
干旱、高温胁迫下大豆SSR标记与产量性状关联分析
2015-06-22 15:02柏锡梁爽邢雪莹东北农业大学生命科学学院哈尔滨150030
东北农业大学学报 2015年10期
柏锡,梁爽,邢雪莹(东北农业大学生命科学学院,哈尔滨150030)
干旱、高温胁迫下大豆SSR标记与产量性状关联分析
柏锡,梁爽,邢雪莹
(东北农业大学生命科学学院,哈尔滨150030)
研究49对SSR标记对36份在干旱、高温处理下的大豆材料多态性扫描和群体多样性分析。STRUCTURE2.3.2软件群体结构分析,Tassel2.1软件MLM模型对始花期等5个农艺性状关联分析,发掘优异等位变异。结果表明,①49个具有多态性的标记共检测出156个等位变异;标记位点的Shannon指数分布范围0.4506~1.3265,多态性信息值PIC分布范围0.2454~0.9059。②36份大豆材料被分为2个亚群。关联分析发现12个位点与大豆产量性状相关,研究各个关联位点找到Satt575-2、Sat_201-1、Satt686-2等表型效应明显的优异等位变异,为后期杂交育种亲本选择以及标记辅助育种提供基础。
大豆;SSR;遗传多样性;群体结构;关联分析;等位变异
作物耐旱性是指作物本身在干旱逆境条件下仍具有生长生存和形成稳定产量能力[1],由其遗传特征及与自然环境之间相互作用决定。大豆耐旱性是复杂的数量遗传性状,在进化过程中形成相对独特的抵御和适应干旱机制。主要分为两种:①避旱(逃逸)机制:一般为早熟品种,生长期较短,始花天数少,能够避开干旱敏感期和不利环境,或在干旱来临时提前完成生长周期;还有一类能够在干旱胁迫下仍保持较高水势,从根系、叶片以及主茎形态上可辨,根系发达,可从深层土壤吸取更多水分,主茎表皮毛浓密,保水能力强,并且可通过卷曲叶片降低水分散失,以达到躲避干旱胁迫目的,这类机制称为生态耐旱性。②耐旱机制:这类大豆品种能忍受干旱,干旱环境中依然可以维持正常生长状态和生理代谢,称为生理耐旱性[2-3]。
近年来,随着SSR标记技术迅速发展,多种分子标记已被广泛应用于大豆产量性状研究和关联分析[4-6]。Wen等对大豆农艺性状、品质性状与SSR标记关联分析[7];范虎等对百粒重、开花期、成熟期等与SSR标记关联分析[8]。但在干旱高温条件下的产量性状关联分析研究很少,本研究利用49对SSR标记引物多态性指数、等位变异以及36份大豆材料的群体结构、产量相关性状、生理指标,对36份在高温、高温干旱处理下的大豆材料综合分析,寻找关联位点,对极显著关联位点表型效应比较分析,筛选出一批具有优异等位变异的材料,为杂交亲本选择及后续标记辅助育种提供理论基础。
1 材料与方法
1.1 胁迫处理
36份材料2013年5月种植于哈尔滨市东北农业大学香坊实验实习基地,行长2 m,行距0.5 m,株距0.2 m,大棚中高温处理及干旱高温双重处理,大棚外正常栽培,利用大棚内温室效应自然升温达到对大豆高温处理,适当通风维持内外温度差为10℃,控制灌溉法将高温干旱双重处理的区域含水量确保在15%以下,高温处理及正常栽培则在40%。始花期、株高、百粒重、分枝数、总荚数统计参照文献[9]。
1.2 引物筛选
在大豆20个连锁群上,均匀随机选取112对 SSR引物,从大豆资源数据库(http://soybase.org/)中下载SSR引物序列,由哈尔滨博莱仕合生物科技有限公司合成。
1.3 分子数据收集
采用CTAB法提取DNA,PCR反应体系:10× Buffer for page 2μL;dNTPs(10 mmol·L-1)1.6μL;Sense primer(10μmol·L-1)0.6μL;Anti-sense primer(10μmol·L-1)0.6μL;Easy Taq DNA Polymerase for page(5 U·μL-1)0.2μL,DNA模版1 μL;灭菌ddH2O 14μL。PCR反应条件:94℃10 min;(94℃30 s,x℃×20 s,72℃30 s)30个循环;72℃10 min;4℃终止反应。x为不同引物相应的退火温度。
PCR反应完成后采用8%非变性聚丙烯酰胺凝胶电泳,银染,试剂均购自北京全式金生物技术有限公司。
1.4 数据分析
利用POPGENE1.32软件群体遗传多样性分析,包括Shannon指数、等位变异数等,根据各微卫星位点等位基因频率计算各位点多态信息含量值PIC=1-公式中pi为第i个等位基因出现的频率。利用Structure2.3.2进行群体遗传结构分析,估计最佳亚群数K,K取值1~10,将MCMC(Markov chain monte carlo)开始时不作数迭代设为10 000,再将不作数迭代后的MCMC设为10 000,迭代次数设为20次重复,依据ΔK估计试验材料亚群数目,对群体分类,并计算Q值[10]。利用SPAGeDi-1.3d处理基因型数据得到个体间亲缘关系kinship矩阵。利用Tassel2.1软件混合线性模型(Mixed linearmodel,MLM,Q+K模型),将亲缘关系kinship矩阵和群体结构Q值作为协变量,选择计算每个标记遗传力(Calculate heritability for each marker)的方式,选择EMMA(Efficient mixedmodelassociation)选项,作性状与标记关联分析,并计算每个标记位点对表型的变异解释率。根据SSR位点等位变异表型效应公式计算各等位变异表型效应值:式中,ai表示第i个等位变异表型效应值,xij为携带第i个等位变异的第j个材料表型值,ni为具有第i个等位变异材料数,nk为携带无效等位变异的第k个材料表型值,nk为具有无效等位变异材料数。
2 结果与分析
2.1 群体遗传多样性分析
利用112对引物对36份材料PCR扩增,筛选,其中49对引物具有多态性,多态性比例为43.75%。49对引物共检测出156个等位基因,变化幅度为2~4个,平均每个标记3.18个。SSR标记位点Shannon指数(Shannon's information index)分布范围为0.4506~1.3265,平均值为0.9662。Shannon指数最大标记为Sat_418,值为1.3265;最小标记为Satt679,值为0.4506;PIC值分布范围为0.2454~0.9059,平均值为0.6060,PIC指数最大标记为Sat_ 195,最小标记为Sct_187(见表1)。
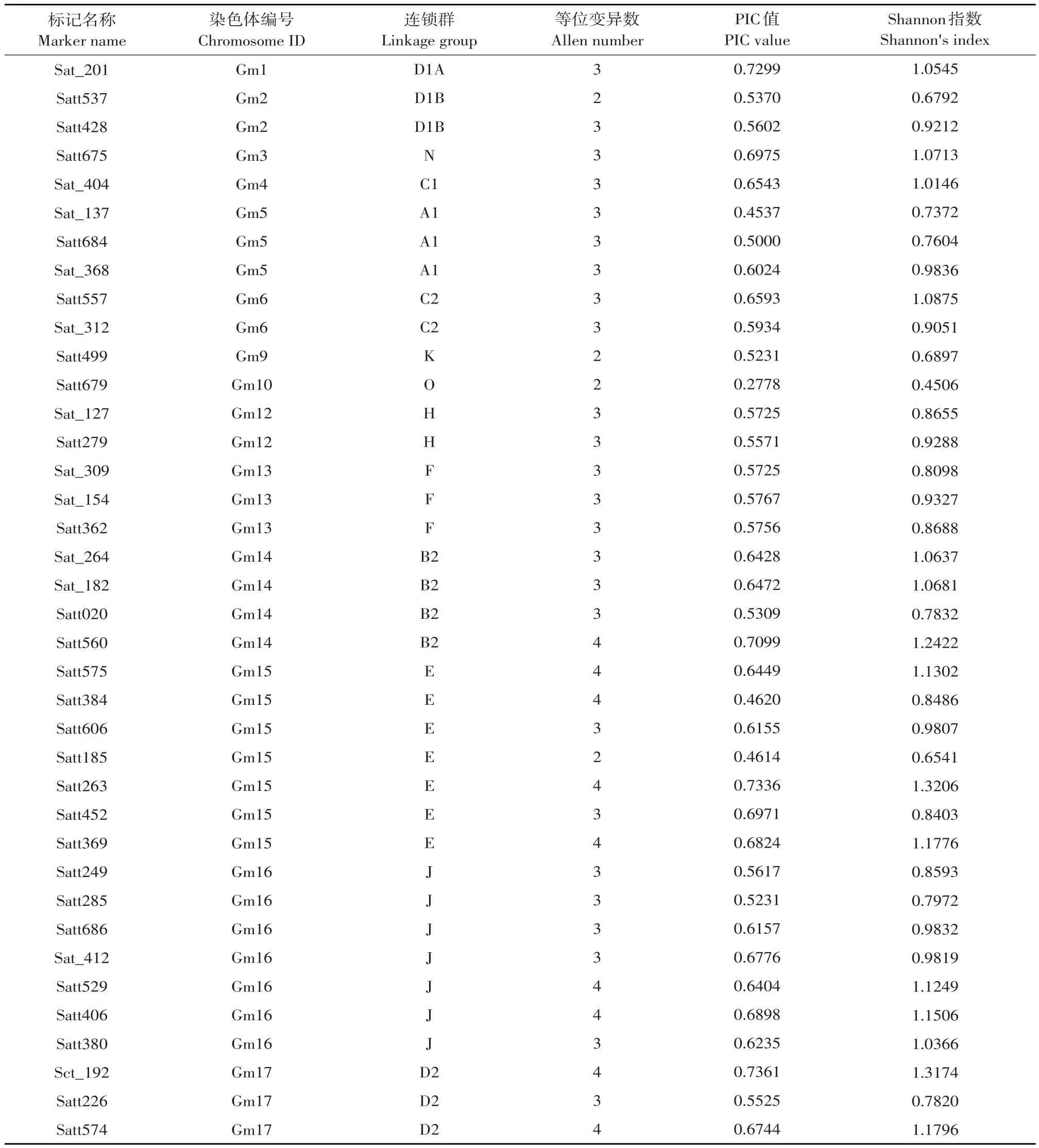
表1 49个SSR标记具体信息及其对36份大豆材料的多样性信息统计Table 1 Information of 49 SSR markers and their diversity statistic value within 36 materials
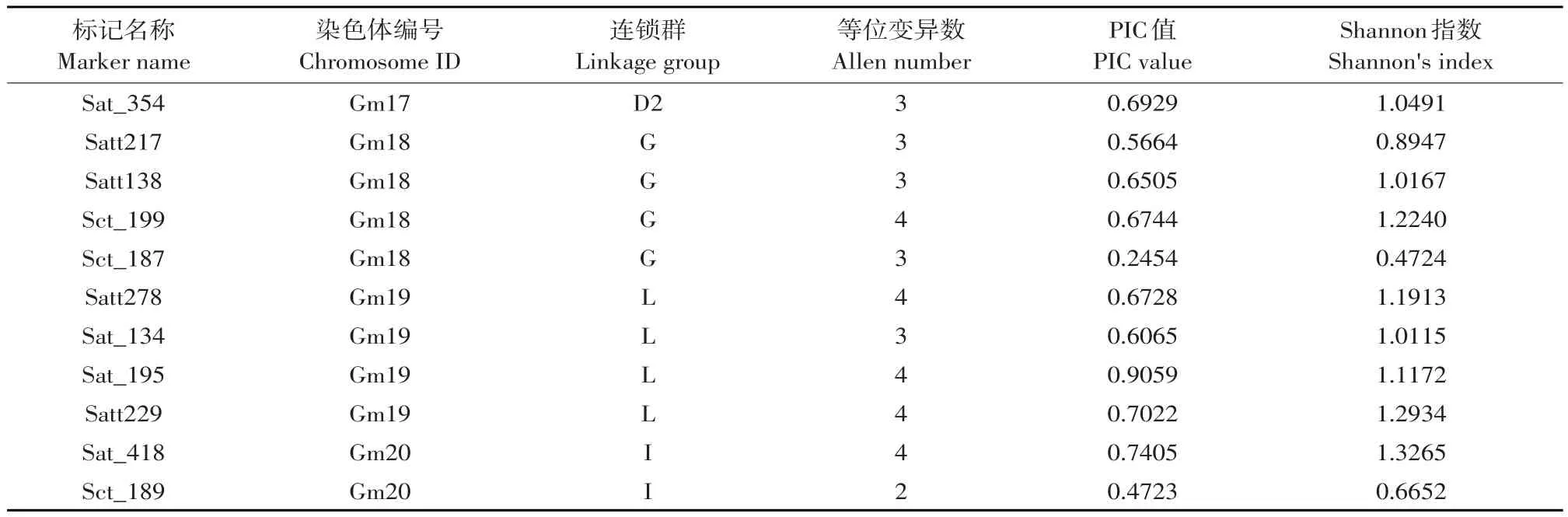
续表
2.2 群体遗传结构分析
选取49个SSR标记遗传信息,利用软件Structure2.3.2群体遗传结构分析,划分群类,并计算各材料Q值(第i个材料其基因组变异源于第k个群体的概率)。如图1所示,LnP(D)值随K值增加而增加,并未出现拐点,因此利用LnP(D)无法确定群体结构。采用ΔK判断最佳群组数,K=2时,ΔK出现最大值(见图2),由此确定材料群体最佳群组数为2,群体分为两个亚群。两亚群分别含22和14个材料,占群体总数比例分别为61%和39%,群体特点为占群体比例61%的亚群始花期较短,株高较低;而占群体比例39%的亚群始花期较长,株高较高。绘制36个材料群体结构图并统计材料相应Q值(见图3)。

图1 LnP(D)随K值变化折线图Fig.1 LnP(D)with the change of K value
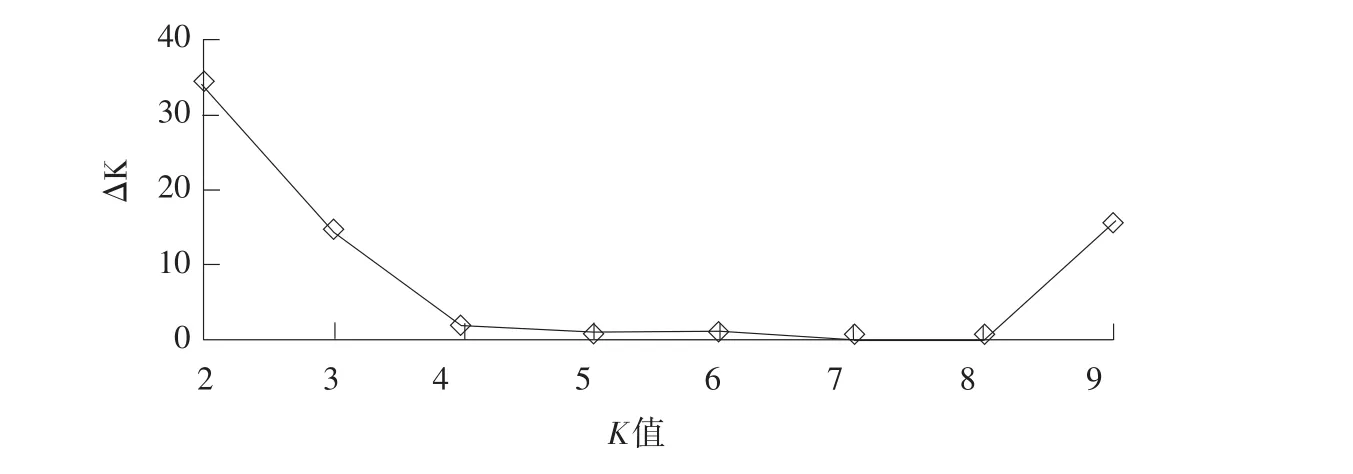
图2 ΔK随K值变化折线图Fig.2ΔK with the change of K value
2.3 SSR标记与大豆性状的关联分析
在群体遗传分析及个体间亲缘关系分析统计基础上,以每个个体相应Q值作协变量,利用Q+K方法对49个SSR标记及大豆产量相关性状和指标关联分析,探求与性状指标关联的标记,并计算相应性状指标变异解释率。本试验检测49个标记中与产量相关性状有12个(见表2)。
与产量性状相关的12个位点中,3个与总荚数相关,9个与始花期相关,未发现与株高、百粒重及分枝数相关的位点。在与总荚数相关的3个位点中,Sat_312与正常条件总荚数相关;Satt428、Satt686与干旱高温处理条件总荚数相关,说明在不同生长环境中,控制总荚数的位点会发生改变。
与始花期相关的9个位点中,Satt362、Satt575、Satt380、Satt452与正常条件下始花天数相关;Satt380、Satt452、Sat_201与高温条件下始花天数相关;Satt575、Satt185与高温干旱条件下始花天数相关。

图3 基于SSR标记的36份大豆材料的群体遗传结构(K=2)Fig.3 Population structure of 36 materials based on SSR markers(K=2)

表2 与产量性状相关标记位点及其对表型变异解释率(P<0.01)Table 2 Marker lociassociated with soybean yield traits and their explained phenotypic variation(P<0.01)
Satt575在正常与高温干旱条件下均与始花天数相关,高温干旱条件下解释率略大于正常条件,说明在高温干旱条件下该位点对表型影响更大。Satt380、Satt452在正常与高温条件下均与始花天数相关,说明高温条件不能改变上述位点作用,但Satt380在高温条件下解释率明显大于正常条件,说明虽高温胁迫未改变Satt380位点作用,但改变该位点对表型的影响程度,使影响程度变大;而Satt452在高温条件下解释率明显小于正常条件,说明胁迫使位点对表型影响程度减小。Satt185位点仅在高温干旱条件下与始花天数关联,在正常及高温条件下均与始花天数无关联,说明该位点作用仅在干旱高温胁迫下被激活。
2.4 关联位点优异等位基因变异分析
分析各关联位点等位变异基因与表型效应之间联系,从而确定优异等位基因(见表3、4)。正常种植环境下始花天数关联位点Satt362、Satt575、Satt380和Satt452的等位变异表型效应范围分别为-14.98~-11.58、-5.18~16.82、-4.20~9.59、-2.85~0.15,变异极明显,4个位点表型效应最小值分别为Satt362-2(-14.98)、Satt575-3(-5.18)、Satt380-1(-4.20)、Satt452-1(-2.85)。高温条件下始花天数关联位点Satt380、Satt452、Sat_201的等位变异表型效应范围为-6.25~8.18、-7.77~-4.66、9.82~10.86,变异极明显,3个位点表型效应最小值分别为Satt380-1(-6.25)、Satt452-2(-7.77)、Sat_201-2(9.82)。高温干旱条件下始花天数关联位点Satt575、Satt185的等位变异表型效应范围为-4.36~ 19.23、8.86,变异极为明显,Satt575位点表型效应最小值为Satt575-3(-4.36)。
正常种植环境下总荚数关联位点Sat_312表型变异范围为-1.77~38.07。干旱条件下总荚数关联位点Satt428、Satt686表型变异范围分别为-11.86~9.38、-9.61~32.68。可根据以上优异等位变异筛选优质亲本,为后续杂交育种及标记辅助育种提供参考。
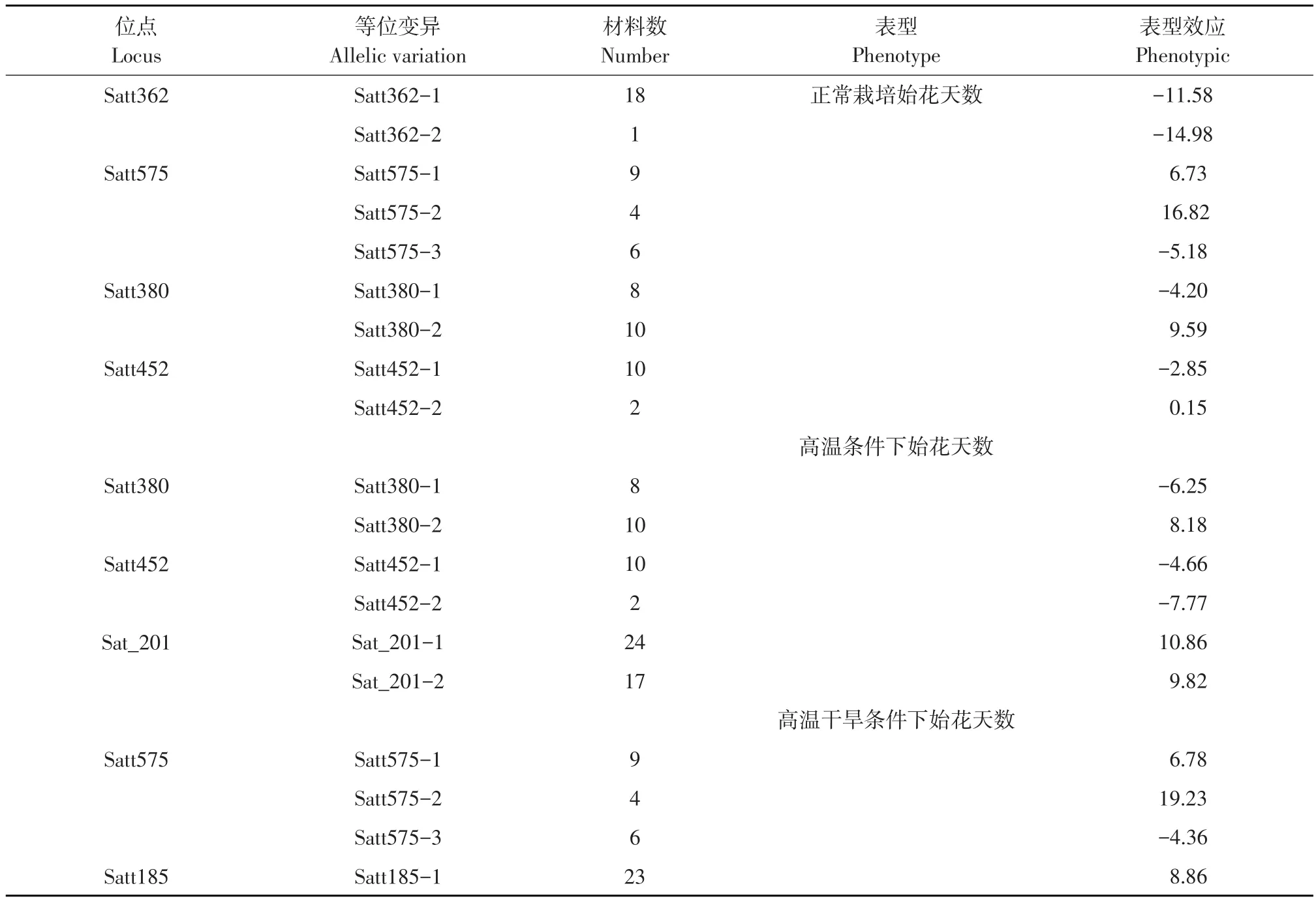
表3 关联位点不同等位变异基因与始花天数表型效应关系Table 3 Relationship between differentallelic variation genes of associated lociand phenotype effects of flowering time
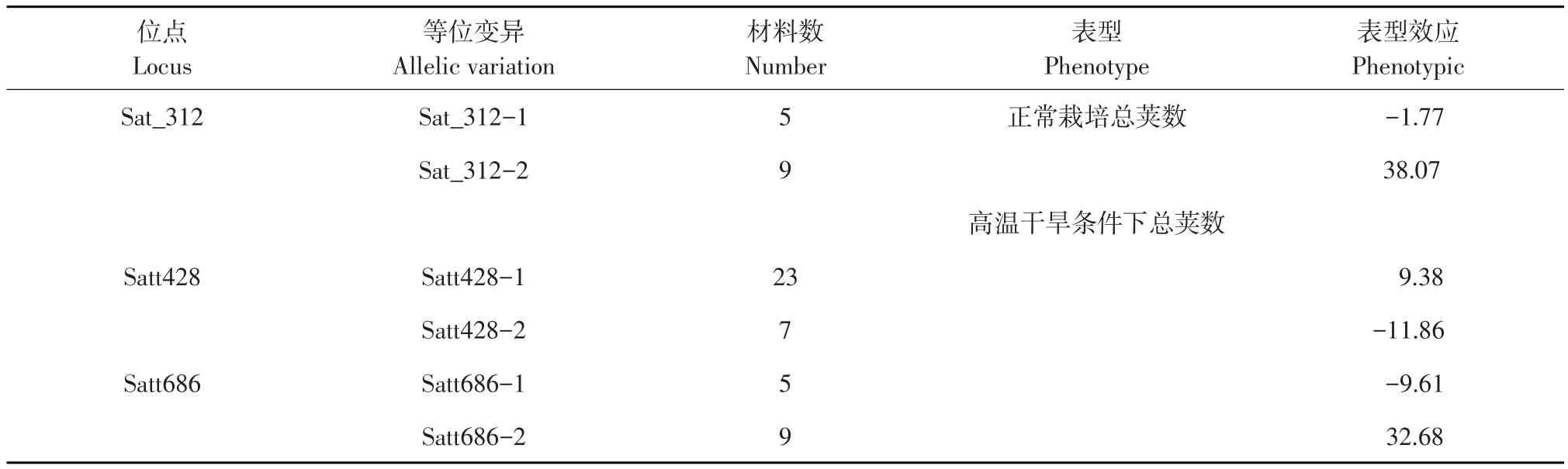
表4 关联位点不同等位变异基因与总荚数表型效应关系Table 4 Relationship between different allelic variation genes of associated lociand phenotype effects ofthe number of pods
3 讨论与结论
本研究中所得12个相关性位点中,Satt362与萎蔫程度相关[11];Satt575、Satt185、Satt428、Sat_312与产量相关[12-13];Satt380与始花期、结荚期相关[14];Satt452与成熟期、产量相关[15-16];Sat_201与Satt686暂无信息。
本研究中正常种植环境条件下与始花期相关的4个位点为Satt362、Satt575、Satt380、Satt4524,其中Satt380与数据库结果一致;Satt452在数据库中与成熟期相关,而成熟期与花期关联密切[17],该位点结果与前人研究基本一致;未发现Satt575、Satt362与始花期相关的报道,上述2个位点对相应表型的作用机制是否为新的QTL有待深入研究。
高温种植环境条件下与始花期相关的3个位点为Satt380、Satt452、Sat_201;其中位点Satt380、Satt452与数据库中公布信息一致,且这2个位点与正常种植环境条件下始花天数相关,说明高温胁迫对始花期造成的变化并未影响上述两个位点与表型的关联,但高温胁迫对表型变异解释率有一定影响,Satt380表型变异解释率变大、Satt452变小。Sat_201在数据库中未公布任何关联性状,是否为新的QTL有待验证。
高温干旱种植环境下与始花期相关的2个位点为Satt575和Satt185,在数据库中均与产量相关,可能是在高温干旱胁迫条件下上述2个位点作用发生改变,但具体发生改变所需胁迫强度及改变过程所涉及途径和机制,还需进一步研究。
正常种植环境下与总荚数相关的位点Sat_312在数据库中与产量相关,所得结果与数据库中一致。
高温干旱种植环境下与总荚数相关的2个位点为Satt428、Satt686,其中Satt428与数据库中公布结果一致;Satt686未公布任何关联性状,是否为新的QTL有待验证。
对关联分析得到的位点作优异等位变异分析,得到正常种植环境条件下对始花天数减效潜力明显的等位变异Satt362-2(-14.98)、Satt575-3(-5.18)、Satt380-1(-4.20)、Satt452-1(-2.85);在高温种植环境下对始花天数减效潜力明显的等位变异Satt380-1(-6.25)、Satt452-2(-7.77);以及在高温干旱种植环境条件下对始花天数减效潜力明显的等位变异Satt575-3(-4.36)。得到正常种植环境条件下对总荚数增效潜力明显的等位变异Sat_312-2(38.07);在高温干旱种植环境条件下对总荚数增效潜力明显的等位变异Satt428(9.38)、Satt686-2(32.68),选取含有以上等位变异的材料可缩短大豆始花天数,增加大豆成熟期总荚数,提高大豆产量。
[1]Ainsworth E A,Yendrek C R,Skoneczka J A,et al.Accelerating yield potential in soybean potential targets for biotechnological improvement[J].PlantCell Environment,2012,35(1):38-52.
[2]李慧峰,李贵全,张海燕.SSR标记在晋大52×晋大57后代群体分析中的应用[J].山西农业大学学报,2006,26(4):321-324.
[3]王丽侠,程须珍,王素华,等.应用SSR标记对小豆种质资源的遗传多样性分[J].作物学报,2009,35(10):1858-1865.
[4]He S,Wang Y,Volis S,et al.Genetic diversity and population structure:Implications for conservation of wild soybean based on nuclear and chloroplast microsatellite variation[J].International Journalof Molecular Sciences,2012,13(10):12608-12628.
[5]Lightfoot D A,Iqbal M J.Molecular mapping and breeding with microsatellite markers[J].Methods in Molecular Biology,2013, 1006(4):297-317.
[6]Galeano C H,Cortés A J,Fernández A C,et al.Gene-based single nucleotide polymorphism markers for genetic and association mapping in common bean[J].BMC Genetics,2012,13 (1):48.
[7]Wen Z X,Zhao T J,Zheng Y Z,et al.Association analysis of agronomic and quality traits with SSR markers in Glycine max and Glycine soja in China:I.Population structure and associated markers[J].Acta Agronomica Sinica,2008,34(7):1169-1178.
[8]范虎,文自翔,王春娥,等.中国野生大豆群体农艺加工性状与SSR关联分析和特异材料的遗传构成[J].作物学报,2013,39(5): 775-788.
[9]邱丽娟,常汝镇.大豆种质资源描述规范和数据标准[M].北京:中国农业出版社,2006:9-31.
[10]赵旭,方永丰,王汉宁.玉米SSR标记杂优类群划分及群体遗传结构分析[J].核农学报,2013,27(12):1828-1838.
[11]Charlson D,Bhatnagar S,King C,Polygenic inheritance ofcanopy wilting in soybean[J].Theor Appl Genet,2009,119(4):587-594.
[12]Wang D,Graef G L,Procopiuk A M,et al.Identification ofputative QTL that underlie yield in interspecific soybean backcross populations[J].Theor ApplGenet,2004,108(3):458-467.
[13]Reinprecht,Y,Poysa,V,Yu,K,etal.Seed and agronomic QTL in low linolenic acid,lipoxygenase-free soybean(Glycine max(L.) Merrill)germplasm[J].Genome,2006,49(12):1510-1527
[14]Tasma IM,Lorenzen L L,Green D E,etal.Mapping genetic loci for flowering time,maturity,and photoperiod insensitivity in soybean[J].Mol Breed,2001,8(1):25-35.
[15]Rossi M,Orf J,Liu L,etal.Genetic basis ofsoybean adaptation to North American vs.Asian mega-environments in two independent populations from Canadian×Chinese crosses[J].Theor Appl Genet,2013,126(7):1809-1823.
[16]宁慧霞,李英慧,刘章雄,等.大豆品种成熟期基因型推测的研究[J].作物学报,2008,34(3):382-388.
[17]樊翠芹.有限结荚类型大豆初花期与成熟期主要性状的遗传与相关分析[J].河北农业技术师范学院学报,1996(2):20-24.
Association analysis for soybean yield traits under drought and heat stress with soybean SSR markers
BAI Xi,LIANG Shuang,XING Xueying
(School of Life Sciences,NortheastAgriculturalUniversity,Harbin 150030,China)
In this research,the diversity of population and complete the polymorphism scan of 36 copies of soybeans were analyzed by using 49 pair of SSR markers.Furthermore,STRUCTURE 2.3.2 was used to analyze the group structure,Tassel2.1 and MLM model were used to perform the correlation analysis of five yield related traits including flowering time.Based on the analysis,excellent allelic variation can be explored.The following results were obtained.①There were 156 allelic variation according to the analysis of the 49 SSR markers;the Shannon index of these marked loci varied from 0.4506 to 1.3265,the polymorphism index contents(PIC)had a distribution range from 0.2454 to 0.9059.②36 copies of soybean materials are divided into two subgroups.Correlation analysis showed thatthere were 12 ofthe lociwere associated with the yield traits,Satt575-2,Sat_201-1,Satt686-2 and other excellent allelic variation were found to have the obvious phenotypic effect. These results could supply basis for the further mapping and the choice ofhybrid breeding parents.
soybean;SSR;genetic diversity;population structure;association analysis;allelic variation
S565.1
A
1005-9369(2015)10-0015-08
时间2015-10-29 13:40:59[URL]http://www.cnki.net/kcms/detail/23.1391.S.20151029.1340.014.html
柏锡,梁爽,邢雪莹.干旱、高温胁迫下大豆SSR标记与产量性状关联分析[J].东北农业大学学报,2015,46(10):15-22.
Bai Xi,Liang Shuang,Xing Xueying.Association analysis for soybean yield traits under drought and heat stress with soybean SSR markers[J].Journal of Northeast Agricultural University,2015,46(10):15-22.(in Chinese with English abstract)
2015-02-09
国家自然科学基金项目(31371544)
柏锡(1975-),男,副教授,博士,研究方向为植物逆境生理与基因工程。E-mail:baixi@neau.edu.cn
猜你喜欢
作物学报(2022年6期)2022-04-08
国际医学放射学杂志(2021年5期)2021-10-22
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
麦类作物学报(2018年4期)2018-05-11
现代园艺(2017年21期)2018-01-03
第一财经(2017年36期)2017-09-25
中国康复理论与实践(2015年10期)2015-12-24
百科知识(2015年18期)2015-09-10
医学研究杂志(2015年5期)2015-06-10
