遗传稳定性。
多态信息含量(PIC)、杂合度(He)和有效等位基因数(Ne)都可用来度量群体内遗传变异,其值的高低反映了群体内个体的均质度,其数值越高,说明遗传变异性就越大,对环境的适应能力越强,具有较大的选择潜力,应用于遗传育种效果越好。根据PIC>0.5为高度多态,PIC<0.25为低度多态,0.25<PIC<0.5则为中度多态[16]。本研究中,甘肃河西5个地区西门塔尔杂交类群NGB基因第3外显子基因座位均呈现低度多态(PIC<0.25),说明5个地区西门塔尔杂交类群在此基因座位的遗传多样性偏低,这可能由于甘肃河西地区引进西门塔尔牛时间较短,繁育次数和代数不多,导致基因多样性较低。武威地区西门塔尔杂交类群的群体杂合度和有效等位基因数最大,遗传多样性最丰富,遗传变异程度最高,遗传潜力大;甘州地区西门塔尔杂交类群的群体杂合度和有效等位基因数最低,其遗传变异程度最小,说明该地区的西门塔尔杂交类群遗传一致性较好。χ2检验结果显示,甘肃河西5个地区西门塔尔杂交类群均处于Hardy-Weinberg平衡状态(P>0.05),可能是由于这5个地区的西门塔尔杂交类群经过长期的杂交改良,选择压力相对趋于稳定,因此在人工选择、迁徙和遗传漂变等各种因素的作用下该基因座位处于动态平衡中。
本研究显示,西门塔尔杂交类群与牦牛和绵羊NGB基因编码区核苷酸序列的同源性最大,亲缘关系最近,而与野猪、猴、人、黑猩猩、兔、鸡的同源性依次降低,亲缘关系也依次渐远。说明NGB基因编码区在长期的生物进化过程中存在一定的差异性,可能会导致NGB基因发生遗传变异。为进一步探讨西门塔尔杂交类群NGB基因的遗传特性提供一定的理论依据。
4 结论
本试验运用PCR-SSCP方法对甘肃河西5个地区西门塔尔杂交类群NGB基因第3外显子进行了多态性分析,结果表明,该基因座位共发现5个核苷酸突变位点,受A、B、C、D、E 5个等位基因控制,存在AA、AB、AC、AD、AE 5种基因型。A等位基因和AA基因型为优势基因和优势基因型。该基因座位在5个地区的西门塔尔杂交类群群体中PIC值均呈低度多态(PIC<0.25)。NGB基因编码区在长期的生物进化过程中存在一定的差异性,可能会导致NGB基因发生遗传变异。
[1]Burmester T, Weich B, Reinhardt S, et al. A vertebrate globin expressed in the brain [J]. Nature, 2000, 407(6803):520-523.
[2]Sun Y, Jin K, Mao XO, et al. Neuroglobin is up-regulated by and protects neurons from hypoxic-ischemic injury [J]. Proc Natl Acad Sci USA, 2001, 98(26):15306-15311.
[3]Li RCP, pouranfar F, Lee SK, et al. Neuroglobin protects PC12 cells against beta-amyloid-induced cell injury [J]. Neurobiol Aging,2008, 29(12):1815-1822.
[4]Greenberg DA, Jin K, Khan AA, et al. Neuroglobin:An endogenous neuroprotectant [J]. Current Opinion in Pharmacology,2008, 8(1):20-24.
[5]赵守财, 储照虎, 马领松, 等 . 脑红蛋白在大鼠局灶性脑组织中的保护作用[J].医学论坛杂志, 2009, 30(22):7-9.
[6]张忠, 张生魁, 李学标, 等 . 应用西门塔尔牛冻精改良河西黄牛的效果[J]. 中国畜牧杂志, 1990, 26(4):51-52.
[7]牛建章, 田侠, 赵慧, 等 . 中国人神经珠蛋白(NGB)基因克隆与序列分析[J]. 河北大学学报:自然科学版, 2005, 25(4)412-417.
[8]张成岗, 李林, 邓美玉, 等 . 大鼠脑红蛋白基因编码区的克隆、多态性分析及该基因组织表达谱研究 [J]. 遗传学报, 2001, 28(11):997-1001.
[9]李盛杰, 杜晓华, 罗玉柱, 等 . 天祝白牦牛NGB基因的克隆及生物信息学分析[J]. 畜牧兽医学报, 2013, 44(3):395-398.
[10]石宁宁, 杜晓华, 罗玉柱, 等 . 甘南牦牛NGB 基因克隆及序列分析[J]. 西北农林科技大学学报:自然科学版, 2013, 41(4):1-7.
[11]张立凡, 刘翀, 连林生, 等 . 低氧适应藏鸡神经珠蛋白的突变研究[J]. 自然科学进展, 2008, 18(1):39-44.
[12]马兰, 白振忠, 靳国恩, 等 . 藏羚羊脑红蛋白基因的组织表达谱分析[J]. 青海医学院学报, 2012, 33(1):12-15.
[13]段文娟, 燕振国, 林丽霞, 等. 高原环境下兔眼球挫伤后视网膜脑红蛋白的表达调控研究[J]. 眼外伤职业眼病杂志,2010, 32(2):87-91.
[14]孙雪婧, 杜晓华, 杨孝朴, 等 . 牦牛CYGB 基因CDS 区克隆与生物信息学分析[J]. 中国农业科学, 2014, 47(13):2690-2698.
[15]闫永红, 刘霞, 刘英, 等 . 天祝白牦牛MSTN基因第3外显子的多态性分析[J]. 中国兽医科学, 2011, 41(9):945-949.
[16] Vaiman D, Mercier D, Goudarzi KM, et al. A set of 99 cattle microsatellites:characterization, synteny mapping, and polymorphism [J]. Mammalian Genome, 1994, 5(5):288-297.
(责任编辑 李楠)
Genetic Variation Study of NGB Gene Exon 3 of Simmental Cattle Hybrid Taxon in Gansu Hexi Regions
Liang Chunhua1,3Liu Xia2,3Sun Xuejing1,3Luo Yuzhu3Gao Xuejin2Du Xiaohua1,3
(1. Gansu Agricultural University College of Veterinary Medicine,Lanzhou 730070;2. Gansu Agricultural University College of Life Science and Technology,Gansu Agricultural University,Lanzhou 730070;3. Gansu Key Laboratory of Herbivorous Animal Biotechnology,Lanzhou 730070)
This study was designed to systematically analyze the genetic polymorphism and variability of NGB gene exon 3 of 283 Simmental cattle hybrid taxon in Gansu Linze, Ganzhou, Wuwei, Jinchang, and Gaotai. DNA fragment containing the exon 3 and part of intron of NGB gene was amplified and genotyped using PCR single-strand conformational polymorphism(SSCP) method. Representative alleles were sequenced for verification of their variations in DNA sequences. The results displayed that there were 5 alleles(A, B, C, D and E)detected, combined five genotypes as AA, AB, AC, AD and AE. Among them, samples from Ganzhou, Wuwei and Jinchang were detected AA and AB genotypes, AA and AE genotypes in Gaotai and AA, AB, AC and AD genotypes in Linze. Allele A was the predominant allele and AA was the predominant genotype in five breeds. Sequence of the SSCP showed 6 mutations in the DNA fragment(75 bp C→T, 78 bp C→G,128 bp G→A, 214 bp G→A, 232 bp C→T, and 233 bp G→A). In which, C→T at 75 bp and C→G at 78 bp were located in intron,while the other mutations were located in exon. G→A mutation at 214 bp, C→T mutation at 232 bp and G→A mutation at 233 bp were lead to Gly→Ser, Arg→Trp and Arg→Gln, respectively. Chi-square testing indicated the three gene mutation were all in Hardy-Weinberg equilibrium(P> 0.05). Population genetics analysis showed the polymorphism information content were 0.0582, 0.0196, 0.0196, 0.0161 and 0.0159 in Linze, Ganzhou, Wuwei, Jinchang, Gaotai, respectively, which was at low polymorphism(PIC <0.25).
Simmental cattle hybrid taxon;NGB gene;exon;genetic variation
10.13560/j.cnki.biotech.bull.1985.2015.02.018
2014-10-29
甘肃省高等学校基本科研业务费资助项目(2013),甘肃省教育厅研究生导师项目(1102-04)
梁春花,女,硕士研究生,研究方向:基础兽医学;E-mail:liang835216@163.com
杜晓华,男,副教授,硕士生导师,研究方向:基础兽医学与动物发育生物学;E-mail:duxh@gsau.edu.cn
猜你喜欢
地理教育(2023年1期)2023-02-09
矿产勘查(2020年8期)2020-12-25
江苏农业科学(2019年5期)2019-09-02
长江文艺(2019年5期)2019-08-07
润·文摘(2018年4期)2018-05-14
广东农业科学(2017年5期)2017-08-29
西藏研究(2016年5期)2016-06-15
少儿科学周刊·儿童版(2015年12期)2016-05-16
应用海洋学学报(2014年1期)2014-11-22
东北师大学报(自然科学版)(2014年1期)2014-02-27
