
NTERA-2的精原干细胞生物学特征鉴定*
2015-08-24 01:22:29王姿力周勇华莫艳秀肖亚梅
激光生物学报 2015年4期
王姿力,周勇华,罗 超,莫艳秀,2,肖亚梅*
(1.湖南师范大学生命科学学院教育部蛋白质化学和鱼类发育生物学重点实验室,湖南长沙410081;2.湘南学院基础医学部,湖南郴州423000)
NTERA-2的精原干细胞生物学特征鉴定*
王姿力1,周勇华1,罗 超1,莫艳秀1,2,肖亚梅1*
(1.湖南师范大学生命科学学院教育部蛋白质化学和鱼类发育生物学重点实验室,湖南长沙410081;2.湘南学院基础医学部,湖南郴州423000)
本研究通过免疫荧光分析,鉴定NTERA-2细胞是否具有精原干细胞特征。研究结果显示,重要精原干细胞标记分子GFRα-1和DDX4在NTERA-2细胞中均表现为阳性表达,NTERA-2细胞具有精原干细胞的特征。初步探讨了重要细胞信号转换蛋白JNK特异性抑制剂SP600125对NTERA-2细胞生长和增殖的影响,为进一步以NTERA-2细胞为精原干细胞模型开展JNK对生殖细胞发育调节功能的研究奠定基础。
NTERA-2;精原干细胞;SP600125;免疫荧光技术
doi:10.3969/j.issn.1007-7146.2015.04.011
睾丸肿瘤是泌尿外科中常见的肿瘤之一,恶性比例高达95%,包括生殖细胞肿瘤、非生殖细胞肿瘤和睾丸继发性肿瘤[1]。睾丸生殖细胞肿瘤(Testicular germ cell tumors,TGCT)约占睾丸肿瘤病例的90%~95%,是睾丸肿瘤中最多见的类型,涉及到几类不同发病机制、病理类型和临床表现,一般又可分为精原细胞瘤、非精原细胞瘤(包括胚胎癌、畸胎瘤、绒毛膜上皮癌等)及混合性生殖细胞瘤[2,3]。Skakkebaek等提出生殖细胞肿瘤是发展于曲精细管内的生殖细胞肿瘤,即睾丸原位癌(Carcinoma in situ,CIS)[4,5]。
NTERA-2(NT2)细胞(Testicular embryonal carcinoma cells)是来源于人睾丸畸胎瘤细胞[6,7]。目前NTERA-2细胞在癌细胞增殖周期分子调控机制、肿瘤细胞凋亡、肿瘤的药物治疗等方面有不少研究报道[8]。同时NTERA-2在细胞扩增和分化方面具有与胚胎干细胞部分相似的分子和细胞生物学特征,维持着胚胎干细胞发育的多能性[9],研究认为NTERA-2细胞可为细胞命运定型以及胚胎干细胞分化过程中的调控机制研究提供细胞研究模型[10,11]。
本研究通过对生殖干细胞表面标志分子的鉴定,确定NTERA-2细胞是否具有生殖干细胞生物学特性,初步探讨了重要细胞信号转换蛋白JNK特异性抑制剂SP600125对NTERA-2细胞生长和增殖的影响,为进一步以NTERA-2细胞为精原干细胞模型开展JNK对生殖细胞发育调节功能的研究奠定基础。
1 实验材料和方法
1.1材料
NT2细胞由本实验室保存。Hoechst 33342购于美国Sigma公司,c-kit,DDX4,GFRα-1抗体均购于Santa Cruz Biotechnology公司,JNK抑制剂SP600125购于碧云天公司,二抗是带荧光标记的羊抗鼠GAM和羊抗兔GAR均购于康为世纪公司,MEM培养基,胎牛血清FBS,双抗青霉素和链霉素均购于Life Technologies公司,其他常规试剂均购自上海生工生物工程有限公司。
1.2细胞培养
NTERA-2细胞用Eagle's Minimum Essential Medium(MEM)培养基培养,培养基中包含10%的胎牛血清(FBS,Life Technologies,Inc.,Grand Island,NY,USA)、1%的抗生素(100 U/mL青霉素和100 ng/mL链霉素;Life Technologies)。在37℃的培养箱中培养。细胞传代时,NTERA-2细胞均经0.25%胰酶(Trypsin-EDTA)溶液消化2-3 min。
1.3细胞免疫荧光染色
细胞接种于四孔板中,接种细胞量约2.5×105cell/孔。待四孔板中的细胞贴壁且生长状态良好后,移去完全培养基,PBS清洗后加入4%多聚甲醛。室温固定30 min,PBS漂洗三次,每次5 min。用现配好的封闭液(含0.1%Triton和2%BSA的PBS缓冲液)封闭处理20 min。滴加一抗(1∶100稀释),4℃孵育过夜。PBS漂洗三次后,加二抗(1∶100稀释),避光室温孵育1 h。PBS漂洗后,加入Hoechst 33342染色液室温避光染15 min,PBS漂洗两次。荧光显微镜下观察。
1.4SP600125抑制处理
将生长处于对数期的NTERA-2细胞接种于6孔板中,细胞接种量约为1.2×106cell/孔。培养24 h后,加入不同浓度的SP600125抑制剂(0,25,50 μmol/L)分别处理24 h,48 h和72 h,置于相差显微镜下观察其生长状态。
2 结果
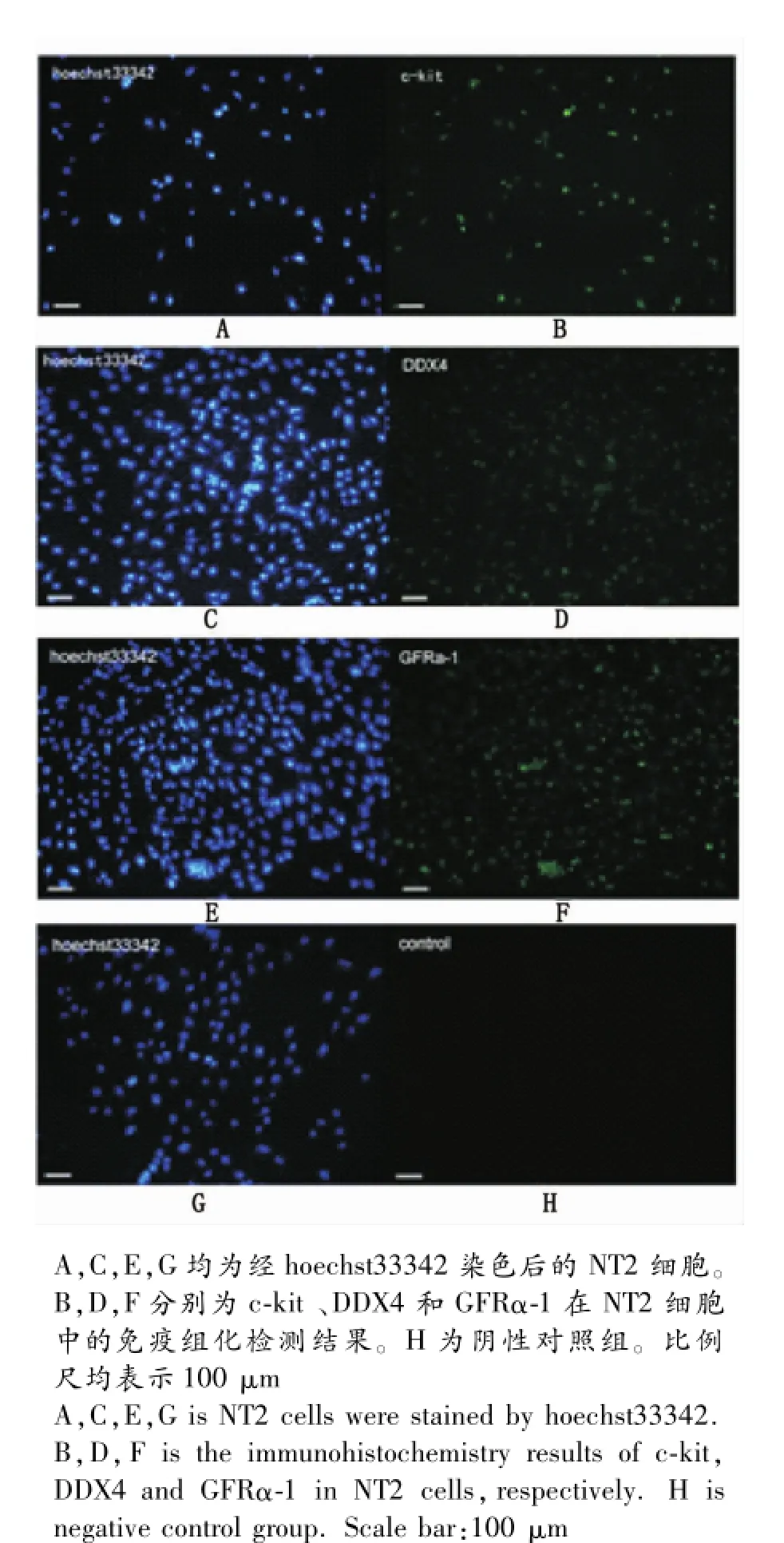
图1 几种重要干细胞因子在NT2细胞的表达鉴定Fig.1 The expression of several important stem cell factor in NT2 cells
2.1几种重要干细胞因子在NTERA-2细胞中的鉴定通过免疫组织化学方法,分析了胚胎干细胞标记分子c-kit和哺乳动物两种重要精原干细胞标记分子GFRα-1和DDX4在NTERA-2细胞中的表达。如图1所示,c-kit、GFRα-1和DDX4在NTERA-2细胞中均呈现出明显的阳性表达。
2.2SP600125抑制剂处理对NTERA-2细胞生长和增殖的影响
用JNK特异性抑制剂SP600125对NTERA-2进行药物孵育实验,观察显示SP600125对NTERA-2的生长具有明显的抑制作用,并导致NTERA-2细胞出现大量凋亡现象(图2)。NTERA-2细胞的凋亡程度与SP600125药物浓度和处理时间呈正相关。
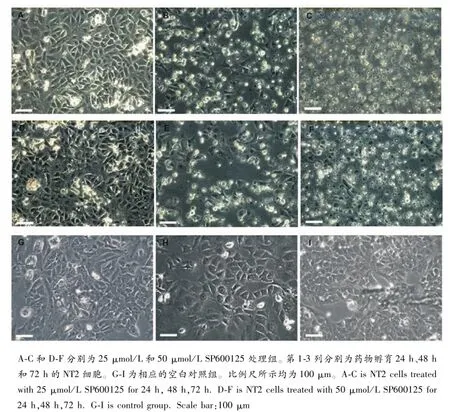
图2 SP600125药物孵育对NT2细胞生长的影响Fig.2 The effect of SP600125 on the NT2 cells
3 讨论
c-kit基因编码一种胚胎时期干细胞生长因子的受体,是生殖细胞和造血干细胞增殖、分化、移行及存活所必需的[12]。DDX4(DEAD box polypeptide 4)基因是DEAD-box基因家族的重要成员,仅在生殖细胞系中特异性表达,在哺乳动物睾丸组织中,DDX4基因从精子发生减数分裂前开始表达一直持续到减数分裂后生精细胞的形成[13]。目前DDX4基因广泛用于配子发生和原生殖细胞的起源、迁移、分化等的研究中。GFRα-1是GDNF(胶质细胞源性神经营养因子)的受体,是啮齿类动物精原干细胞(SSCs)特异的细胞表面分子标志物[14]。研究结果显示,在NTERA-2细胞中不仅有c-kit基因的表达,而且也检测到DDX4和GFRα-1这些精原干细胞特异标记分子的表达,表征着NTERA-2细胞具有精原干细胞特征。
JNK(c-Jun氨基末端激酶)是促分裂原活化蛋白激酶超家族中的主要成员之一。生长因子、细胞因子、应激等多种细胞外刺激通过激活JNK信号通路,在动物的生长发育、细胞凋亡、癌症的发生中发挥着重要作用[15,16]。最新的研究报道了JNK参与维系干细胞发育潜能,在诱导多能性干细胞建立上具有重要调控作用[17]。本实验室也已证实JNK参与动物生殖腺早期分化和发育调控[18]。SP600125是一种常用的c-Jun N-terminal kinase(JNK)的高选择性抑制剂,SP600125处理斑马鱼胚胎或鱼苗,能显著影响斑马鱼性腺的分化发育,证实JNK在鱼类性腺早期分化发育中重要调节功能[19]。但JNK通路在生殖细胞发育调控中的分子机制尚不明晰。本文初次以NTERA-2作为生殖干细胞模型,用JNK抑制剂SP600125处理NTERA-2细胞,研究表明,SP600125能显著抑制NTERA-2细胞的生长和增殖,为进一步深入探讨JNK在精原干细胞增殖与凋亡中的功能奠定了基础。
[1] ULBRIGHT T M.Germ cell neoplasms of the testis[J].The A-merican Journal of Surgical Pathology,1993,17(11),1075-1091.
[2]PIETENPOL J A,BOHLANDER S K,SATO Y,et al.Assignment of the human P27 gene to 12p and its analysis in leukemias[J].Cancer Res,1995,55(6):1206-1210.
[3]HORWICH A,SHIPLEY J,HUDDART R.Testicular germ-cell cancer[J].Lancet,2006,367(9512):754-765.
[4]SKAKKEBAEK N E.Possible carcinoma-in-situ of the testis[J].Lancet,1972,300(7776):516-517.
[5]ALMSTRUP K,HOEI HANSEN C E,NIELSEN J E,et al.Genome-wide gene expression profiling of testicular carcinoma in situ progression into overt tumours[J].Br J Cancer,2005,92(10):1934-1941.
[6]LOOIJENGA L,OOSTERHUIS J W.Pathogenesis of testicular germ celltumours[J].Reviews of Reproduction,1999,4(2): 90-100.
[7]LOOIJENGA L,OOSTERHUIS J W.Pathogenesis of testicular germ celltumours[J].Reviews of Reproduction,1999,4(2): 90-100.
[8]易朵,李晓峰,周畅,等.阿司匹林诱导人恶性睾丸生殖细胞瘤NTera-2细胞凋亡[J].中国生物化学与分子生物学报,2012,28(7):630-636. YI Duo,LI Xiaofeng ZHOU Chang,et al.Curcumin inhibits AP-2γ to induce apoptosis in human malignant testicular germ cell line NTera-2[J].Chinese Journal of Biochemistry and Molecular Biology,2012,28(7):630-636.
[9]LOOIJENGA L,OOSTERHUIS J W.Pathogenesis of testicular germ celltumours[J].Reviews of Reproduction,1999,4(2): 90-100.
[10] RAJARSHI PAL,GEETA RAVINDRAN.Assessment of pluripotency and multilineage differentiation potential of NTERA-2 cells as a model for studying human embryonic stem cells[J]. Cell Prolif,2006,39(6):585-598.
[11]PRZYBORSKI S A,CHRISTIE V B,et al.Human embryonal carcinoma stem cells:models of embryonic development in humans[J].Stem Cells Dev,2004,13(4):400-408.
[12]RONNSTRAND L.Signal transduction via the stem cell factor receptor/c-Kit[J].Cellular and Molecular Life Sciences,2004,61(19-20):2535-2548.
[13]HICKFORD D E,FRANKENBERG S,DASK A J.et al. DDX4(VASA)is conserved in germ cell development in marsupials and monotremes[J].Biology of Reproduction,2011,85(4):733-743.
[14]TADOKORO Y,YOMOGIDA K,OHTA H,et al.Homeostatic regulation of germinal stem cell proliferation by the GDNF/FSH pathway[J].Mechanisms of Development,2002,113(1): 29-39.
[15]VAN HORN R D,CHU S,FAN L,et al.Cdk1 activity is required for mitotic activation of aurora A during G2/M transition of human cells[J].2010,285(28):21849-21857.
[16]WESTON C R,DAVIS R J.The JNK signal transduction pathway[J].Curr Opin Cell Biol,2007,19(2):142-149.
[17]SCHWARTZ C M,SPIVAK C E,BAKER S C et al.NTera2: a model system to study dopaminergic differentiation of human embryonic stem cells[J].Stem Cells Dev,2005,14(5): 517-534.
[18]KATNER A L,GOOTMA P,HOANG Q B,et al.A recombinant adenoviurs expressing P7(kiP1)induces cell cycle arrest and apoptosis in human 786-0 renal carcinoma cell[J].J Urol,2002,168(2):766-773.
[19]YAMEI XIAO,YONG HUA,XIONG ZHEN,et al.Involvement of JNK in the embryonic development and organogenesis in Zebrafish[J].Mar Biotechnol(NY),2013,15(6): 716-725.
NTERA-2 cells with Characteristics of Spermatogonial Stem Cells
WANG Zili1,ZHOU Yonghua1,LUO Chao1,MO Yanxiu1,2,XIAO Yamei1*
(1.Key Lab of Protein Chemistry and Developmental Biology of Education Ministry of China,College of Life Sciences,Hunan Normal University,Changsha 410081,Hunan,China;2.Department of Bascal Medicine,Xiang Nan University,Chenzhou 423000,Hunan,China)
This study was to identify the characteristics of NTERA-2 cells to find whether out has it the characteristics of spermatogonial biology.Our results showed that NTERA-2 cells expressed some markers of spermatogonial stem cells,such as GFRα-1 and DDX4.It suggest that NTERA-2 cells maybe useful as a model for studying spermatogonial stem cell proliferation and differentiation in vitro.With the treatment by SP600125,the proliferation of NTERA-2 cells is seriously inhibited.The results are the foundation of further research on the function of JNK in germ cell development.
NTERA-2;spermatogonial stem cell;SP600125;immunofluorescence
Q492
A
1007-7146(2015)04-0364-04
2014-12-15;
2015-04-14
国家自然科学基金(31172399)资助项目;湖南省教育厅项目资助(15C1274);湖南省研究生科研创新项目资助(CX2013B212)
王姿力(1989-),女,汉族,湖南津市人,硕士研究生,主要从事动物发育中的信号传导机制研究。(电话)15874113790;(电子邮件)camilla.wzl@foxmail.com
肖亚梅(1968-),女,湖南宜章人,湖南师范大学生命科学学院教授,主要从事动物发育和人类疾病中的信号传导机制研究。(电话)0731-88872297;(电子邮箱)yameix@126.com
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26 07:11:58
中国生殖健康(2020年5期)2021-01-18 02:59:48
国际放射医学核医学杂志(2020年4期)2020-07-27 01:53:28
中国生殖健康(2019年7期)2019-01-06 09:27:30
中国生殖健康(2018年5期)2018-11-06 07:15:38
精准医学杂志(2018年5期)2018-02-12 19:03:57
临床与实验病理学杂志(2017年6期)2017-03-07 16:52:29
新疆医科大学学报(2015年10期)2015-12-26 12:33:30
中国卫生(2014年11期)2014-11-12 13:11:24
安徽医药(2014年9期)2014-03-20 13:14:09
