
糖能兼用甘蔗主要性状的遗传参数与改良效果的分析
2015-10-21 19:04唐仕云王伦旺杨荣仲杨丽涛李翔黄海荣经艳邓宇驰谭芳黄家雍
热带作物学报 2015年4期
关键词:甘蔗
唐仕云 王伦旺 杨荣仲 杨丽涛 李翔 黄海荣 经艳 邓宇驰 谭芳 黄家雍

摘 要 为了解糖能兼用甘蔗主要性状的遗传变异、选择和改良的效果,以自育的14个新品系以及双对照新台糖22号和新台糖16号为参试材料,进行了1新1宿品比试验。结果表明:糖能兼用甘蔗新品系在新植和宿根的工艺成熟期,株高、茎径、公顷有效茎数、锤度、公顷甘蔗产量、公顷产糖量、公顷总可发酵糖量,新植总苗数、宿根发株率和宿根黑穗病发病率均具有显著或极显著的差异。公顷甘蔗产量、公顷产糖量、公顷总可发酵糖量均具有较高的广义遗传率和较大的遗传变异系数,说明受环境的影响小,对产量性状直接选择效果好。表型和遗传相关分析表明,新植和宿根蔗的公顷有效茎数与公顷甘蔗产量、公顷产糖量、公顷总可发酵糖量均具有很高的正相关关系。以公顷产糖量作为糖料蔗品种的选择性状时,8个新品系超过双对照而被入选;以公顷总可发酵糖量作为糖能兼用甘蔗品种的选择性状时,6个新品系超过双对照而被入选。这2种选择方法所入选的新品系类型基本相同 ,并且公顷有效茎数、公顷甘蔗产量、公顷产糖量和公顷总可发酵糖量都具有较大的遗传增益。本研究有望为糖能兼用甘蔗新品种选育提供理论参考和依据。
关键词 甘蔗;糖能兼用; 遗传参数;遗传增益
中图分类号 S566.1 文献标识码 A
甘蔗是重要的糖料作物和能源作物,广泛种植于热带和亚热带地区。由于不可再生能源终将枯竭,开发可再生生物质能源变得越来越必要和重要。巴西和美国十分重视甘蔗生物质能源的开发,巴西是世界上最大的应用甘蔗生产燃料乙醇的国家,用于生产酒精的甘蔗已经超过用于制糖的甘蔗。中国是世界上第三大甘蔗和食糖生产大国,发展能源甘蔗具有巨大的潜在优势[1-3]。
品种是甘蔗种植的重要物质基础,选育优良的品种是甘蔗生产燃料乙醇的核心技术,开展能源或糖能兼用甘蔗新品种的选育对我国可再生能源事业的发展具有重要的经济意义和社会意义[4]。目前国内多家甘蔗育种单位都开展了能源或糖能兼用甘蔗种质资源收集利用和新品种选育的研究工作[5-7],并已成功选育出了不少优良新品种[4,8-11]。糖能兼用甘蔗品种以总生物量、总可发酵糖量为育种目标,对生物产量和可发酵糖产量要求较高,同时蔗茎蔗糖分和重力纯度能满足制糖工艺的要求,而且甘蔗纤维分、蔗茎产量和蔗糖分必须达到国家糖料甘蔗品种审(鉴)定的标准,糖能兼用甘蔗品种的主要经济技术指标与糖料蔗的主要经济技术指标相近[12]。但是,甘蔗常规杂交育种的周期长,可预见性差,育种的成效并不很显著,尤其是关于能源或糖能兼用甘蔗,我国的研究起步较晚[1,13],新品种选育所积累的经验相对糖料甘蔗较少,对各性状的遗传规律及选择效应的评价有待完善。
本研究应用数理统计的方法,对糖能兼用甘蔗新品种的主要性状进行了遗传参数和遗传改良效应分析,以期为糖能兼用甘蔗新品种选育提供理论参考,进而选育出更多更好的甘蔗新品种,更好地为整个甘蔗产业生产服务。
1 材料与方法
1.1 材料
以广西农科院甘蔗研究所2008年度部分实生苗材料经常规育种选育而来的14个新品系,分别为GT08-88、GT08-221、GT08-278、GT08-297、GT08-460、GT08-921、GT08-1045、GT08-1180、GT08-1235、GT08-1314、GT08-1409、GT08-1533、GT08-1589、GT08-1628,作为试验参试材料,以新台糖22号(ROC22)和新台糖16号(ROC16)为双对照。
1.2 方法
1.2.1 试验设计及管理 2012~2014年,在常规育种的品比阶段,在广西南宁市隆安县广西农业科学院甘蔗研究所叮当基地进行了1年新植和1年宿根的品比试验。采用完全随机区组设计,重复3次,每小区4行区,行长6 m,行距1.2 m,试验于2012年3月28~30日种植,下种量10.5万芽/hm2,2013年3月19日砍收,之后进行宿根蔗破垄松蔸施肥等管理,所有的田间管理与一般大田生产相同,2014年2月28日砍收宿根。
1.2.2 性状调查 调查的性状包括新植蔗苗数高峰期时的总苗数,宿根蔗的发株率、黑穗病发病率、6月底株高,新植和宿根蔗的年底株高、茎径、公顷有效茎数(MSH)、田间锤度、公顷甘蔗产量(TCH)、甘蔗蔗糖分、纤维分。每年的11月至翌年2月,每个月每个品种(系)取6条甘蔗委托农业部甘蔗及制品检测中心(南宁)化验分析甘蔗蔗糖分与纤维分,其它所有性状均按甘蔗常规调查方法进行,公顷产糖量(TSH)=公顷甘蔗產量(TCH)×甘蔗蔗糖分;公顷总可发酵糖量(TFH)参照王伦旺等[11]的方法:TFH=TCH×(甘蔗纤维分+甘蔗锤度)。
1.2.3 数据处理与统计分析 统计分析:试验按单因素安全随机区组设计进行方差分析,以小区为单位,宿根黑穗病发病率采用反正弦转化,其它性状不做转化,按参考文献[14]介绍的方法进行。广义遗传力、表型和遗传相关系数按刘来福[15]方法进行,hB2=100×VG/VP=100×(V1-V2)/[V1+(r-1)V2],其中,hB2、VG、VP、V1、V2、r分别为广义遗传率、基因型方差、表现型方差、品种(系)间方差、机误、重复次数。表型变异系数(PCV)和遗传变异系数(GCV)按马育华[16]、李玉潜[17]方法进行,PCV(%)=(σp/X)×100%,GCV(%)=(σp/X)×100%,σp、σg、X分别为表型标准差、遗传型标准差和群体均值。选择响应按孔繁玲[18]的方法进行,选择响应R=hB2×S,hB2为广义遗传力,S为选择差,S=X1-X, X1为某性状入选个体的平均值,X为群体的性状平均值。遗传增益ΔG按刘忠松[19]的方法进行,ΔG=R/X×100%,遗传增益为选择响应除以群体平均值所得的百分率。所有数据处理和参数分析采用Excel和DPS系统软件[20]分析。
2 结果与分析
2.1 糖能兼用甘蔗主要性状的遗传变异及广义遗传力分析
从表1可以看出,所参试的16个品种(系)材料,除6月底株高外,其它性状的F值均达显著或极显著水平,说明除6月底株高外,其它各性状不同参试材料间具有显著或极显著的差异,为进一步测定遗传力等遗传参数和甘蔗新品种的选择及选择效应分析提供了遗传基础,具有改良的潜力。
从变异系数来看,年底株高、茎径、公顷有效茎数、锤度、公顷甘蔗产量、公顷产糖量、公顷总可发酵糖量的表型和遗传变异系数在新植蔗和宿根蔗之间相差不大,并且变化趋势也相同,说明分别对新植和宿根的这些性状进行选择时,将具有大体相同的效应。表型和遗传变异系数最大的性状是宿根黑穗病发病率,新植总苗数、宿根發株率、新宿公顷有效茎数、新宿公顷甘蔗产量、新宿公顷产糖量、新宿公顷总可发酵糖量的表型和遗传变异系数均较高,而新宿株高、新宿茎径和新宿锤度的表型和遗传变异系数均较低。
从性状遗传力的大小来看,宿根黑穗病发病率、新宿茎径、新宿锤度、新宿公顷甘蔗产量、新宿公顷产糖量和新宿公顷总可发酵糖量具有较高广义遗传率,广义遗传率均达50%以上。新宿年底株高和新宿公顷有效茎数的遗传力居中,广义遗传率在35%~65%之间。而新植总苗数、宿根发株率、宿根6月株高的遗传力较低,广义遗传率在35%以下。
由变异系数和遗传力分析可知,无论新植还是宿根,公顷甘蔗产量、公顷产糖量和公顷总可发酵糖量均具有较大的变异系数和较高的广义遗传率,直接选择的效果较好。而宿根6月株高的变异系数和广义遗传率都很低,选择效果差,不能作为选择性状。
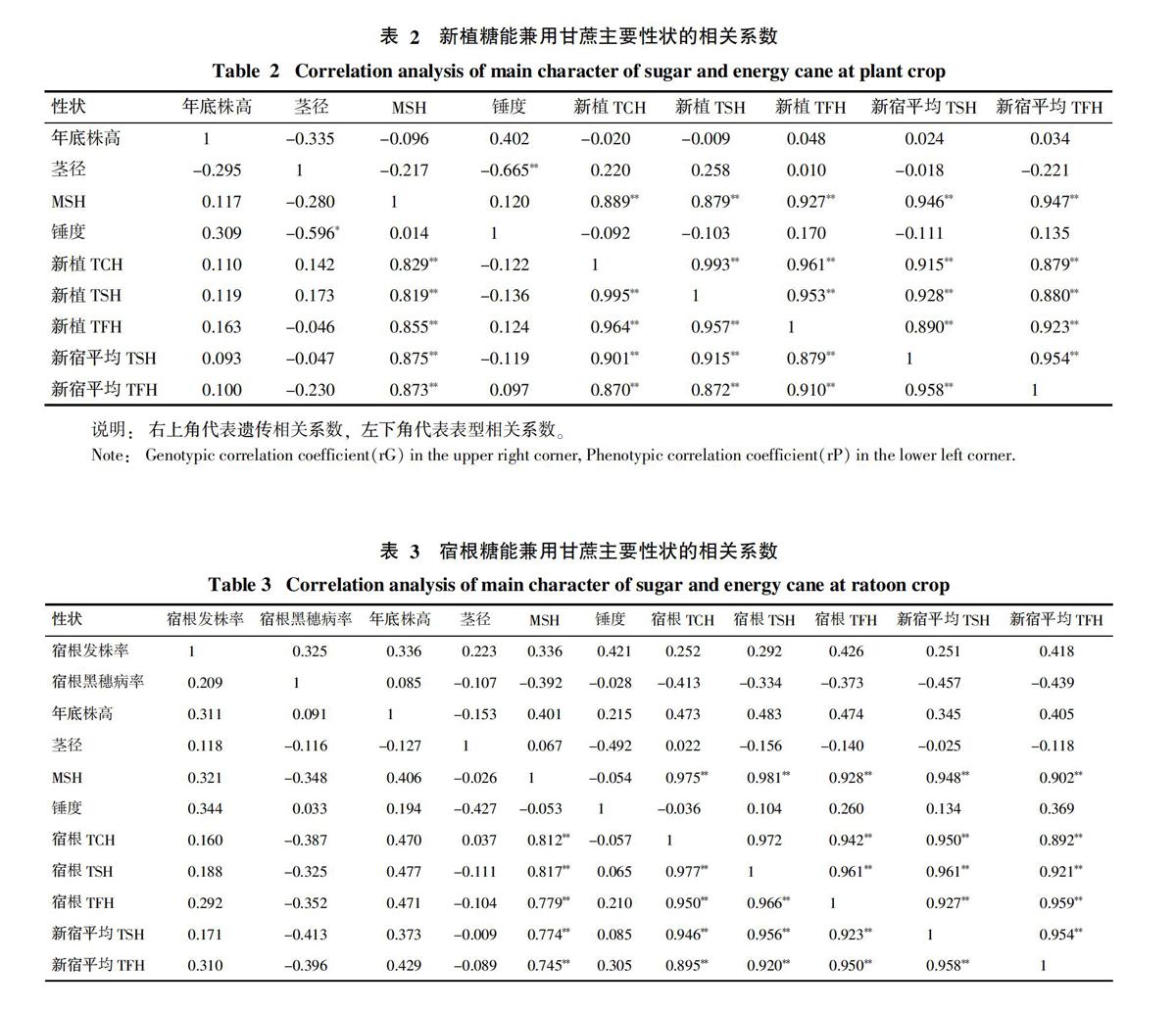
2.2 糖能兼用甘蔗主要性状的相关分析
从表2可以看出,在新植蔗时,16个参试材料的公顷有效茎数与新植公顷甘蔗产量、新植公顷产糖量、新植公顷总可发酵糖量、新宿平均公顷产糖量和新宿平均公顷总可发酵糖量都具有极高的表型和遗传正相关系数,其次是锤度与茎径间存在较高的表型和遗传负相关关系。年底株高、茎径、锤度与新植公顷甘蔗产量、新植公顷产糖量、新植公顷总可发酵糖量、新宿平均公顷产糖量和新宿平均公顷总可发酵糖量之间的表型和遗传相关系数都很小,说明通过新植年底株高、茎径和锤度的选择对促进本次试验的甘蔗产量、糖产量和总可发酵糖产量的效果不大。
从表3可以看出,在宿根蔗时,各性状间的相关关系与新植蔗时的变化趋势大体相同,公顷有效茎数与宿根公顷甘蔗产量、宿根公顷产糖量、宿根公顷总可发酵糖量、新宿平均公顷产糖量和新宿平均公顷总可发酵糖量都具有极高的表型和遗传正相关系数,茎径与锤度间具有较高的表型和遗传负相关关系。年底株高与宿根公顷甘蔗产量、宿根公顷产糖量、宿根公顷总可发酵糖量、新宿平均公顷产糖量和新宿平均公顷总可发酵糖量具有较高的表型和遗传正相关系数,比新植蔗时相关性要高。而锤度、茎径、宿根发株率与宿根公顷甘蔗产量、宿根公顷产糖量、宿根公顷总可发酵糖量、新宿平均公顷产糖量和新宿平均公顷总可发酵糖量的表型和遗传相关系数较小,而宿根黑穗病发病率与宿根公顷甘蔗产量、宿根公顷产糖量、宿根公顷总可发酵糖量、新宿平均公顷产糖量和新宿平均公顷总可发酵糖量具有较高的表型和遗传负相关系数,说明黑穗病是导致各种产量减产的重要原因之一。
综合新植和宿根蔗的各性状相关关系来看,公顷有效茎数对公顷甘蔗产量、公顷产糖量、公顷总可发酵糖量都具有很高的表型和遗传正相关系数,因而,在本次试验中,对各产量目标性状进行间接选择时,公顷有效茎数可以作为重要的间接辅助选择性状加以利用。
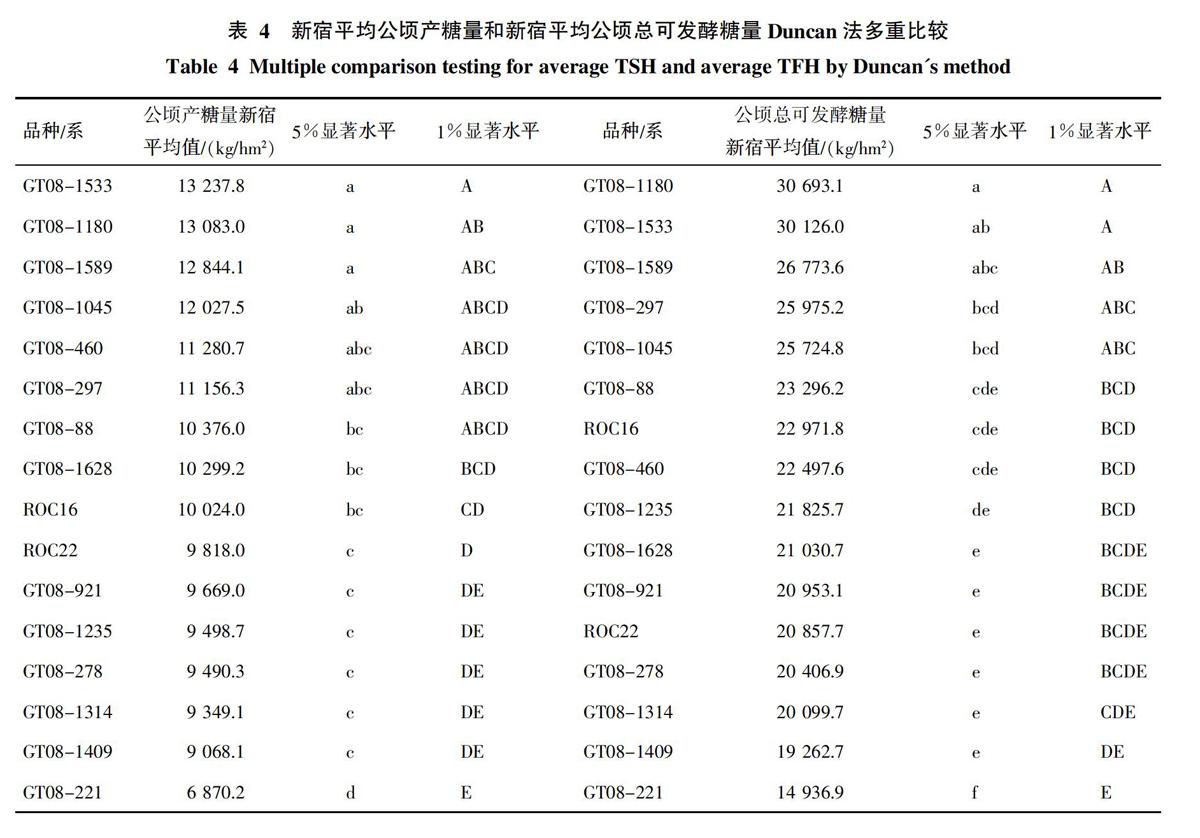
2.3 糖能兼用甘蔗选择性状的多重比较及选择
表4列出了14个参试新品系和2个对照品种的平均值及多重比较的结果,可以看出,若以新宿平均公顷产糖量为选择性状时,有GT08-1533、GT08-1180、GT08-1589、GT08-1045、GT08-460、GT08-297、GT08-88、GT08-1628共8个新品系的平均值超过双对照,以超过双对照的品系为入选材料,则入选率为50%。
若以新宿平均公顷总可发酵糖量为选择性状时,有GT08-1180、GT08-1533、GT08-1589、GT08-297、GT08-1045、GT08-88共6个新品系超过双对照,入选率为37.5%,与以新宿平均公顷产糖量为选择性状相比,仅GT08-460、GT08-1628没有入选,因而,这两种选择方法所入选的品系数量和类型总体上差别不太大,两种选择方法间并不矛盾。
2.4 糖能兼用甘蔗不同选择方法遗传增益大小的估计
从表5可知,除新宿株高、新宿茎径和宿根黑穗病发病率以新宿平均公顷产糖量为选择性状时的遗传增益比以新宿平均公顷总可发酵糖量为选择性状时的遗传增益略大以外,其余性状均以新宿平均公顷总可发酵糖量为选择性状时的遗传增益大。各性状遗传增益大小的变化趋势在两种选择方法上一致,表现为新宿公顷甘蔗产量、新宿公顷产糖量和新宿公顷总可发酵糖量均有较大的遗传增益,其次是新宿公顷有效茎数和新植总苗数,而宿根黑穗病率的遗传增益最小,为负值。说明以新宿平均公顷产糖量和新宿平均公顷总可发酵糖量为选择性状时,均能有效提高甘蔗新品种(系)群体的新宿公顷甘蔗产量、新宿公顷产糖量、新宿公顷总可发酵糖量、新宿公顷有效茎数、新植总苗数,降低宿根黑穗病率,而对新宿株高、新宿茎径、新宿锤度的改进不大。
3 讨论与结论
优良新品种的选育是降低生产成本、提高产业竞争力最为有效的措施之一。能源甘蔗以高生物量、高可发酵糖含量为选育目标,又可分为能源专用和糖能兼用2种类型,糖能兼用甘蔗进入工艺成熟期,蔗糖分和重力纯度能满足制糖工艺要求,可生产蔗糖或乙醇[21]。因而,能源甘蔗品种的选育方法与糖料蔗新品种的选育程序并没有本质的差别[12]。本研究对新宿平均公顷产糖量与新宿平均公顷总可发酵糖量2种选择方法的改良效果的分析表明,各相关主要性状的遗传增益在2种选择方法中具有大体相同的变化趋势,因而糖料甘蔗品种与糖能兼用甘蔗品种的选育并不矛盾。
遗传参数的估计在遗传改良和育种工作中具有重要的理论指导意义,遗传力反映群体中根据表型值的优劣选择基因型值的可靠程度[22-23],变异系数可以衡量各性状的选择潜力[24]。本次试验中,公顷甘蔗产量、公顷产糖量和公顷总可发酵糖产量在新植蔗和宿根蔗中均具有较高的广义遗传力和遗传变异系数,因而,在甘蔗育种的品比阶段,对各产量性状进行直接选择效果较好,能取得较大的遗传进度,这与谭中文[25]、陈坚[26]、李杨瑞[27]、陈西文[28]等多人报道的一致。但对甘蔗的锤度品质性状而言,本研究的遗传增益较小甚至为负值,这与李玉潜[17]报道的一致,说明对锤度等品质性状的改良幅度有限,在现有的育种程序中所需改良的时间较短。从性状构成来看,株高、茎径和锤度的遗传变异系数都很小,尤其是6月株高,品系之间无显著差异,这可能与甘蔗育种的程序及选择方法有关,在杂种圃至预备品比圃,对甘蔗产量、产糖量等综合性状的选择多以间接选择为主,选择性状主要采用遗传力较高的株高、茎径、锤度和生势等性状,以性状独立淘汰法进行严格选择,只要有一个性状有明显缺陷的品系就会被淘汰,因而,到品比圃阶段时,这些性状在品系间差异较小,可进行再选择的空间有限。而单位面积有效茎数由于在低级圃时,不同品系的蔗种数量少且不一致,种植密度较难保持一致,给这一性状的选择带来了很大困难,从本次试验可以看出,尽管各品系的公顷有效茎数遗传变异系数较大,但广义遗传力一般,选择的增益并不是特别高,因而应适当加大田间小区、增加重复次数和加强田间管理,提高遗传力,以提高选择的效果。
从表型与遗传相关来看,公顷有效茎数与公顷甘蔗产量、公顷产糖量、公顷总可发酵糖产量均具有较高的相关性,这与谭中文[29]、李杨瑞[27]、龚德明[30]、黄家雍[31]等的报道相同。相关分析揭示了性状间关系的密切程度,可根据相关系数进行性状的间接选择[32-33]。从本次试验可以看出,公顷有效茎数在常规育种后期,作为产量性状的间接选择指标,具有重要的意义。在新植和宿根蔗中,锤度与茎径有较高的负相关,说明在品种选择中,容易选到中小茎而高糖的品种,但由于在目前手工砍收甘蔗的条件下,农民不喜欢小茎的甘蔗品种,因而不能过于追求高糖而把小茎品种选育上来。黑穗病是甘蔗生产中危害极严重的一种病害,从本试验的分析可以看出,黑穗病发病率与产量性状间有较高的负相关,通过对各种产量的选择,可以显著降低黑穗病的发生。
本次试验仅仅是一个试点的试验结果,且在新植蔗种植时,由于天气原因,土地整理困难,蔗种堆放的时间久,出苗时因干旱天气影响甘蔗出苗,后期因受到较强台风影响,甘蔗倒伏严重,因而,对遗传参数估计及试验结果的可靠性会产生一定的影响,试验所得结论还有待更进一步的试验验证。
参考文献
[1] 李杨瑞, 谭裕模, 李 松,等. 甘蔗作为生物能源作物的潜力分析[J]. 西南农业学报, 2006, 19(4): 742-746.
[2] Kostka G, Polzin C, Scharrer J. The future of sugar cane in (the)Peoples Republic of China and India-Supply constraints and expansion potential[J]. Applied Energy. 2009, 86: 100-107. [3] Nguyen T L T, Hermansen J E, Sagisaka M. Fossil energy savings potential of sugar cane bio-energy systems[J]. Applied Energy, 2009, 86: 132-139.
[4] 李 松, 游建華, 余坤兴, 等. 几个糖能型甘蔗新品种高产栽培比较试验[J]. 广西蔗糖, 2009(4): 3-5.
[5] 吴嘉云, 邓祖湖, 徐良年, 等. 能糖兼用甘蔗新品种的评价与利用[J]. 福建农林大学学报(自然科学版), 2009, 38(1): 1-5.
[6] 毛 钧, 陆 鑫, 刘新龙, 等. “甘蔗复合群”能源植物遗传育种研究进展[J]. 植物分类与资源学报, 2014, 36(1): 89-98.
[7] 李 昱, 李奇伟, 邓海华, 等. 我国能源植物概况与能源型甘蔗斑茅后代前景展望[J]. 甘蔗糖业, 2014(3): 51-58.
[8] 谢惠珏, 朱文华, 周正邦, 等. 10个糖能兼用甘蔗品种(系)的性状表现[J]. 贵州农业科学, 2014, 42(2): 149-151.
[9] 谭显平, 吕 达, 陈道德, 等. 优质高产早熟多抗糖能兼用甘蔗新品种川蔗27号(川糖99-8602)选育研究报告[J]. 甘蔗糖业, 2013(4): 4-19.
[10] 刘福业, 邓海华, 杨俊贤, 等. 糖能兼用甘蔗新品种粤糖96-86的选育与种性分析[J]. 种子, 2011, 30(6): 98-101.
[11] 王伦旺, 李 翔, 黄家雍, 等. 16个糖能兼用甘蔗品种(系)种性比较[J]. 中国农学通报, 2010, 26(17): 408-415.
[12] 李杨瑞. 现代甘蔗学[M]. 北京: 中国农业出版社, 2010: 521.
[13] 林彦铨, 何启钧, 邓 强, 等. 甘蔗高生物量性状表现的重演力分析[J]. 甘蔗, 1997, 4(4): 6-10.
[14] 盖钧镒. 试验统计方法[M]. 北京: 中国农业出版社, 2000: 227-232.
[15] 刘来福, 毛盛贤, 黄远樟. 作物数量遗传[M]. 北京: 农业出版社, 1984: 110-180.
[16] 马育华. 植物育种的数量遗传学基础[M]. 江苏: 江苏科学技术出版社, 1982: 300-302.
[17] 李玉潜. 甘蔗遗传参数的研究*产量和品质性状的基因型变异、广义遗传力和相对遗传进度[J]. 甘蔗糖业, 1983(8): 35, 41-42.
[18] 孔繁玲. 植物数量遗传学[M]. 北京: 中国农业出版社, 2006: 183-239.
[19] 刘忠松, 罗赫荣. 现代植物育种学[M]. 北京: 科学出版社, 2010: 274-283.
[20] 唐启义. DPS数据处理系统实验设计、 统计分析及数据挖掘[M]. 北京: 科学出版社, 2007.
[21] 范水生. 中国能源甘蔗——燃料乙醇的开发探析[J]. 世界农业, 2006(12): 51-54.
[22] 单保山. 遗传力的概念及其发展Ⅰ. 关于传统遗传力的几个问题[J]. 河北农业大学学报, 1988, 11(1): 39-44.
[23] 于福来, 赵志刚, 王文全, 等. 栽培甘草产量相关性状遗传潜力及选择方法[J]. 中药材, 2013, 36(10): 1 551-1 555.
[24] 李芳东, 乌云塔娜, 杜红岩. 杜仲高产胶无性系遗传参数估计与改良效果分析[J]. 林业科学研究, 2007, 20(4): 458-463.
[25] 譚中文, Martin F A. 甘蔗的遗传相关及与环境互作的研究[J]. 华南农业大学学报, 1987, 8(3): 47-52.
[26] 陈 坚. 甘蔗性状遗传参数的研究[J]. 江西农业科技. 1993(3): 14-16.
[27] 李杨瑞. 甘蔗引进品种产量性状的遗传、相关及选择指数[J]. 福建农学院学报, 1986, 15(4): 317-326.
[28] 陈西文, 陈勇生, 邓海华. 斑茅蔗BC1后代产量性状遗传分析[J]. 广东农业科学, 2011(5): 44-46.
[29] 谭中文, 谭启超, 梁耀祥. 甘蔗品种的遗传、 相关及其通径分析[J]. 甘蔗糖业(甘蔗分刊), 1982(4): 12-17.
[30] 龚得明, 林彦铨, 陈如凯. 甘蔗蔗茎产量、蔗糖分和蔗糖产量的通径分析[J]. 甘蔗糖业, 1992(4): 9-14.
[31] 黄家雍, 诸葛莹, 刘海斌, 等. 甘蔗主要性状的遗传相关及通径分析[J]. 甘蔗, 1999, 6(3): 6-9.
[32] 戴君惕, 杨 德, 尹世强, 等. 相关遗传力及其在育种上的应用[J]. 遗传学报, 1983, 10(5): 375-383.
[33] 李会霞, 王玉文, 田 岗, 等. 谷子一些数量性状的遗传相关分析及其在育种上的应用[J]. 甘肃农业科技, 2005(4): 12-14.
猜你喜欢
课堂内外(小学版)(2022年3期)2022-04-14
旅游(2020年2期)2020-04-29
小猕猴学习画刊(2019年8期)2019-09-16
文萃报·周二版(2019年20期)2019-09-10
广东第二课堂·小学(2019年1期)2019-03-06
小星星·阅读100分(低年级)(2018年11期)2018-12-04
上海故事(2018年7期)2018-07-09
作文·初中版(2018年4期)2018-05-22
广东第二课堂·初中(2018年4期)2018-05-08
小天使·三年级语数英综合(2017年10期)2017-11-20
