
枯草芽孢杆菌21抑菌物质的稳定性分析及初步分离
2015-08-06 03:31:06郭春兰于春生张雨竹宋志儒杨月肖亚静孙冬梅
黑龙江八一农垦大学学报 2015年1期
郭春兰,于春生,张雨竹,宋志儒,杨月,肖亚静,孙冬梅
(黑龙江八一农垦大学生命科学技术学院,大庆 163319)
枯草芽孢杆菌(B.subtilis)作为植物病害主要生防细菌之一,备受各国研究工作者的青睐[1]。B.subtilis在生长代谢过程中能够产生不同种类的拮抗物质,主要为抗生素和抗菌蛋白,大多数能忍受蛋白酶的作用,但也有一些对蛋白酶敏感[2-3];且多数能拮抗细菌,少数拮抗真菌[4-5]。国内对枯草芽孢杆菌及其代谢产物稳定性分析研究也很多,枯草芽胞杆菌B-916中分离出来的对真菌具有较强抑菌活性的蛋白,大小为41.9 kDa,具有一定耐热性,对蛋白酶敏感,具有较宽的pH 适应范围[6];具有抑制黄瓜枯萎病原菌作用的枯草芽胞杆菌中纯化出具有抑菌活性的蛋白L37,其分子量为15 kDa,该蛋白对多种病原真菌具有抑制效果,且对蛋白酶、温度等均不敏感,是一种非常稳定的抑菌蛋白[7]。拮抗棉花黄萎菌的枯草芽孢杆菌BDT- 2 能产生分子量为约为30 kD 的抑菌物质,有较好的酸碱稳定性[8]。通过对枯草芽孢杆菌21发酵液抑菌物质的稳定性进行分析(包括蛋白酶稳定性、温度稳定性、酸碱稳定性),并将生的抑菌物质进行了初步分离,为该菌产生的抑菌物质成分进一步分离、纯化和结构测定打下了基础。
1 材料及方法
1.1 材料
1.1.1 供试菌种
拮抗枯草芽孢杆菌21、玉米圆斑病菌dq-1 由黑龙江八一农垦大学生命科学技术学院生物技术系微生物实验室分离并保存。
1.1.2 供试培养基
马铃薯培养基(PDA):去皮马铃薯200 g,蔗糖20 g,琼脂15 g,蒸馏水1000 mL,pH 自然。
牛肉膏蛋白胨(培养基):牛肉膏3 g,蛋白胨10 g,氯化钠5 g,琼脂15 g,蒸馏水1 000 mL,pH 自然。
1.2 实验方法
1.2.1 枯草芽孢杆菌21 上清液的准备
将活化的枯草芽孢杆菌21 接种到无菌的牛肉膏蛋白胨液体培养基中,180 r·min-1、28 ℃摇床振荡培养24 h,作为种子液;将种子液8 mL 接种于装液量为100 mL 的牛肉膏蛋白胨培养基中,培养条件同上培养24 h,所得发酵液经4 000 r·min-1离心,取上清备用。
1.2.2 抑菌物质的稳定性分析
1.2.2.1 抑菌物质的温度稳定性分析
将1.2.1 获得的发酵上清液分别经40、50、60、70、80、90、100 ℃等不同温度下处理30 min,以未经处理的发酵上清液为对照,以玉米圆斑病菌为指示菌,采用牛津杯定量扩散法进行抑菌效果研究,用直径0.55 cm 的打孔器将活化的玉米圆斑病菌打成大小均匀的菌碟接到PDA 平板的中央,距离菌碟边缘3 cm 处接上牛津杯,将150 μL 的无菌发酵液接到牛津杯中,3 d 后测量并记录抑菌半径。
1.2.2.2 抑菌物质的酸碱稳定性分析
将1.2.1 获得的发酵上清液分别用1 mol·L-1HCl或 NaOH 调 成 不 同 pH 值,pH 梯 度 分 别 为2.0,3.0,4.0,5.0,6.0,7.0,8.0,9.0,10.0,平衡30 min后,以玉米圆斑病菌作为供试菌进行活性检测,以未经处理的发酵上清液为对照,采用牛津杯定量扩散法进行抑菌效果研究。
1.2.2.3 抑菌物质的蛋白酶稳定性分析
将1.2.1 获得的发酵上清液分别用1N 的NaOH调至pH=7,加入胰蛋白酶、木瓜蛋白酶、蛋白酶K、碱性蛋白酶至终浓度为0.2 mg·mL-1,37 ℃水浴中酶解4 h。以未经处理的发酵上清液作对照,以玉米圆斑病菌为指示菌,采用牛津杯定量扩散法进行抑菌效果研究。相对抑菌能力=处理后的抑菌圈半径/对照组抑菌半径×100%。
1.2.3 抑菌物质的初步分离
1.2.3.1 硫酸铵分级盐析
取枯草芽孢杆菌21 发酵液500 mL,经4 000 r·min-1离心10 min 后取上清液,弃除菌体和发酵残渣,向上清液中加入固体硫酸铵至20%饱和度(冰浴中),冰箱中静置6 h 后,4 000 r·min-1离心15 min,收集沉淀。将收集的上清液在冰浴中再加入固体硫酸铵,依次按照同样的方法调节硫酸铵饱和度分别为30%、40%、50%、60%、70%、80%、90%。收集每一个饱和度的沉淀蛋白,分别溶解在10 mL 50 mmol·L-1pH6.0 的磷酸缓冲液中,转入8 000~14 000 kDa 透析袋中,放在外液为磷酸缓冲液的容器中充分透析脱盐,3~4 h 更换一次外液,更换外液4~5 次,中间用5%氢氧化钡检测除盐情况。透析好的的蛋白粗提液经0.22 μm 孔径的细菌过滤器过滤制成无菌滤液,以玉米圆斑病病菌为指示菌,分别测定各沉淀成分的活性。
2 结果分析
2.1 发酵液的稳定性分析
2.1.1 发酵液的温度稳定性分析
发酵液经40~50 ℃的不同温度处理后,与对照组总比抑菌效果没有明显的改变,相对抑菌能力稳定在88%~100%,与对照差异不显著(P=0.05)。发酵液经80~90 ℃处理后,抑菌效果发生明显变化,相对抑菌能力低于65%,当处理温度为100 ℃时,发酵液几乎没有抑菌效果(图1,2)。

图1 抑菌活性物质的温度稳定性Fig.1 Heat stability of the antibacterial activity
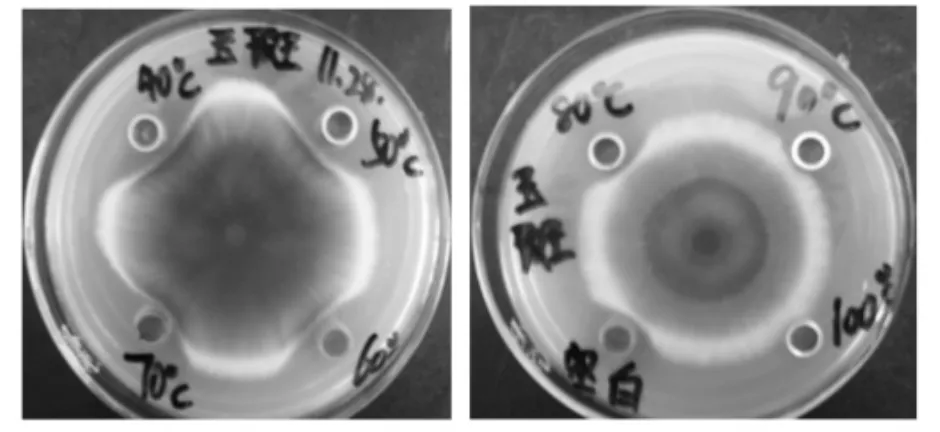
图2 抑菌活性物质的温度稳定性Fig.2 Heat stability of the antibacterial activity
2.1.2 发酵液的酸碱稳定性分析
发酵液经不同的pH 处理后,在pH 5 之间时抑菌活性稳定,与对照组比差异不显著(P=0.05),当pH 值4,6~9 时。抑菌效果发生明显的下降趋势,相对抑菌能力在60%~50%,且与对照组差异显著(P=0.05),当pH值低于4 或高于9 时发酵液几乎没有活性(图3,4)。
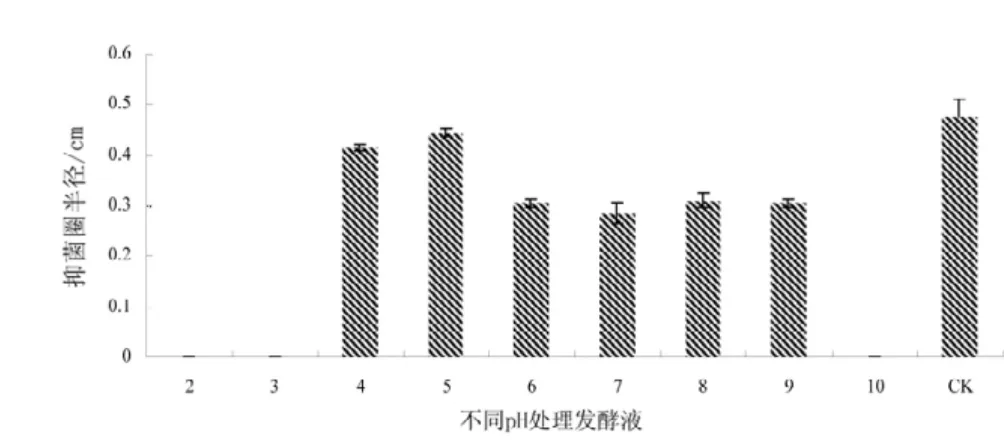
图3 不同pH 处理对发酵液的抑菌活性影响Fig.3 Activity of the antibacterial compounds at different pH values

图4 不同pH 处理对发酵液的抑菌活性影响Fig.4 Activity of the antibacterial compounds at different pH Values
2.1.3 发酵液的蛋白酶稳定性分析
用不同的蛋白酶对发酵液进行处理后,发酵液的抑菌活性在碱性蛋白酶和木瓜蛋白酶的作用下变化较明显,相对抑菌能力下降到70%~80%,与对照组比差异显著(P=0.05)胰蛋白酶和蛋白酶K 对发酵液的抑菌活性影响不大,相对抑菌能力仍在95%以上,与对照组比差异不显著(P=0.05)(图5,6)。

图5 不同蛋白酶处理对发酵液抑菌活性物质的稳定性Fig.5 Sensitivity of the antibacterial compounds to proteolytic enzymes
发酵液的稳定发性分析得到发酵液在温度40~50 ℃、pH5,有较好的稳定性,但对不同蛋白酶稳定性不同,其对碱性蛋白和木瓜蛋白酶较稳定,对胰蛋白酶和蛋白酶k 表现很好的稳定性。
2.2 粗提蛋白的抑菌效果
由图7,8 可知,拮抗物质在硫酸铵饱和度为30%~70%时被沉淀下来,饱和度为60%以下时沉淀的拮抗物质较多,且多次离心等操作会使拮抗物质有所损失或失去活性,所以,选定硫酸铵的沉淀浓度为60%。将所得到的粗提物有经过茚三酮反应,初步确定粗提物为蛋白类。

图7 不同硫酸铵饱和度沉淀的蛋白质的抑菌效果Fig.7 The antibacterial effects of the crude extract proteins after ammonium sulfate precipitation

图8 不同硫酸铵饱和度沉淀的蛋白质的抑菌效果Fig.8 The antibacterial effects of the crude extract proteins after ammonium sulfate precipitation
3 结论与讨论
枯草芽孢杆菌的利用途径有两种:一是将制成活菌制剂使用;二是通过发酵,利用其产生的抗菌活性物质。枯草芽孢杆菌分泌的抗菌物质主要有两类,即低分子量抗菌素和高分子量抗菌蛋白质[9]。
研究通过对枯草芽孢杆菌21 的抑菌物质进行稳定性分析和初步分离可知:抑菌成分对蛋白酶具有一定的耐受性,特别是对蛋白酶K 和胰蛋白酶,对温度和酸碱度比较敏感。除分离的具有耐热性、耐酸碱性的低分子的抗菌物质,也已经分离到的分子量较大的蛋白类抑菌物质,大部分表现出耐热性、对酸碱环境不同的适应性以及对蛋白酶的敏感性[10-12]。硫酸铵沉淀法浓缩和初级纯化蛋白质常用的方法,大多数的研究者都是通过先用硫酸铵沉淀,在经过柱层析或薄层层析,SDS-PAGE 电泳的方法将所得到的蛋白进一步纯化,进行提纯的;孙瑶[13],贾翠英[14],陈爱香[15]等分离的抑菌蛋白都是先由硫酸铵沉淀方法进行粗提的,高浓度的盐离子在蛋白质溶液中可与蛋白质竞争水分子,从而破坏蛋白质表面的水化膜,降低其溶解度,使之从溶液中沉淀出来,各种蛋白质的溶解度不同,因而可利用不同浓度的盐溶液来沉淀不同的蛋白质,这种方法称之为盐析。经不同硫酸铵饱和度沉淀得到了具有抑菌作用的物质为蛋白类的抑菌成分,可能为蛋白类或肽类,是否含有其他的抑菌成分,还需要进一步研究,从发酵液中能得到该物质,表明起作用的应属典型的分泌型蛋白,分泌型蛋白的种类现在主要包括低分子抗菌肽和大分子蛋白类,在今后的实验中经进一步纯化该抑菌蛋白,更好地利用该枯草芽孢杆菌提供理论依据。
[1]杨佐忠.枯草杆菌拮抗体在植物病害生物防治中的应用[J].林业科技,2001,22(3):41-43.
[2]张成省,孔凡玉,关小红,等.烟草根围细菌Tpb55 菌株的鉴定及其抑菌活性[J].中国生物防治,2008,24(1):63-68.
[3]邱德文.微生物蛋白农药研究进展[J].中国生物防治,2004,20(2):91-94.
[4]吴艳,闰豫君,赵思峰,等.组合芽孢杆菌抑菌物质特性及其抑菌效果研究[J].西北农业学报,2007,16(5):266-270.
[5]陈其军,李娟,孙启利,等.水稻内生枯草芽孢杆菌G87抑菌蛋白的分离纯化及理化特性[J].微生物学报,2010,50(10):1353-357.
[6]王艳红,曹宁,于海威,等.拮抗番茄早疫病原菌的放线菌WL07 发酵产物理化性质的研究[J].黑龙江八一农垦大学学报,2008,20(3):1-4.
[7]孙瑶.棉花黄萎病菌拮抗细菌TYG-2 抑菌蛋白的分离纯化[J].山西农业大学学报:自然科学版,2011,31(5):400-407.
[8]Joshi S,Bharucha C,Desai A J,et al..Production of biosurfactant and antifungal compound by fermented food isolate Bacillus subtilis 20B [J].Bioresource Technol,2008,99(11):4603-4608.
[9]Nakarin S,Jaturong K,Boonsom B,et al.Biocontrol of Rhizoctonia solani AG-2,the causal agent of damping-off by Muscodor cinnamomi CMU-Cib 461 [J].World J Microbiol Biotechnol,2012,28:3171-3177.
[10]刘永峰.枯草芽孢杆菌(Bacillussubtilis)Bs-916 胞外抗菌蛋白质的纯化及其鉴定[D].南京:南京农业大学,2006.
[11]周艳芬,杜红方,袁洪水,等.棉花黄萎病拮抗蛋白的分离与纯化[J].棉花学报,2007,19(2):98-101.
[12]卢娟.拮抗香蕉枯萎病菌的LX1 菌株的分离鉴定及共抑菌蛋白的分离纯化与该基因的克隆[D].开封:河南大学,2011.
[13]孙瑶,李宝庆,马平.棉花黄萎茵拮抗细菌BDT-25 抑菌蛋白的分离纯化[J].河北农业科学,2011,15(6):42-45.
[14]贾翠英,张玉辉,马晓况.一株拮抗细菌所产抑菌物质的特性分析及初步鉴定[J].湖北大学学报:自然科学版,2012,43(9):378-383.
[15]陈爱香.细链格孢菌抗细菌枯草芽孢杆菌XL12 菌株发酵产抗菌蛋白条件研究[J].黑龙江农业科学,201l(9):42-44.
猜你喜欢
中国化肥信息(2022年6期)2023-01-06 05:36:01
湖南饲料(2021年4期)2021-10-13 07:32:46
中国化肥信息(2020年3期)2021-01-07 02:12:58
扬子江(2019年3期)2019-05-24 14:23:10
中国化肥信息(2018年8期)2018-10-08 03:17:10
天然产物研究与开发(2018年4期)2018-05-07 06:47:53
中成药(2018年1期)2018-02-02 07:20:03
中国化肥信息(2014年37期)2014-02-27 13:41:26
食品科学(2013年14期)2013-03-11 18:25:13
食品科学(2013年13期)2013-03-11 18:24:34
