
工作记忆中后颞叶与其他脑区的同步效应
2015-09-18 06:52梅丹丹黄锐敏
中国生物医学工程学报 2015年2期
梅丹丹 闫 铮 黄锐敏
(华侨大学信息科学与工程学院,福建 厦门 361021)
工作记忆中后颞叶与其他脑区的同步效应
梅丹丹 闫 铮 黄锐敏*
(华侨大学信息科学与工程学院,福建 厦门 361021)
采用脑电图(EEG)相干分析法构建功能网络,目的在于研究人脑负责工作记忆(WM)的主要区域——大脑后颞叶在不同模式工作记忆过程中的作用地位和区域职能。为此设计两种实验范式,要求12名受试记忆分别表征WM的两个成分,即语音回路和视空间模板的可言辞表达的字母串和非言辞表达的图形。采集19导EEG,分别计算语音回路和视空间模板的相干系数矩阵,绘制功能网络。结果显示,与静息态相比,在语音回路和视空间模板中,后颞叶(T5、T6)与其他脑区的θ(4~7 Hz)相干明显增强(P<0.05),表明后颞叶与其他脑区的θ频段同步效应参与WM;此外,在语音回路、视空间模板的功能网络中,节点T5、T6的度差异显著,但都远高于其他节点,表明左、右后颞叶分别在语音回路、视空间模板WM中扮演着专家化区域的角色。
工作记忆;后颞叶皮层;相干同步;功能网络
引言
工作记忆(working memory,WM)的概念是1974年由Baddeley和Hitch在短时记忆基础上提出的。他们在报道中提出,工作记忆是对信息进行暂时加工和保存的容量有限的记忆系统,在表象、言语、学习、推理和解决问题的高级认知活动中起着非常重要的作用[1]。工作记忆可以分为中央执行系统(central executive)、语音回路(phonological loop)和视空间模板(visual-spatial sketch pad )3个部分。其中,中央执行系统负责工作记忆中的注意控制,负责指挥各类次级系统的活动;语音回路和视空间模板是两个平行的子系统,分别负责与语音和空间想象有关的信息加工与贮存。
目前国内外对于工作记忆脑机制的研究,大多集中在功能性磁共振成像(functional magnetic resonance imaging,fMRI)和事件相关电位(event-related potential,ERP)的研究[2]。fMRI的原理是基于神经元功能活动对局部氧耗量和脑血流影响程度不匹配所导致的局部分子磁场性质变化,并不是基本神经活动的直接反映[3];且fMRI的费用较高,对研究造成一定的不便。与之相比,脑交流电位(alternating current,AC)不仅是神经活动的直接反映,而且可操作性强、低耗费、普及度高,因此近年来ERP在该领域的应用日益增多[4]。该技术的突出特点是时间分辨率高,可以精确到毫秒级,因此可以对特定认知事件引发的脑电位进行实时性测量。但是,一些ERP成分的工作记忆内涵不是很明确,导致不同研究结果之间的分歧比较大;而且ERP研究侧重于对特定脑区进行独立的检测与分析,忽视了脑区活动之间的相互联系。然而,神经元不是独立工作的,它们组成了部件和网络,进而借由兴奋性和抑制性的突触连接相互影响。研究表明,神经元活动的同步功能参与了工作记忆,以此来支持相距较远的大脑各区域之间的相互作用[5]。同步,这里指的是各脑区振荡之间的相位同步,靠提高神经可塑性来促进神经的沟通行为。相位同步通过使数个突触激励在同一时间内到达突触后面的神经元来支持神经通信[6],因此可以运用功能网络来探讨工作记忆的机制[7,8]。笔者采集脑电图(electroencephalogram,EEG)作为信号源,使用相干分析方法量化两两信号的同步程度,再由得到相干系数构建网络连接矩阵,进而研究分析大脑主要皮层区域在工作记忆中的作用。
设计实验,让受试者执行延迟响应任务,记忆分别表征语音回路和视空间模板的可言辞表达和非言辞表达的刺激项目,对记录的数据进行处理分析。结果发现,后颞叶与其他脑区的θ频段(4~7 Hz)同步效应参与了工作记忆过程。此外,对比语音回路、视空间模板的功能网络发现,左、右后颞叶分别在语音回路和视空间模板中扮演着专家化区域的角色。
1 材料和方法
1.1实验设计
由于语音回路和视空间模板是两个平行的子系统,所以Sarnthein等将实验范式设计为两组,分别要求受试者在短时内记忆呈现的可用言辞表达和不可言辞表达的刺激项目并进行复述[5]。本实验对其实验范式进行了改进和补充,具体包括两个子实验。在实验1中,刺激项目为6~8个大写英文字母。首先,字母串在电脑屏幕上呈现6 s,供受试者认知(认知状态);接着,字母串消失,受试者记忆刚刚呈现的字母串,此段持续4 s(记忆状态);最后,要求受试者默写出记忆的内容,此段持续10 s(召回状态)。3个状态一组,作为刺激项目呈现一次,数量为6、7、8个字母的刺激项目分别呈现28次,实验流程如图1所示。对于实验2,刺激物由16宫格、空心圆和实心圆组成,“圆”图形随机出现在不同的格子中。再者,设定实心圆的数量(1、2或3)表征记忆负荷梯度,设置2个空心圆搭配,构成图形。实验流程及次数与实验1一致。英文字母串的可读性较高,比起其他符号字符,更便于受试者采用默读复述的方式进行记忆;16宫格和实、空心圆组合的简单图形可充分表征形状和空间位置,受试者更易采用“拍照”式的空间想象方式进行记忆。然而,因个人习惯和记忆技巧的差异,不排除有受试者依据字母形状的视觉信息进行加工与贮存,所以需采集足够多的样本,并使男女受试者的数量相当,这样用结构较为对称的较大样本总量均值,消除个别依据字母形状记忆的受试者的结果以及性别等无关因素对整体实验结果的干扰。综上所述,实验1和实验2可以分别用来研究WM的两个平行子系统,而两组实验作为一个整体又可以用来探讨WM的注意控制机制,可以全面地研究WM的机制。作为对照,每组实验开始部分,要求受试者注视白字幕黑背景的电脑屏幕(同记忆状态的刺激样式)2 min,受试者尽量使大脑处于无负荷状态(静息态)。

图1 语音回路(上)和视空间模板(下)任务中刺激呈现一次的流程Fig.1 Schematic drawing of an event on each trial of the verbal (Top) and visual-spatial (Bottom) working memory tasks
12位成年男女(年龄23±3岁,6男6女,均有英语基础)参与了实验,受试者均签署知情同意书。采用新拓放大器,19导联电极帽采集EEG,双侧耳垂作为参考,采样频率为100 Hz。对EEG进行35 Hz低通滤波,去混叠,去工频干扰。
1.2EEG相干
这里指的“同步”是不同脑区振荡之间的相位同步。相位同步的量化方法在各研究成果中已经被大量描述[9-10]。这里采用的量化方法为相干分析法,因为这种时频分析方法算法简单,物理含义明确,又是对时域和频域测量技术的补充和完善[11]。具体方法是对每个独立时间序列做傅里叶变换得到各自的自功率谱Pxx(f)、Pyy(f),再进行互傅里叶变换得到互谱密度函数Pxy(f),归一化互谱密度函数得到相干系数Cxy(f)。相干系数表示两导信号间的振幅、频率、相角的一致性程度,以此来量化说明它们之间的同步关系。相干系数计算如下:

(1)
Cxy(f)各元素的值在0~1之间,它表征目标频率的相位分散程度。Cxy(f)=0意味着引脚之间的信号在该频率相位上是完全分散的、不相干的;Cxy(f)=1则表示信号x和信号y完全同步,在该频率相位完全锁定。概括来说,“同步”描述的是信号的相位同步,而相位同步用相干分析法来量化,所以相干系数增大,就意味着信号的同步性增强。
对预处理后的EEG信号做2 s傅里叶变换,求平均功率谱和每对导联(171对)的互功率谱,由式(1)求单次任务的f-C关系,再分别求静息态、记忆字母串、记忆图片状态的12人平均f-C关系。
1.3构建网络
将上述f-C关系在δ (2~4 Hz)、θ(4~7 Hz)、α1(7~10 Hz)、α2(10~13 Hz)、β(13~19 Hz)各波段求平均,分别得到19×19的静息态、记忆字母串和记忆图片状态的各波段平均相干系数矩阵Cc、Cve和Cvi。Cve、Cvi分别与Cc做差,得到ΔCve和ΔCvi。将其建立成连接图,构建二值矩阵ΔB,有
(2)
式中,T为阈值,在各波段计算、选取各自合适的值,使得连接图的网络密度小于20%,分别得到ΔBve和ΔBvi,即得到语音回路和视空间模板基于各波段相干的功能网络。
1.4基于权值和度的网络定量分析
在得到以上功能网络的基础上,为考察网络连接的强度和节点的作用地位,笔者引入图论(graph theory)对网络做定量分析。在图论中,一个网络可以表述为一个图。图由2个集合构成:节点(vertex或node)集合和边(edge或link)集合。节点集合的大小表示网络的规模,边集合的大小表示网络边的总数。权值(weight),即为边赋权值,表示节点i、j之间关系的强弱。度(degree)是对节点互相连接统计特性最重要的描述。节点i的度定义为网络与该节点直接相连的其他所有节点的数量。节点的度越大,该节点的连接就越多,意味着该节点某种意义上在网络中的地位越重要[8]。
2 结果
2.1θ波段相干分析
在171对导联中随机选取30对作为样本,绘制f-C关系曲线;记忆字母串和记忆图片状态分别与静息状态对比,观察两导信号在δ (2~4 Hz)、θ(4~7 Hz)、α1(7~10 Hz)、α2(10~13 Hz)、β(13~19 Hz)和γ(19~32 Hz) 各波段的相干性。结果表明,WM状态,信号对在θ(4~7 Hz)频段同步效应出现明显增强的现象。如图2所示:(a)4~7 Hz频段,受试者记忆字母串时,F3-T5信号的相干系数明显高于静息态;(b)4~7 Hz频段,受试者记忆图片时,C3-T5信号的相干系数明显高于静息态。

图2 f-C关系曲线。(a)F3-T5信号对在静息态(虚线)和记忆字母串状态(实线)下的f-C关系曲线;(b)C3-T5信号对在静息态(虚线)和记忆图片状态(实线)下的f-C关系曲线Fig.2 f-C curves.(a):f-C curves of F3-T5 during resting (the dotted line) and retention of characters strings (the solid line) intervals; (b):f-C curves of C3-T5 during resting (the dotted line) and retention of drawings (the solid line) intervals

图3 基于θ (4~7 Hz)频段相干的工作记忆功能网络。(a)语音回路;(b)视空间模板;(c)视空间模板比语音回路增多的连接Fig.3 WM functional networks based on the θ (4~7 Hz) band coherence.(a) Phonological loop; (b) Visual-spatial sketch pad; (c) Increased connections of visual-spatial sketch pad compared with phonological loop
2.2WM功能网络
经计算,选取T=0.035,构建基于θ(4~7 Hz)波段相干的ΔBve和ΔBvi,绘制θ波段的WM功能网络,如图3所示。图3(a)表示与静息状态相比,记忆字母串时,θ相干明显增强的连接,表明后颞叶(T5)、中央(Cz、C3、C4)、额叶(F3、F4)、顶叶(Pz、P4)和枕叶(O1、O2)之间的同步效应明显增强;图3(b)表示与静息状态相比,记忆图片时θ相干明显增强的连接;说明人类在记忆图片时,颞叶(T3、T4)、后颞叶(T5、T6)、中央(C3、C4、Cz)、额叶(F3、F4、Fz)、顶叶(Pz、P3、P4)和枕叶(O1、O2)这些脑区之间的同步效应明显增强。记忆图片与记忆字母串对比,二者的头皮连接模式存在很大差异。图3(c)是记忆图片比记忆字母串增加的连接,可以看到记忆图片参与进来的脑区范围更广,连接更多;并且字母串记忆时连接主要出现在大脑左半球,而记忆图片时T6的连接增多,使得连接模式基本左右对称。
统计语音回路和视空间模板网络中各连接的权值如图4所示,横坐标为WM功能网络中的连接,纵坐标表示连接的权值,即WM态连接的相干系数与静息态的差值,差异显著性检验采用方差分析(analysis of variance,ANOVA)。例如,图4(a)中,F3-T5表示图3(a)中的连接F3-T5;图4(a)中F3-T5的值为0.039,表示图3(a)中F3-T5连接的权值为0.039,即与静息态相比,记忆字母串时F3-T5的脑电图 θ频段相干系数增加了0.039,且P<0.01,具有显著差异。图4表明,WM功能网络中各连接的相干系数较静息态有显著提高,且均满足差异显著性检验。
为使结果更加直观,统计图3(a)和(b)中每个节点的度,即每个电极的连接数,结果如表1所示。表1表明,语音回路功能网络中,T5与其它电极共建立7个连接,T6只1个连接;视空间模板网络中,T5的连接数为14个,T6的连接数增加到6个。与其它电极相比,T5,T6的连接数较多,且在WM两个子系统中差异较大,可见T5,T6在WM功能网络中占据重要地位。

表1 语音回路和视空间模板功能网络中各节点的度Tab.1 Degrees of each node in the functional networks of phonological loop and visual-spatial sketch pad.

图4 工作记忆功能网络中各连接的权值。(a)语音回路(P值表示记忆字母串态与静息态差异的显著水平);(b)视空间模板(P值表示记忆图片态与静息态差异的显著水平)Fig.4 Weights of the connections in the WM functional networks.(a) Phonological loop (The value of P indicates significant difference level between retention of characters strings and resting intervals); (b) Visual-spatial sketch pad (The value of P indicates significant difference level between retention of drawings and resting intervals)
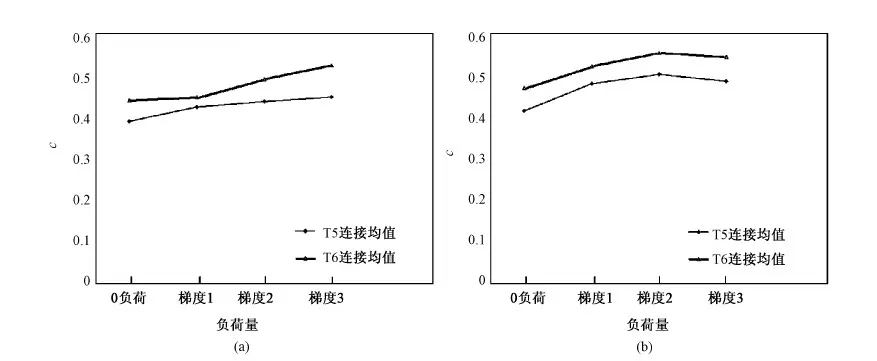
图5 相干值与记忆负荷折线。(a)语音回路;(b)视空间模板Fig.5 Line chart of the relationships between Coherence C and memory loads.(a) Phonological loop; (b) Visual-spatial sketch pad
2.3后颞叶连接与记忆负荷
上述结果表明了后颞叶于WM系统的重要性,因而有必要将结果按记忆的负荷量来分组,观察后颞叶连接的相干强度与记忆负荷之间的关系。以负荷量为自变量、相干值为变量绘制曲线(见图5),这里的相干值指的是后颞叶所有连接的平均相干值。在记忆字母串任务中,T5、T6连接均值都随着记忆负荷的增加而增大,见图5(a)。在记忆图片时,后颞叶连接均值有明显的递增趋势,但当记忆负荷达到一定阈值时,相干值不再增加,见图5(b)。
3 讨论
从图3可以看到,WM中后颞叶、额叶、中央、顶叶和枕叶之间的θ频段EEG相干增强,尤其是后颞叶与其他脑区的连接尤为显著。由图3(a)和表1看出,语音回路的功能网络出现了左侧优势现象,即左后侧颞叶相比右后侧颞叶在字母串记忆任务中贡献更为突出;图3(b)的左侧优势现象减弱;由图3(c)和表1来看,右后颞叶活跃地参与了图片记忆,缓解了偏侧化现象。结果还显示,WM中额叶只与大脑后半球的左后侧颞叶建立了唯一的连接。研究表明,θ同步支持工作记忆和长时记忆,而后颞叶在基于θ波段相干的功能网络中占据重要地位[5-6]。可见,后颞叶是WM的关键区域。
3.1语音回路的专家化区域
本实验观测到的结果与最近对人类和大鼠的广泛实验与研究结果相吻合,即颞叶对WM具有重要意义。例如,朱丹彤等的研究表明,左侧颞叶切除的病人,其言语记忆、命名和语言概念能力重度受损;而右颞叶癫痫病人较左颞叶癫痫病人,言语记忆要好[12]。又如,Zhao等采用内容不相关任务(content-irrelevant task)设计实验,刺激物为汉字、人脸、汉字笔画组合和线条画,记录左右脑(P7,P8,PO7,PO8,PO5,PO6,PO3,PO4,CB1,CB2)的N170[13],结果发现汉字N170振幅的左半球高于右半球,存在左侧优势的现象。上述研究中的言语记忆、命名等能力,识别、记忆汉字和本实验中的记忆字母串,都主要调用WM的子系统——语音回路(识别汉字和记忆字母串可以采用默读复述,将符号转化为语音)。解剖学研究证实,切除左侧颞叶影响病人的语言能力;ERP研究表明,在与语音有关的认知活动中,N170的振幅左颞叶高于右颞叶;而本实验研究得到的语音回路功能网络,出现了明显的后颞叶左侧优势现象。因此可以推断,左后侧颞叶与语音认知有关,进而说明在WM中左后侧颞叶是负责语音回路的专家化区域。
3.2视空间模板的专家化区域
研究表明,视空间模板与右侧大脑半球的结构和功能存在偏相关[14-15]。朱丹彤等的研究表明,大鼠空间记忆任务的完成对海马区是否受损敏感,而人类则对近中颞叶结构是否损害敏感[12]。视觉空间记忆与右海马有关,而言语记忆与左海马有关。右颞叶病变的病人对不易用语言描述的物体,如面容、抽象的图案、旋律和空间定位的记忆能力受到损害。由图3(c)和表1来看,视空间模板与语音回路网络最明显的差异是右后颞叶(T6)与其他脑区的连接明显增加,使得视空间模板网络左右基本对称。可以推测,右后颞叶主要负责视空间模板,相对左后颞叶负责语音回路。以Baddeley工作记忆模型[1]为依据设计的两组实验,得到的结果表明语音和视觉WM有各自的专家化区域,反过来又证实了该模型的正确性,语音和视觉记忆是有不同的生理通路的。
3.3负责工作记忆注意控制机制的区域
虽然说左后颞叶主要负责与语音有关的加工,但在实验2中大脑加工图片信息时,左后侧颞叶的连接没有减少反而增加。两组实验结果的相似之处是:无论是语音回路还是视空间模板,左后侧颞叶与中央、顶叶、枕叶都有很广泛的联系,尤其是额叶与大脑后半球建立的唯一连接也是通过左后侧颞叶。另外,Prida等人用EEG相干研究视觉注意机制,发现“注意”状态与“转移”状态相比,左颞叶和顶枕区出现了类似很明显的相干性增强,而右颞叶则没有这种现象,体现了左颞叶对视觉注意调制的重要性[16]。因此推测,左后侧颞叶除了起到加工语音信息的作用之外,还有某种最基本的功能,它对各种认知活动都起到调制作用,具体来说可能是协同连接步调的作用。研究表明,海马体包含θ步调发生器(θ-pacemakers),并且在解剖学和功能上它与大脑皮质所有区域都有很广泛的联系[17-18],再者锁相海马皮质回路(phase-locked hippocampo-cortical loops)与记忆的编码和检索记忆都有关[17-18]。在WM和注意等其他认知活动中,很有可能是左海马协调各脑区的θ振荡同步,从而促进各区域间的沟通。通过上述实验观察结果,结合相关文献研究报道分析,启示我们做以下推论:左后侧颞叶很有可能是负责WM模型第3个部分——中央执行系统(central executive)的主要区域。Sarnthein等认为,前额是负责WM注意和控制的主要区域[5],然而本研究并没有发现前额参与到WM中来,并且Prida等在对注意机制进行研究时,也没有发现前额的相应活动,连接最为广泛的仍然是左颞叶[16]。当然,这样的结果可能是由于本研究数据处理用的时间窗比较粗糙(4 s),而额叶皮质的注意调制被描述为是一个快速短暂的过程[19-20]。
3.4相干强度与记忆负荷
如图5所示,后颞叶连接的相干值随着记忆负荷的增加而增大,这一结果与笔者的预期相一致,证明了用EEG相干来研究WM神经机制的合理性。随着记忆负荷的增强,大脑处理的信息量增大,对于WM系统尤为重要的后颞叶与其他脑区之间的同步效应随之增强;当记忆负荷达到脑容量的最大值时,同步效应达到最强;再增加记忆负荷,同步效应不增反减。 在实验中观察到:受试者在记忆较少的字母串或较简单的图形时,表现得较为轻松;随着记忆负荷量的加大,受试者投入更多的注意力和精力;当负荷量过大时,受试者往往表现出消极记忆的态度,正确率明显下降。EEG相干结果与行为结果一致。
3.5方法局限性与改进
本实验所观测到的语音回路偏侧化现象与Zhao等的研究所得到的结果吻合,即汉字N170的左侧优势现象与受试者的识字水平正相关,阅读能力越强,左偏侧化现象越明显[21]。但是,语音回路的偏侧化现象,究竟是人类大脑与生俱来的构造决定的,还是人类在从小到大习得语言的过程中大脑与学习相互作用的结果,依然还有疑问,还需要对不同年龄段和受教育水平的受试进行更为广泛和系统的研究。负责中央执行系统的主要区域是前额、颞叶,还是其他脑区,需要今后用更小的时间窗来研究WM的动态功能网络。除此之外,对于式(2)中阈值T的选取未能提供一个明确准则,仅是参照网络密度和差异性检验,经过多次计算、比较得到的数值。阈值的选取对网络的拓扑结构有很大影响,也是构建网络的一个难点,这需要在下一步研究中深入探讨。采用无向网络分析WM的机制,忽略了神经活动之间的因果性,从而探讨WM大脑实时的功能活动机制,也是今后的研究方向之一。
4 结论
通过构建基于EEG相干同步的功能网络,研究大脑后颞叶在不同模式WM过程中的作用和职能,结果表明:后颞叶与额叶、中央、顶叶和枕叶之间的θ频段(4~7Hz)相干同步效应调节了WM过程,其中右后侧颞叶负责视空间模板,左后侧颞叶与语音回路有关。结合相关文献研究报道进行分析,得出推论:左后侧颞叶很有可能是负责WM中央执行系统的主要区域,有可能起到了协同各脑区θ连接步调、负责WM中的注意控制的作用。
致谢(感谢徐鑫和万小姣在搭建实验平台和编写数据处理平台方面给予的帮助)
[1] Baddeley AD,Hitch GJ.Working memory [M]// Bower GH,eds.The Psychology of Learning and Motivation.New York:Academic Press,1974,8:47-89.
[2] 丁锦红,郭春彦.工作记忆的脑机制研究[J].心理科学,2001,24(5):583-585.
[3] 功能性磁共振成像[DB/OL].http://zh.wikipedia.org/wiki/FMRI,2014-05-16/2014-07-19.
[4] Guo Chunyan,Lawson AL,Jiang Y.Distinct Neural Mechanisms for Repetition Effects of Visual Objects[J].Neuroscience,2007,149:747-759.
[5] Sarnthein J,Petsche H,Rappelsberger P,etal.Synchronization between prefrontal and posterior association cortex during human working memory[J].Proc Natl Acad Sci USA,1998,95:7092-7096.
[6] Fell J,Axmacher N.The role of phase synchronization in memory processes[J].Nature Reviews,Neuroscience,2011,12:105-118.
[7] Yan Zheng,Gao Xiaorong.Functional connectivity analysis of steady-state visual evoked potentials[J].Neuroscience Letters,2011,499:199-203.
[8] 梁夏,王金辉,贺永.人脑连接组研究:脑结构网络和脑功能网络[J].科学通报,2010,55(16):1565-1583.
[9] Tass P,Weule J,Kurths J,etal.Detection of n:m phase locking from noisy data:application to magnetoencephalography[J].Phys Rev Lett,1998,81:3291-3294.
[10] Lachaux JP,Rodriguez E,Martinerie J,etal.Measuring phase synchrony in brain signals[J].Hum Brain Mapp,1999,8:194-208.
[11] 赵丽娜,王保强,尧德中.基于信号处理的脑电相位同步性分析方法研究[J].生物医学工程学杂志,2008,25(2):250-254.
[12] 朱丹彤,杨晓苏,金丽娟.颞叶癫痫与认知功能[J].现代康复,2001,5(12):54-55.
[13] Zhao Jing,Li Su,Lin Sien,etal.The selectivity of N170 in the left hemisphere as an electrophysiological marker for expertise in reading Chinese[J].Neurosci Bull,2012,28(5):577-584.
[14] Migliaccio R,Agosta F,Toba MN,etal.Brain networks in posterior cortical atrophy:a single case tractography study and literature review[J].Cortex,2012,48(10):1298-1309.
[15] Stretton J,Winston G,Sidhu M,etal.Neural correlates of working memory in temporal lobe epilepsy-an fMRI study[J].Neuro Image,2012,60(3):1696-1703.
[16] Prida LM,Barceló F,Pozo MA,etal.Local pattern of synchronization in extraestriate networks during visual attention[C]//IWANN '99,Engineering Applications of Bio-Inspired Artificial Neural Networks.Alicante:LNCS,1999,1607:273-278.
[17] Buzsaki G.Cereb[J].Cortex,1996,6:81-92.
[18] Miller R.Cortico-Hippocampal Interplay and the Representation of Contexts in the Brain[M].Berlin:Springer-Verlag Berlin and Heidelberg GmbH & Co K,1991.
[19] Barceló F,Rubia FJ.Non-frontal P3b-like activity evoked by the Wisconsin Card Sorting Test[J].Neuroreport,1998,9:747-751.
[20] Mclntosh AR,Grady CL,Ungerleider LG,etal.Network analysis of cortical visual pathways mapped with PET[J].J Neurosci,1994,14:655-666.
[21] Li Su,Lee Kang,Zhao Jing,etal.Neural competition as a development process:Early hemispheric specialization for word processing delays specialization for face processing[J].Neuropsychologia,2013,51:950-959.
SynchronizationbetweenPosteriorTemporalLobeandotherBrainRegionsintheWorkingMemory
Mei Dandan Yan Zheng Huang Ruimin*
(CollegeofInformationScienceandEngineering,HuaqiaoUniversity,Xiamen361021,Fujian,China)
In this paper,the coherence method of electroencephalogram (EEG) was employed to establish functional networks to study the main areas which are responsible for the working memory (WM) —— the role and function of posterior temporal lobes presented in the processes of different WM modes.Twelve subjects performed our experiments which required the retention of either verbalizable strings of letters or abstract drawings.These two paradigms were designed specifically for corresponding to the two components of working memory —phonological loop and visual-spatial sketch pad respectively.A 19-channels EEG signal was recorded,based on which the coherent coefficient matrices were calculated.In consequence,the brain functional networks were constructed and discussed in this paper.Results showed that,compared with the resting state,for both phonological loop and visual-spatial sketch pad,significantly increased θ (4~7 Hz) coherence values were detected between posterior temporal lobes (T5,T6) and other brain regions (P<0.05).This result suggested between the posterior temporal lobes and other brain regions the θ synchronization phenomenon could be observed during the process of working memory.Furthermore,the degrees of nodes T5 and T6 were largely different in the functional networks of the two components.However,both the degrees of T5 and T6 were bigger than other nodes,suggesting the left and right posterior temporal lobes may serve as specialized regions for the phonological loop and visual-spatial sketch pad respectively.
working memory; posterior temporal lobes; coherent synchronization; functional network
10.3969/j.issn.0258-8021.2015.02.007
2014-08-18,录用日期:2015-01-06
国家自然科学基金(61203369);中央高校基本科研业务费专项基金(12Y0308)
R318
A
0258-8021(2015) 02-0176-08
*通信作者(Corresponding author),E-mail:hrm@hqu.edu.cn
猜你喜欢
浙江大学学报(理学版)(2021年1期)2021-01-26
科学(2020年2期)2020-01-08
家庭科学·新健康(2019年9期)2019-10-21
中老年保健(2019年7期)2019-01-13
中学科技(2018年9期)2018-12-19
健康管理(2017年3期)2017-04-20
中国生物医学工程学报(2017年6期)2017-02-10
中西医结合心脑血管病杂志(2016年20期)2016-03-01
医学研究杂志(2015年9期)2015-07-01
中国医学科学院学报(2013年3期)2013-03-11
