
1—甲基环丙烯控制采后‘油木奈’果实腐烂与抗病相关酶诱导的关系
2015-10-21 19:04李辉林毅雄林河通袁芳林艺芬陈艺晖
热带作物学报 2015年4期
李辉 林毅雄 林河通 袁芳 林艺芬 陈艺晖


摘 要 探讨1-甲基环丙烯(1-MCP)对采后‘油木奈果实腐烂的控制与抗病相关酶诱导的关系。采后‘油木奈果实用0(对照)和1.2 μL/L的1-MCP处理12 h后,在(25±1)℃下贮藏。贮藏期间定期测定腐烂率、抗病相关酶如几丁质酶、β-1,3-葡聚糖酶、苯丙氨酸解氨酶(PAL)、多酚氧化酶(PPO)和过氧化物酶(POD)等活性的变化。研究结果表明:与对照果实相比,1-MCP处理能有效降低‘油木奈果实腐烂率,提高几丁质酶、β-1,3-葡聚糖酶、PPO和POD的活性。因此认为,1-MCP控制采后‘油木奈果实的腐烂与抗病相关酶活性的升高有关,抗病性诱导是1-MCP控制采后‘油木奈果实腐烂的重要原因之一。
关键词 油木奈;果实;1-甲基环丙烯;采后腐烂;控制;抗病相关酶;诱导
中图分类号 TS255.4;S662.3 文献标识码 A
木奈(Prunus salicina Lindl.)又称桃形李、歪嘴李和柰李,属蔷薇科(Rosaceae)李属(Pruns),是中国南方的一种特色水果。木奈果桃形李实,外形似桃,又具李之内涵,兼有桃、李之风味。因此,木奈又被称为“桃夹李”。木奈果实果大肉厚、甜酸适口、营养丰富、可食率高,深受消费者欢迎,是国内外市场畅销的名贵果品。但木奈果属于呼吸跃变型果实,其果实成熟于7月下旬至8月上旬,正值盛夏高溫时节,果实采后生理代谢旺盛,在常温下极易后熟软化和腐烂变质,引起极大的采后损失。因此,研究木奈果采后保鲜贮运具有重要的理论和实践意义。
1-甲基环丙烯(1-methylcyclopropene,简称1-MCP)是一种新型乙烯受体抑制剂,具有无毒、高效、安全等优点,目前被广泛应用于桃、杏等呼吸跃变型果实的保鲜,能有效控制其后熟软化和腐烂,延长果实保鲜期[1-2]。近年来的研究结果发现,1-MCP能够诱导桃、杏等果实的采后抗病性,有效降低采后病害和腐烂的发生[1-2]。例如,1-MCP能有效控制由扩展青霉(Penicillium expansum)引起的桃果实采后腐烂[1]、增强采后杏果实的抗病性而减少腐烂的发生[2]。但也有报道认为,1-MCP对葡萄、柑桔、草莓等果实采后病害和腐烂的控制效果不明显[3-5]。例如,1-MCP对控制由指状青霉(Penicillium digitatum)引起的采后葡萄果实腐烂没有显著影响;此外,1-MCP加速了由指状青霉(P. digitatum)引起的柑桔果实采后腐烂[4]和草莓果实采后病害[5]。由此看来,1-MCP对采后果实腐烂和病害的影响因果树品种不同而异。前期的研究结果表明,1-MCP处理可减少‘油木奈果实腐烂[6]。目前还未见有关1-MCP控制采后木奈果实腐烂与抗病相关酶诱导关系的相关研究报道。笔者以福建省主栽品种‘油木奈(Prunus salicina Lindl. cv. Younai)果实为材料,研究1-MCP处理对采后‘油木奈果实腐烂及抗病相关酶活性的影响,以期为生产上应用1-MCP处理控制采后‘油木奈果实腐烂、延长其保鲜期提供科学依据和技术参考。
1 材料与方法
1.1 材料及处理
供试‘油木奈果实采自福建省古田县科技示范果园,在果实约9成熟时采收,当天运至福建农林大学农产品产后技术研究所食品贮藏保鲜实验室。试验所用果实要求大小均匀、果形端正、色泽一致、无机械伤和病虫害。先用60 mg/L的ClO2溶液浸泡5 min对果实进行杀菌消毒,晾干后进行以下处理:(1)1-MCP处理:本试验所用1-MCP为纸片型的安喜布,由台湾利统股份有限公司提供。前期研究结果表明,25 ℃时1-MCP处理‘油木奈果实适宜的浓度为1.2 μL/L,时间为12 h[6]。因此,本试验选取1-MCP的处理浓度为1.2 μL/L。果实装入体积约0.04 m3的泡沫箱后,根据处理浓度裁取适宜大小的纸片型1-MCP,将其用蒸馏水喷湿后平铺于果实上,迅速将泡沫箱密封,并在(25±1)℃下处理12 h。(2)对照(CK):果实放入体积约0.04 m3的泡沫箱内,在(25±1)℃下密闭12 h。处理好的果实用0.015 mm厚的聚乙烯薄膜袋包装,每处理重复3次,每个处理50袋,每袋装果10个,之后在(25±1)℃,相对湿度90%条件下贮藏,贮藏期间每隔3 d取样观察果实腐烂率和测定相关指标。
1.2 测定项目与方法
1.2.1 腐烂率的测定 贮藏期间定期随机取样2袋(20个果实),通过肉眼观察评价‘油木奈果实贮藏期间的腐烂率。当果实可见腐烂区域超过1 mm宽,就被认为是腐烂果,腐烂率/%=(腐烂果数/贮前总果实数)×100。
1.2.2 几丁质酶活性的测定 参照Zheng等[7]的方法测定几丁质酶活性。
1.2.3 β-1,3-葡聚糖酶活性的测定 参照Zheng等[7]的方法测定β-1,3-葡聚糖酶活性。
1.2.4 苯丙氨酸解氨酶(PAL)活性的测定 参照曹建康等[8]的方法测定苯丙氨酸解氨酶(PAL)活性。
1.2.5 多酚氧化酶(PPO)活性的测定 参照Yang等[9]的方法测定多酚氧化酶(PPO)活性。
1.2.6 过氧化物酶(POD)活性的测定 参照Yang等[9]的方法测定过氧化物酶(POD)活性。
1.3 数据分析
各指标测定均重复3次,数据采用SPSS 16.0 数据分析软件进行方差分析(ANOVA)和Duncan多重比较法进行差异显著性分析。
2 结果与分析
2.1 1-MCP处理对‘油木奈果实腐烂率的影响
由图1可知,‘油木奈果实的腐烂率随采后贮藏时间的延长而增加,但不同处理的变化幅度不同。其中,对照和经1-MCP处理的果实在贮藏0~3 d内都保持完好,第6天开始出现不同程度的腐烂,贮藏9~18 d内,对照果实腐烂率快速增加,而经1-MCP处理的果实较快增加。进一步比较发现,在贮藏6~18 d内的同一贮藏时期,经1-MCP处理的果实腐烂率都低于对照果实。如贮藏至18 d时,经1-MCP处理的果实腐烂率为32%,而对照果实的腐烂率为52%,两者间差异显著(p<0.05)。以上研究结果表明,1-MCP处理能有效降低贮藏期间果实的腐烂。
2.2 1-MCP处理对‘油木奈果实几丁质酶活性的影响
由图2可知,贮藏0~15 d内,对照和经1-MCP处理的‘油木奈果实几丁质酶变化趋势基本一致。其中,贮藏0~3 d内快速下降,3 d时酶活性分别降至0 d时的47.1%和64.4%,3~9 d内较快上升,并在第9天时活性达到高峰,此时1-MCP处理果实几丁质酶活性比对照高出10.6%,9~12 d内又快速下降,12~15 d内有所回升,15 d后对照果实的几丁质酶活性有所下降,而经1-MCP处理的果实几丁质酶活性急速升高。进一步比较发现,在整个贮藏期间的同一贮藏时期,经1-MCP处理的果实几丁质酶活性均高于对照果实,且二者之间差异显著(p<0.05)。以上研究结果表明,1-MCP处理诱导了‘油木奈果实贮藏期间几丁质酶的活性,增强了抗病性。
2.3 1-MCP处理对‘油木奈果实β-1,3-葡聚糖酶活性的影响
由图3可知,‘油木奈果实β-1,3-葡聚糖酶活性在采后贮藏期间总体呈先上升后下降的趋势,都在贮藏第12天时活性达到高峰。进一步比较发现,在整个贮藏期间的同一贮藏时期,经1-MCP处理的果实β-1,3-葡聚糖酶活性均高于对照果实。如贮藏至12 d时,对照和经1-MCP处理的果实β-1,3葡聚糖酶活性分别为3.60 U/g FW和4.15 U/g FW,二者间差异显著(p<0.05)。以上研究结果表明,1-MCP處理诱导了‘油木奈果实贮藏期间β-1,3-葡聚糖酶的活性。
2.4 1-MCP处理对‘油木奈果实PAL活性的影响
由图4可知,对照和经1-MCP处理的果实PAL活性在贮藏期间变化趋势基本一致,总体呈下降、上升再下降的趋势。其中,对照果实PAL活性在贮藏0~3 d内快速下降,3~6 d内有所回升,6~12 d内快速下降,并在第12天时降到最低值,此时酶活性为初始值的25.69%,12~15 d内急剧升高至约0 d时的水平,之后又快速下降。而经1-MCP处理的果实PAL活性在贮藏0~3 d内快速下降,3~9 d内较快下降,9 d时降至最低值,此时酶活性为0 d时的19.42%,9~12 d内略有上升,12~15 d内迅速升高,并在第15天时达到峰值,此时酶活性为同期对照的62.56%,之后快速下降。进一步比较发现,经1-MCP处理的果实PAL活性显著(p<0.05)低于对照果实,说明1-MCP处理抑制了‘油木奈果实贮藏期间PAL的活性。
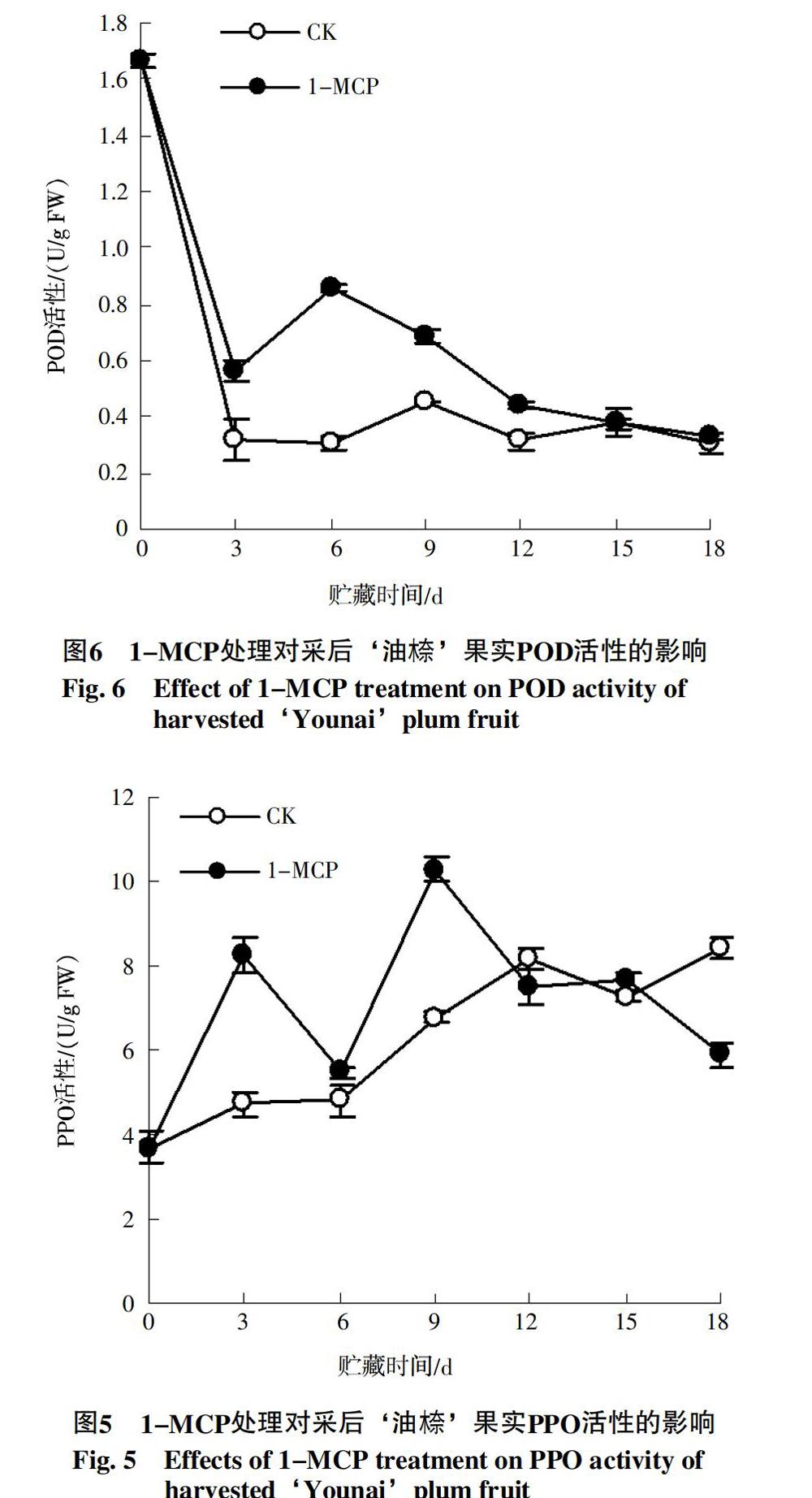
2.5 1-MCP处理对‘油木奈果实PPO活性的影响
由图5可知,对照果实PPO活性在贮藏0~3 d内较快上升,3~6 d内变化不大,6~12 d内持续快速上升,12~15 d内较快下降,之后回升至12 d时的水平。而经1-MCP处理的果实PPO活性在贮藏至3 d和9 d时出现酶活性高峰,分别为同期对照的1.75和1.52倍,9~12 d内快速下降,12~15 d内基本不变,之后快速下降,18 d时PPO活性为同期对照果实的70%。进一步比较发现,经1-MCP处理的果实PPO活性在贮藏前期(0~9 d)高于对照果实,说明1-MCP处理对PPO具有一定的激活作用。
2.6 1-MCP处理对‘油木奈果实POD活性的影响
由图6可知,‘油木奈果实POD活性随贮藏时间的延长总体呈下降趋势。其中对照果实POD活性在0~3 d内急剧下降,3~6 d内变化不大,6~9 d内有所上升,9~12 d内缓慢下降,之后变化不大。而1-MCP处理的果实POD活性在0~3 d内急剧下降,3~6 d快速上升,在贮藏第6 天时POD活性达到一个小高峰,之后持续下降。进一步比较发现,在贮藏3~12 d内,经1-MCP处理的果实POD活性均高于对照果实,且二者间差异显著(p<0.05),说明1-MCP处理诱导了‘油木奈果实贮藏前期POD活性的升高。
3 讨论与结论
果蔬遭受病原菌的侵染后,并非被动接受,而是会积极作出一系列的防卫反应,通过多种方式抵御病原物的侵染,包括抗菌物质(如酚类)和植保素的合成和积累、参与酚类物质合成的PAL、POD等酶的诱导合成及病程相关蛋白,如几丁质酶、β-1,3-葡聚糖酶活性的增强等。
病程相关蛋白是植物遭受病原菌侵染、机械伤或逆境胁迫刺激后产生的一类防御蛋白。几丁质酶和β-1,3-葡聚糖酶分别属于PR-3和PR-2家族的病程相关蛋白,在高等植物体内普遍存在。几丁质酶和β-1,3葡聚糖酶活性的增高是植物遭受病原物侵染后表现出的一种抗性反应。几丁质酶和β-1,3-葡聚糖酶可以直接作用于病原物真菌,降解细胞壁物质几丁质和β-1,3-葡聚糖,破坏其结构,所以具有直接的抗菌作用。前人的研究结果表明,几丁质酶和β-1,3-葡聚糖酶在枇杷、杨梅和芒果果实的采后抗病性中起重要作用[10-12]。本研究结果表明,1-MCP处理可诱导‘油木奈果实几丁质酶和β-1,3-葡聚糖酶活性的升高,且在贮藏期间一直保持在较高水平(图2、图3),同时果实的腐烂指数显著低于对照(图1),说明1-MCP控制‘油木奈果实采后腐烂与诱导果实几丁质酶和β-1,3葡聚糖酶活性的升高密切相关。病程相关蛋白在细胞中的分布不同,通常情况下,酸性蛋白位于细胞间隙中,而几丁质酶和β-1,3-葡聚糖酶等碱性蛋白主要位于液泡中。贮藏后期由于细胞衰老或病原菌的侵染等细胞膜结构的破坏,细胞结构的区室化结构丧失,位于液泡内的几丁质酶和β-1,3-葡聚糖酶可释放出来,直接对病原真菌细胞壁发生降解作用,而病原物侵染刺激可诱导几丁质酶和β-1,3-葡聚糖酶活性的升高,这可能是贮藏后期对照和1-MCP处理果实几丁质酶和β-1,3-葡聚糖酶活性升高的原因。
酚类物质是植物体内普遍存在的一类重要的次生代谢产物,也是植物体内的天然抗病性物质,能够抑制病原菌的侵染。酚类物质的合成主要通过莽草酸途径和苯丙烷途径。PAL是苯丙烷代谢途径的第一个酶,也是关键酶和限速酶。PAL参与酚类、植保素和木质素等抗病性物质的生物合成,因此PAL活性可以反映植物的抗病性。前人的研究结果表明,果实抗病性的增加与PAL活性增强有关[13-15]。但本研究结果发现,1-MCP处理降低了PAL活性(图4),这可能是由于PAL的产生某种程度上与乙烯作用有关。衰老或逆境胁迫下,外源或内源乙烯的产生可能会增强PAL活性[16],而1-MCP处理降低了乙烯的生成,这可能导致了PAL活性较低。此外,PAL是酚类物质生物合成的关键酶,当植物组织遭受了机械损伤或微生物侵染,PAL活性会增加,PAL活性较低也意味着1-MCP处理降低了‘油木奈果实的发病率。
PPO和POD都是植物体内重要的防御反应相关酶,它们都能被病原微生物侵染所诱导。PPO能将一些酚类物质氧化生成醌类物质,醌类物质再经非酶聚合反应形成黑色素等深色物质,对入侵的微生物有高毒性。在H2O2存在的条件下,POD也能氧化酚形成醌,参与黑色素物质的生成。此外,POD还是抗病性物质木质素和植保素合成的关键酶。可见,PPO和POD在植物的抗病性中起着重要作用。前人研究结果表明,PPO和POD活性的增加可增强果实的抗病性[17-18]。本研究结果发现,1-MCP处理可诱导‘油木奈果实PPO和POD活性的升高,且在贮藏期间一直保持在较高水平(图5、图6),说明1-MCP控制‘油木奈果实采后腐烂与诱导果实PPO和POD活性上升也有密切相关。
综上所述,果实的抗病性与后熟衰老密切相关,1-MCP处理可以有效降低‘油木奈果实的腐烂,1-MCP诱导的抗病性可能与几丁质酶、β-1,3-葡聚糖酶、PPO和POD等抗病相关酶活性的增加及1-MCP通过抑制乙烯延缓果实的后熟衰老,从而保持较高的对病原菌的防御能力有关,诱导和提高抗病性是1-MCP处理控制‘油木奈果实采后腐烂的重要作用机理之一。
参考文献
[1] Liu H X, Jiang W B, Zhou L G, et al. The effects of 1-methylcyclopropene on peach fruit(Prunus persica L. cv. Jiubao)ripening and disease resistance[J]. International Journal of Food Science & Technology, 2005, 40(1): 1-7.
[2] Dong L, Lurie S, Zhou H W. Effect of 1-methylcyclopropene on ripening of ‘Canino apricots and ‘Royal Zee plums[J]. Postharvest Biology and Technology, 2002, 24(2): 135-145.
[3] Mullins E D, McCollum T G, McDonald R E. Consequences on ethylene metabolism of inactivating the ethylene receptors sites in diseased non-climacteric fruit[J]. Postharvest Biology and Technology, 2000, 19(2): 155-164.
[4] Marcos J F, Gonzalez-Candelas L, Zacarias L. Involvement of ethylene biosynthesis and perception in the susceptibility of citrus fruits to Penicillium digitatum infection and the accumulation of defence-related mRNAs[J]. Journal of Experimental Botany, 2005, 56(418): 2 183-2 193.
[5] Jiang Y M, Joyce D C, Terry L A. 1-Mehtylcyclopronpene treatment affects strawberry fruit decay[J]. Postharvest Biology and Technology, 2001, 23(3): 227-232.
[6] 李 輝, 林河通, 袁 芳, 等. 不同浓度1-MCP处理对采后油木奈果实的保鲜效应[J]. 农业机械学报, 2012, 43(5): 114-121.
[7] Zheng Y, Shen L, Yu M M, et al. Nitric oxide synthase as a postharvest response in pathogen resistance of tomato fruit[J]. Postharvest Biology and Technology, 2011, 60(1): 38-46.
[8] 曹建康, 姜微波. 采后ASM诱导处理对鸭梨果实黑霉病的控制[J]. 园艺学报, 2005, 32(5): 783-787.
[9] Yang Z F, Cao S F, Cai Y T, et al. Combination of salicylic acid and ultrasound to control postharvest blue mold caused by Penicillium expansum in peach fruit[J]. Innovative Food Science & Emerging Technologies, 2011, 12(3): 310-314.
[10] Cao S F, Zheng Y H. Effect of 1-methylcyclopropene on anthracnose rot caused by Colletotrichum acutatum and disease resistance in loquat fruit[J]. Journal of the Science of Food and Agriculture, 2010, 90(13): 2 289-2 294.
[11] Wang K T, Cao S F, Jin P, et al. Effect of hot air treatment on postharvest mould decay in Chinese bayberry fruit and the possible mechanisms[J]. International Journal of Food Microbiology, 2010, 141(1-2): 11-16.
[12] Hu M J, Yang D P, Huber D J, et al. Reduction of postharvest anthracnose and enhancement of disease resistance in ripening mango fruit by nitric oxide treatment[J]. Postharvest Biology and Technology, 2014, 97: 115-122.
[13] Zhang Z Q, Tian S P, Zhu Z, et al. Effects of 1-methylcyclopropene(1-MCP) on ripening and resistance of jujube(Zizyphus jujuba cv. Huping)fruit against postharvest disease[J]. LWT-Food Science and Technology, 2012, 45(1): 13-19.
[14] Aghdam M S, Dokhanieh A Y, Hassanpour H, et al. Enhancement of antioxidant capacity of cornelian cherry(Cornus mas)fruit by postharvest calcium treatment[J]. Scientia Horticulturae, 2013, 61: 160-164.
[15] Shao X F, Wang H F, Xu F, et al. Effects and possible mechanisms of tea tree oil vapor treatment on the main disease in postharvest strawberry fruit[J]. Postharvest Biology and Technology, 2013, 77: 94-101.
[16] Lafuente M T, Zacarias L, Martinez-Tellez M A, et al. Phenylalanine ammonia-lyase as related to ethylene in the development of chilling symptoms during cold storage of citrus fruits[J]. Journal of Agricultural and Food Chemistry, 2001, 49(12): 6 020-6 025.
[17] Jin P, Zheng Y H, Tang S S, et al. Enhancing disease resistance in peach fruit with methyl jasmonate[J]. Journal of the Science of Food and Agriculture, 2009, 89(5): 802-808.
[18] Yuan L, Bi Y, Ge Y H, et al. Postharvest hot water dipping reduces decay by inducing disease resistance and maintaining firmness in muskmelon(Cucumis melo L.)fruit[J]. Scientia Horticulturae, 2013, 161: 101-110.
猜你喜欢
中国当代医药(2020年9期)2020-04-16
医学食疗与健康(2019年9期)2019-09-10
安徽农业科学(2018年18期)2018-05-14
分析化学(2018年2期)2018-03-02
化学教与学(2017年7期)2017-07-18
江苏农业科学(2016年1期)2017-05-17
分析化学(2014年7期)2014-12-13
食品安全导刊(2014年7期)2014-10-21
现代农业科技(2009年19期)2009-03-20
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26
