
蜘蛛类天敌与茶尺蠖幼虫空间关系的聚块样方方差分析
2015-06-12 12:36刘飞飞柯胜兵王建盼毕守东邹运鼎周夏芝党凤花徐劲峰禹坤赵学娟
浙江大学学报(农业与生命科学版) 2015年2期
刘飞飞, 柯胜兵, 王建盼, 毕守东*, 邹运鼎, 周夏芝, 党凤花, 徐劲峰, 禹坤, 赵学娟
(1.安徽农业大学理学院,合肥 230036;2.安徽省潜山县植保站,安徽 潜山 246300)
蜘蛛类天敌与茶尺蠖幼虫空间关系的聚块样方方差分析
刘飞飞1, 柯胜兵1, 王建盼1, 毕守东1*, 邹运鼎1, 周夏芝1, 党凤花1, 徐劲峰2, 禹坤1, 赵学娟1
(1.安徽农业大学理学院,合肥 230036;2.安徽省潜山县植保站,安徽 潜山 246300)
用聚块样方方差分析法、灰色关联度分析法、空间格局聚集强度指数法和ρ指数法对安徽省潜山县茶园茶尺蠖及其8种蜘蛛之间的关系进行分析。结果表明:与茶尺蠖空间上跟随关系密切的天敌是三突花蟹蛛(关联度值为0.856 8)和八点球腹蛛(0.788 8),跟随关系较不密切的天敌是鞍形花蟹蛛(0.631 2)和日本球腹蛛(0.569 8);聚块样方方差分析和聚集强度分析表明,在聚块内有1~32个基本样方时,随着聚块内基本样方数的增多,聚集格局的扩散系数C值不断增大,均匀和随机格局的扩散系数C值不断减小;茶尺蠖及其蜘蛛在聚块基本样方数为2、4、8、16和32时与为1时之间的空间分布聚集程度差异均不显著;茶尺蠖的种群聚集均数λ在多数情况下大于2,聚集是由其本身原因造成的,在种群聚集均数为正值时,随着聚块内基本样方数的增加,种群聚集均数不断增大;用茶尺蠖不同大小聚块的ρ指数判断其聚集时个体群的最小范围表明,茶尺蠖个体群聚集格局的最小面积是聚块中具有1个基本样方,即该文的1.7 m2,这为对该害虫抽样时确定样方大小提供了科学依据。
茶尺蠖; 蜘蛛类天敌; 空间关系; 聚块样方方差分析; 灰色关联度分析
Summary The relationships between insect pests and their natural enemies include the quantitative relations of their occurrence, the temporal relationships of their occurrence as well as their spatial distribution patterns. In previous studies, the quantitative relationships between tea pests and their spider natural predators have been researched by grey correlation degree analysis. In addition, the niche overlap index and niche similarity coefficient analysis methods have been used to assess the natural predators of pests, which studied the spatial and temporal relationships of the insect pests and their natural predators. Moreover, the ranges of spatial dependence (RSDs) of natural enemies and insect pests were respectively calculated by the method of geostatistics. Then the RSDs were analyzed using grey correlation degree analysis to systematically investigate the spatial relationships between pests and enemies.
Spiders are the major natural enemies ofEctropisobliquehypulina. Therefore, scientifically utilizing and conserving spider natural enemies for integrated insect pest control of tea is necessary, and can provide scientific basis for evaluating the dominant natural enemies ofE.obliquehypulina. In this work, the close degree of spatial following relationship betweenE.obliquehypulinaand its natural enemies was examined. And the sampling size of quadrat ofE.obliquehypulinain tea garden in Qianshan area, Anhui Province was estimated by blocked quadrat variance analysis, grey relation analysis, aggregated-intensity index method of spatial patterns andρindex method.
The results showed that the most closely followed spider natural enemies ofE.obliquehypulinawereMisumenopstricuspidatusandTheridionoctomaculatum, with correlation degree values of 0.856 8 and 0.788 8, respectively, whilst the less close following relationships were found to beXysticusephippiatusandEnoplognathajaponica, with correlation degree values of 0.631 2 and 0.569 8, respectively. Combining the blocked quadrat variance analysis with aggregated-intensity analysis, the results showed the diffusion coefficientC-value of aggregation pattern increased with the increase of basic quadrats when there were 1-32 basic quadrats in the poly-block. But as for uniform and random pattern, it was continuous decrease. When the numbers of basic quadrat within the poly-block were 2, 4, 6, 8, 16 and 32 as compared with 1, the gathering degree of spatial distribution ofE.obliquehypulinaand its spider predators was not significant difference. In the most cases, the averageλvalues ofE.obliquehypulinawere more than 2, indicating that the gathering was caused by the pest itself. When theλvalues were positive, along with the increase of quadrats in the poly-block, the gathering averages were continuously increasing. The smallest scope of individual population aggregation was evaluated by usingρindex ofE.obliquehypulinaunder different sizes of poly-blocks. As a result, the minimum area of individual population aggregation pattern ofE.obliquehypulinahad one basic quadrat with 1.7 m2in this study, which provided a scientific basis for determining the size of quadrat on the pests sampling.
茶尺蠖(Ectropisobliquehypulina)是我国主要茶树害虫之一,广泛分布于各主要茶区,在长江中下游茶区尤以苏、浙、皖接壤地区危害严重;国外分布于日本。茶尺蠖幼虫取食常使茶丛光秃,同时还危害大豆、豇豆、芝麻、向日葵、辣蓼等植物。在安徽省宣城地区1年发生6代,浙江省杭州市1年发生7代[1]。温度对茶尺蠖繁殖力的影响明显,最适于21~26 ℃发生,繁殖力最强;大于26 ℃时,虫口急剧下降;大于29 ℃时生活力明显减弱[2]。茶尺蠖的天敌种类较多,茶园蜘蛛是茶尺蠖捕食性天敌的主要类群,三突花蟹蛛(Misumenopstricuspidatus)日捕食其1~2龄幼虫1.4~1.7头,鞍形花蟹蛛(Xysticusephippiatus)日捕食1~3头[3-5],幼虫期茶尺蠖绒茧蜂(Apantelessp.)、单白绵绒茧蜂(Apantelessp.)等的寄生率均在20%~30%,最高达70%以上[6]。许宁等[7]分离和鉴定了引诱茶尺蠖天敌寄生蜂的茶树挥发物种类;张xx正群等[8]研究了14种植物精油对茶尺蠖行为的影响;叶恭银等[9]研究表明茶尺蠖核型多角体病毒对宿主种群有一定的控制作用;曹士先等[10]基于cDNA扩增片段长度多态性发掘了茶树被茶尺蠖取食诱导的相关差异基因及其表达特征;任红楼等[11]研究了5种新型低毒农药对茶尺蠖的田间防控效果;孙钦玉等[12]研究了在进行生物防治、物理防治的同时,使用15%茚虫威、2.5%联苯菊酯、22%阿立卡等对茶尺蠖的防治效果。天敌与害虫之间在数量、时间、空间方面跟随关系的密切程度直接与天敌对害虫的控制作用大小有关。本文尝试用聚块样方方差分析法、灰色关联度分析法、聚集程度指数法和ρ指数法,综合分析天敌对茶尺蠖幼虫在空间上跟随关系的密切程度、种群聚集原因和个体群聚集的范围大小,为茶尺蠖幼虫的天敌优势种评定和抽样样方大小的确定提供科学依据。
1 材料与方法
1.1 供试茶园概况
供试茶园为安徽省潜山县园艺茶场(30°41′ N,116°34′ E),属小平原,面积25.3 hm2,海拔46 m;单作,茶树品种为8 年生天柱山群体种,行距1.7 m。茶园管理水平精细,茶树生长茂密,试验区茶园一直不施用化学农药。
1.2 调查方法
采用平行跳跃法。随机取10行,每行间隔2 m取一个1 m长的样方,共取104个样方。盆拍前,先目测,然后每样方随机选取10片叶,调查一些不易振落的害虫和天敌种类,然后用洗衣粉水液粘虫对样方中所有枝条进行盆拍,调查记载害虫及其天敌物种数和个体数。
1.3 调查时间
2009年4月15日至10月28日,15~20 d调查1次,共11次。
1.4 数学分析方法
1.4.1 茶尺蠖与其天敌空间动态的聚块样方方差分析 聚块样方方差分析[13]是在不同大小样方上的方差分析,是一种简单、有效的生态学空间格局分析方法。该方法要求供试田块上的样方在空间上相互连接,随着聚块所包含的基本样方数目从1,2,4,8,…,n(指数级数)不断增加,聚块方差值常随之改变,通过不同大小聚块方差值的变化,了解研究对象随尺度增大的变化动态。
在一样带上连续分布的样方,变量(茶尺蠖和其8种天敌在每个样方上的数量)为X,让聚块内基本样方数成指数增大,计算其均方差(mean square, MS)。当每聚块仅包含1个基本样方时,其均方差计算公式为
式中:MS(1)为聚块含1个基本样方时的均方差值;k为聚块所含基本样方数。
当每个聚块包含2个基本样方时,
依此类推,直到聚块所含的基本样方数为n/2,这时均方差的计算公式为
(Xi+1+Xi+2+…+Xi+n/2)]2。
聚块样方方差分析的目的是分析聚块大小对方差的影响。如果均方差在某一聚块大小上出现峰值,则表明田间害虫及其天敌的空间分布具有规律性,害虫和其天敌聚集范围的空间大小大致等于峰值出现时聚块的大小。如果同时出现几个峰值,则表明田间害虫和其天敌可能存在几种不同尺度的聚集空间,本文以最大峰值为对象进行分析。如果均方差值为一常数,即不随聚块大小而变化,则表明害虫和其天敌聚集的空间大小是无规律的。
1.4.2 茶尺蠖与其天敌空间关系的灰色关联度分析[14]将茶尺蠖及其天敌均方差峰值的基本样方数(聚集空间大小)分别看作一个本征性灰系统,茶尺蠖每次峰值的基本样方数(聚集空间大小Yi)分别作为该系统的参照序列。将不同时点上的茶尺蠖聚集空间大小Yi与天敌峰值聚块的基本样方数(聚集空间大小)Xj在第k点上的效果白化值进行双序列关系分析:
Yi={Yi(1),Yi(2),…,Yi(n)},i=1,2,3,4,5;
Xj={Xj(1),Xj(2),…,Xj(n)},j=1,2,3,…,6。
经数据均值化后得:
Yi={yi(1),yi(2),…,yi(n)},i=1,2,3,4,5;
xj={xj(1),xj(2),…,xj(n)},j=1,2,3,…,6。
Yi与Xj在第k点上的关联系数rij(k)为
rij(k)=[min min|yi(k)-xj(k)|+ρmaxmax|yi(k)-xj(k)|]/[|yi(k)-xj(k)|+ρmaxmax|yi(k)-xj(k)|],k=1,2,3,…,n。
式中:ρ为分辨系数,取值区间[0,1],一般取ρ=0.5,为扩大各关联度之间的差异,本文取ρ=0.8;Δij(k)=yi(k)-xj(k)为yi与xj序列在第k点上的绝对值差;min|yi(k)-xj(k)|为1级最小差,表示找出yi与xj序列对应点差值中的最小差;而min min|yi(k)-xj(k)|为2级最小差,表示在1级最小差的基础上再找出其中的最小差;max|yi(k)-xj(k)|与max max|yi(k)-xj(k)|分别为1级和2级最大差,其含义与上述最小差相似。R(Yi,Xj)=1/n∑rij(k),为第j种天敌(Xj)与茶尺蠖聚集空间大小的关联度,其大小反映了天敌Xj对茶尺蠖Yi在空间上跟随的密切程度。关联度值越大,表明二者在空间上关系越密切。

2 结果与分析
调查结果表明:茶园共有52种害虫和37种捕食性天敌,茶尺蠖是主要害虫之一,其主要捕食性天敌蜘蛛有草间小黑蛛(Erigonidiumgraminicolum)、八点球腹蛛(Theridionoctomaculatum)、锥腹肖蛸(Tetragnathamaxillosa)、鞍型花蟹蛛、日本球腹蛛(Enoplognathajaponica)、茶色新圆蛛(Neosconatheisi)、三突花蟹蛛、斑管巢蛛(Clubionareichini);4月29日以后茶尺蠖幼虫数量迅速增加,6月下旬和10月下旬数量最多,即第3代和第6代发生量大;春夏季草间小黑蛛、八点球腹蛛和锥腹肖蛸数量较多,秋冬季三突花蟹蛛和斑管巢蛛数量较多(图1)。
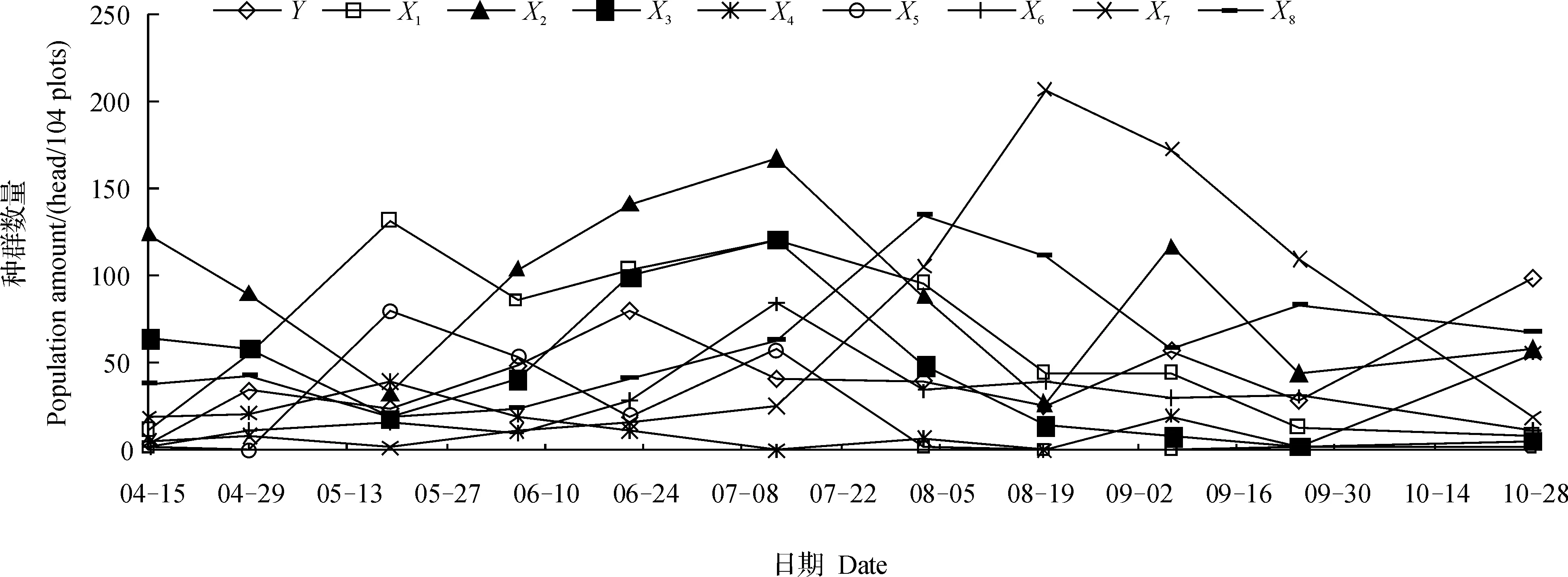
Y:茶尺蠖 (Ectropis oblique hypulina);X1:草间小黑蛛(Erigonidium graminicolum);X2:八点球腹蛛(Theridion octomaculatum);X3:锥腹肖蛸(Tetragnatha maxillosa);X4:鞍型花蟹蛛(Xysticus ephippiatus);X5:日本球腹蛛(Enoplognatha japonica);X6:茶色新圆蛛(Neoscona theisi);X7:三突花蟹蛛(Misumenops tricuspidatus);X8:斑管巢蛛(Clubiona reichini)。
2.1 茶尺蠖及其8种天敌空间关系的聚块样方方差分析
在一般情况下害虫数量多时害虫与天敌的动态可较准确地反映天敌与害虫之间关系的密切程度。本文选择6月6日、6月22日、7月13日、8月3日、9月7日和10月28日数量较多时的茶尺蠖及其8种主要天敌草间小黑蛛、斑管巢蛛、八点球腹蛛、三突花蟹蛛、日本球腹蛛、茶色新圆蛛、锥腹肖蛸和鞍形花蟹蛛的数据进行聚块样方方差分析(表1)。从中可以看出,茶尺蠖均方差峰值的基本样方数按时间先后依次是4、32、4、32、32和16个。为了分析天敌对茶尺蠖在空间上跟随关系的密切程度,对茶尺蠖及其8种天敌均方差峰值时的聚块内基本样方数(表2)进行灰色关联度分析,结果是三突花蟹蛛(X7)和八点球腹蛛(X2)与基本样方数的关联度值最大,分别为0.856 3和0.788 8,其余依次是锥腹肖蛸(X3)、斑管巢蛛(X8)、草间小黑蛛(X1)、茶色新圆蛛(X6)、鞍形花蟹蛛(X4)和日本球腹蛛(X5),关联度依次是0.770 4、0.711 0、0.644 9、0.631 2和0.569 8。说明三突花蟹蛛和八点球腹蛛对茶尺蠖在空间上的跟随关系最密切,鞍形花蟹蛛和日本球腹蛛跟随关系较不密切。
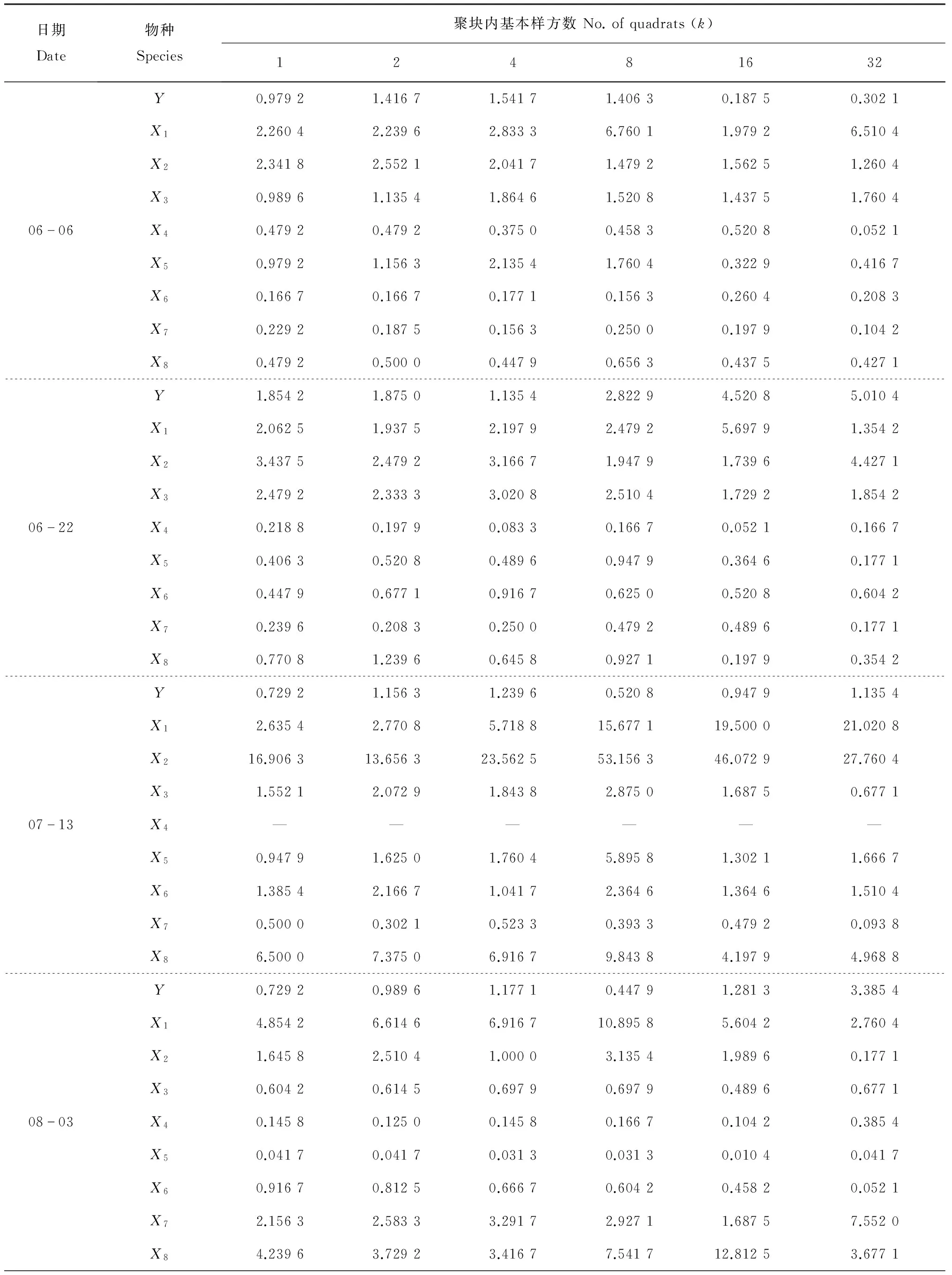
表1 茶尺蠖与其8种天敌在不同大小聚块下的均方差

续表1 茶尺蠖与其8种天敌在不同大小聚块下的均方差
Y,X1~X8代表的属种名详见图1注;—:无真值.
Please see the footnote of Fig.1 for details ofYandX1-X8. —: No truth-value.
表2 茶尺蠖及其8种天敌聚集分布格局范围(均方差峰值时的样方数)
Table 2 Scope of aggregation distribution pattern ofE.obliquehypulinaand eight species of natural enemies (the number of quadrats on the peak value of mean-variance)
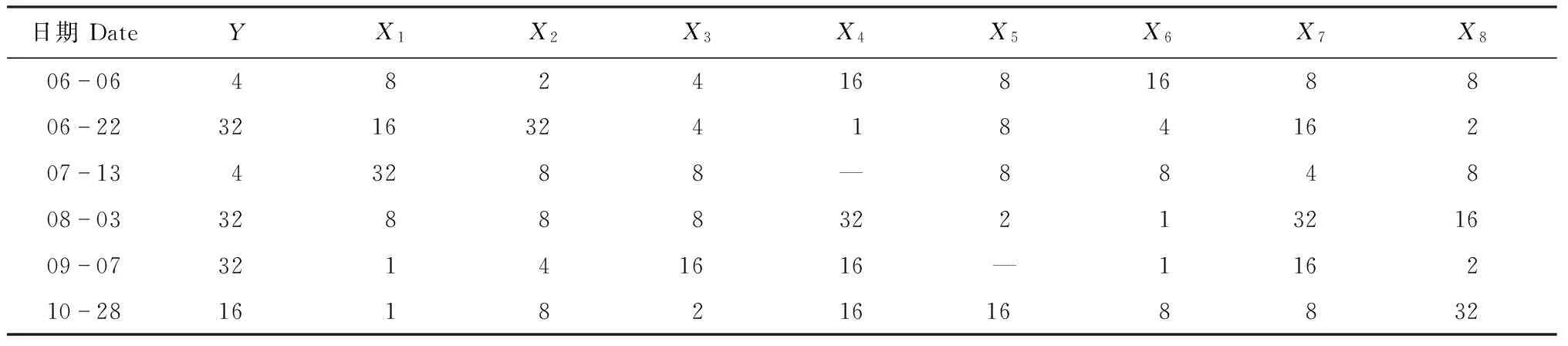
日期DateYX1X2X3X4X5X6X7X806064824168168806223216324184162071343288—884808033288832213216090732141616—116210281618216168832
Y,X1~X8代表的属种名详见图1注;—:无真值.
Please see the footnote of Fig.1 for the details ofYandX1-X8. —: No truth-value.
2.2 以不同大小聚块为单位的茶尺蠖及其天敌的空间格局
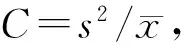

表3 茶尺蠖与其8种天敌在不同大小聚块的扩散系数C值
续表3 茶尺蠖与其8种天敌在不同大小聚块的扩散系数C值
Continuation Table 3 Diffusion coefficientC-value ofE.obliquehypulinaand eight species of natural enemies under different sizes of poly-blocks

物种Species自由度df聚块内基本样方数No.ofquadrats(k)日期Date060606220713080309071028X49511.02321.0383—1.22582.82151.52034720.83200.9728—1.16444.16991.89762340.98630.7550—1.63345.07852.16491181.20090.8843—1.70149.52713.88845161.56840.7455—1.85718.48013.77742320.36841.4545—4.42865.60000.1321X59511.20331.15791.42380.9910—1.00004721.33661.77302.04220.9779—1.00002341.91541.65222.25690.9569—1.00001181.32732.24243.94550.9089—1.00005160.94821.20001.63270.8000—1.00002320.94551.16672.03642.0000—1.0000X69510.90500.98401.00211.30541.17521.30904721.01291.39021.19571.10630.97641.21722341.23471.72571.42130.95650.79711.23471181.27271.91531.83640.77270.73741.27285162.32002.02071.46000.57500.77770.88002322.80002.72411.96250.22950.77780.4000X79510.89460.84191.01051.25452.07491.12974720.78710.93630.76601.63452.64301.34112340.75501.26081.04352.07163.76491.64521180.78511.54550.72722.49205.12462.80875160.74551.60000.80002.61186.88891.94732320.63640.80000.37505.32350.22222.5790X89511.07790.98405.63001.80240.92941.13054721.06081.39026.38411.99120.89691.53162341.02481.72575.96112.56730.62881.81251181.15581.91536.43544.18850.72731.48295161.22862.02073.65264.73640.39991.74752321.00002.72416.15793.97560.07692.2787
Y,X1~X8代表的属种名详见图1注;—:无真值.
Please see the footnote of Fig.1 for the details ofYandX1-X8. —: No truth-value.
n1=96-1=95,n2=∞,F0.05=1.000 0;n1=48-1=47,n2=∞,F0.05=1.49;n1=24-1=23,n2=∞,F0.05=1.55;n1=12-1=11,n2=∞,F0.05=1.81;n1=6-1=5,n2=∞,F0.05=2.21;n1=3-1=2,n2=∞,F0.05=3.00.
2.3 茶尺蠖及其天敌在不同聚块基本样方数时空间分布格局的聚集程度差异
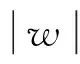
表4 天敌与茶尺蠖在聚块基本样方数为2、4、8、16和32时与为1个时的分布格局聚集程度差异|w|值
Table 4 Difference in spatial distribution pattern |w| value ofE.obliquehypulinaand its natural enemies between the basic blocks of 2, 4, 6, 8, 16, 32 and 1
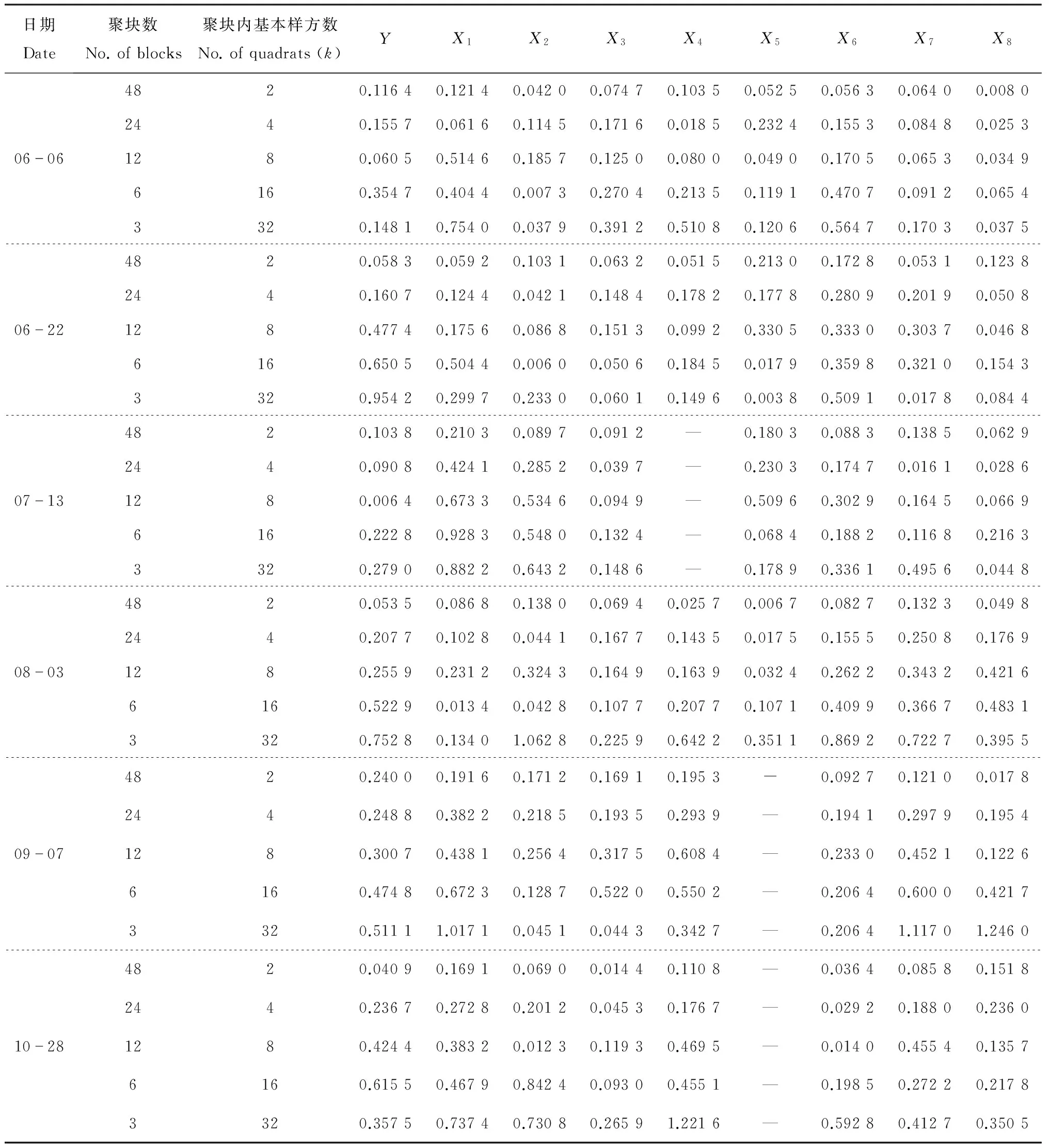
日期Date聚块数No.ofblocks聚块内基本样方数No.ofquadrats(k)YX1X2X3X4X5X6X7X806064820.11640.12140.04200.07470.10350.05250.05630.06400.00802440.15570.06160.11450.17160.01850.23240.15530.08480.02531280.06050.51460.18570.12500.08000.04900.17050.06530.03496160.35470.40440.00730.27040.21350.11910.47070.09120.06543320.14810.75400.03790.39120.51080.12060.56470.17030.037506224820.05830.05920.10310.06320.05150.21300.17280.05310.12382440.16070.12440.04210.14840.17820.17780.28090.20190.05081280.47740.17560.08680.15130.09920.33050.33300.30370.04686160.65050.50440.00600.05060.18450.01790.35980.32100.15433320.95420.29970.23300.06010.14960.00380.50910.01780.084407134820.10380.21030.08970.0912—0.18030.08830.13850.06292440.09080.42410.28520.0397—0.23030.17470.01610.02861280.00640.67330.53460.0949—0.50960.30290.16450.06696160.22280.92830.54800.1324—0.06840.18820.11680.21633320.27900.88220.64320.1486—0.17890.33610.49560.044808034820.05350.08680.13800.06940.02570.00670.08270.13230.04982440.20770.10280.04410.16770.14350.01750.15550.25080.17691280.25590.23120.32430.16490.16390.03240.26220.34320.42166160.52290.01340.04280.10770.20770.10710.40990.36670.48313320.75280.13401.06280.22590.64220.35110.86920.72270.395509074820.24000.19160.17120.16910.1953-0.09270.12100.01782440.24880.38220.21850.19350.2939—0.19410.29790.19541280.30070.43810.25640.31750.6084—0.23300.45210.12266160.47480.67230.12870.52200.5502—0.20640.60000.42173320.51111.01710.04510.04430.3427—0.20641.11701.246010284820.04090.16910.06900.01440.1108—0.03640.08580.15182440.23670.27280.20120.04530.1767—0.02920.18800.23601280.42440.38320.01230.11930.4695—0.01400.45540.13576160.61550.46790.84240.09300.4551—0.19850.27220.21783320.35750.73740.73080.26591.2216—0.59280.41270.3505
Y,X1~X8代表的属种名详见图1注;—:无真值.
Please see the footnote of Fig.1 for the details ofYandX1-X8. —: No truth-value.
n=48,|w|=0.364 7;n=24,|w|=0.521 3;n=12,|w|=0.753 8;n=6,|w|=1.118 0;n=3,|w|=1.767 8.
2.4 天敌和茶尺蠖在不同大小聚块条件下种群聚集均数的变化
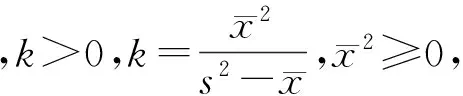

表5 不同大小聚块条件下茶尺蠖及其天敌的种群聚集均数λ值
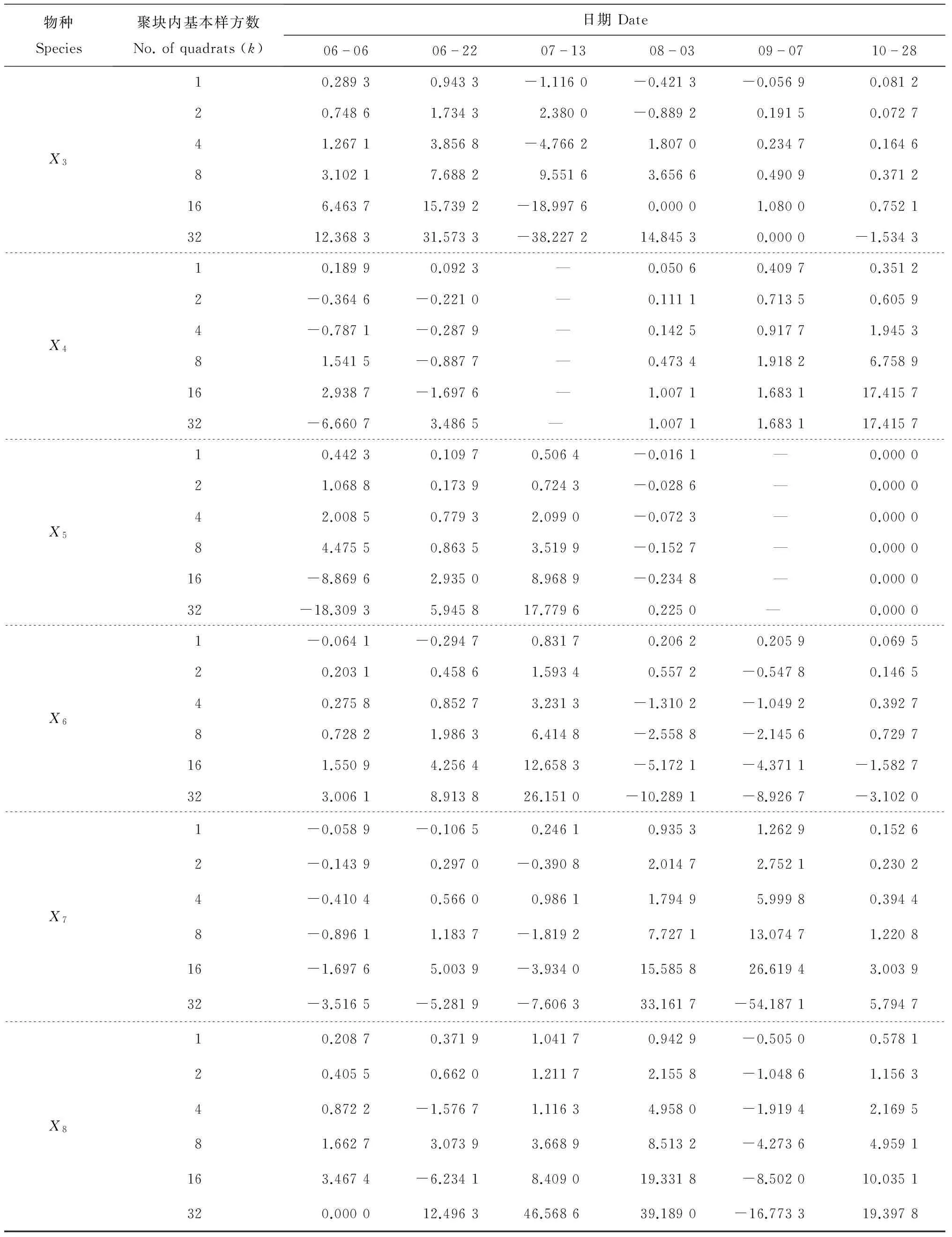
续表5 不同大小聚块条件下茶尺蠖及其天敌的种群聚集均数λ值
Y,X1~X8代表的属种名详见图1注;—:无真值.
Please see the footnote of Fig.1 for the details ofYandX1-X8. —: No truth-value.
2.5 不同大小聚块条件下茶尺蠖的ρ指数
对个体群聚集时占据的面积,Iwao[20]提出用ρ指数来测定。该方法只对生活在样方大小按规律增加的生境中的动物适用,ρ指数说明了样方大小范围根据个体群面积发生的急剧变化。当ρ指数随着样方大小范围(即本文中聚块的基本样方数)增大而变化,以ρ指数为纵轴,样方大小为横轴作图,其中ρ指数最大的2个样方之间即是该动物个体群聚集时占据的最小面积。不同聚块大小条件下茶尺蠖的ρ指数见表6。从中可以看出,聚块基本样方数由2到1时的ρ指数值最大,为1.547 8,表明茶尺蠖个体群处于聚集格局时占据的最小范围是聚块中具有1个基本样方。按照本文的调查方法计算,每一聚块面积应是1.7 m2,即取样调查茶尺蠖时每个样方应不小于1.7 m2。
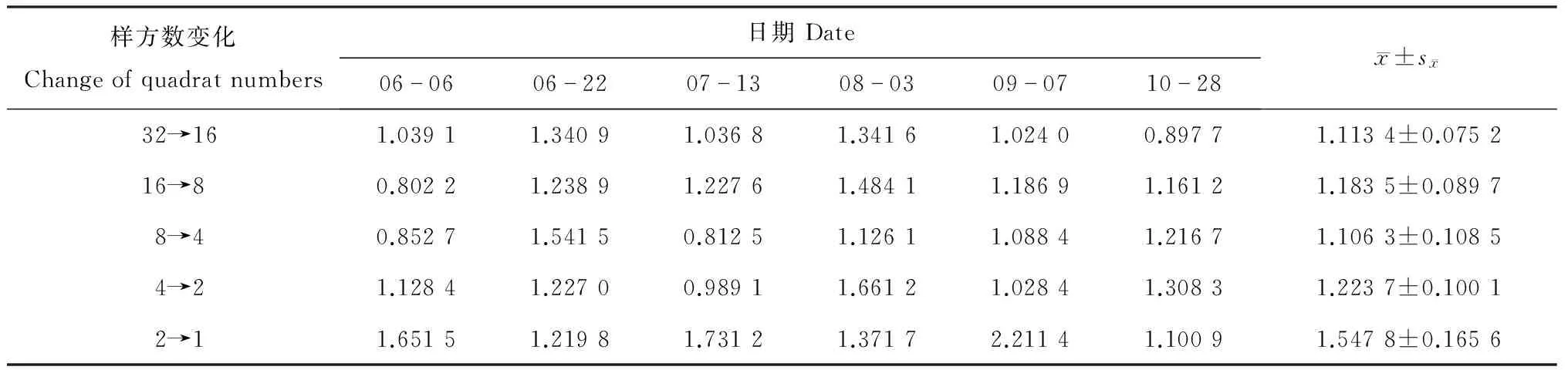
表6 不同聚块大小条件下茶尺蠖的ρ指数
3 讨论
天敌与害虫之间的空间关系是评定天敌优势种的重要内容[16]。目前关于天敌与害虫空间关系的研究,一是采用聚集强度指数如扩散系数C、久野指数CA等比较天敌和害虫的空间分布格局类型[21-22];二是利用地学统计学方法求得害虫和天敌半变异函数的变程,将二者变程大小进行比较或进一步对二者变程进行灰色关联度分析,比较二者之间关联度值的大小[23-33];第三种方法是分析害虫和天敌之间空间生态位重叠和相似情况,比较二者空间生态位重叠指数大小和相似系数大小[34]。本文采用聚块样方方差分析法分析害虫与天敌之间的空间关系,一是可以明确害虫处于聚集分布格局时个体群占据的范围大小,可为抽样时确定样方大小提供参考;二是可以通过害虫和天敌之间的均方差峰值时样方数的数学分析明确各种天敌在空间上对害虫跟随关系的密切程度,为综合评判害虫的天敌优势种提供科学依据。
4 结论
运用聚块样方方差分析法、灰色关联度分析法、聚集强度指数法和ρ指数法研究安徽省潜山县茶园天敌对茶尺蠖空间上的跟随关系和个体群占据的最小面积,其结果如下:
1)与茶尺蠖空间上跟随关系密切的天敌是三突花蟹蛛和八点球腹蛛;
2)鞍形花蟹蛛和日本球腹蛛与茶尺蠖空间上的跟随关系较不密切;
3)在聚块内基本样方数为1~32个时,随着聚块内样方数的增多,聚集格局的扩散系数C值一直在增大,均匀和随机格局的扩散系数C值不断减小;
4)茶尺蠖及其天敌在聚块内基本样方数为2、4、8、16和32时与为1个时的空间聚集程度差异均不显著;
5)茶尺蠖在不同聚块条件下的聚集是由该害虫本身原因引起的,在种群聚集均数为正值时,随着聚块中基本样方数的增多,种群聚集均数不断增大;
6)用不同大小聚块的ρ指数判断茶尺蠖个体群聚集时占据的最小面积为聚块中具有1个基本样方。
致谢 南京师范大学博士生导师马飞教授在论文撰写过程中给予了大力帮助,谨致谢意!
[1] 张汉鹄,谭济才.中国茶树害虫及其无公害治理.合肥:安徽科学技术出版社,2004:155-160.
Zhang H G, Tan J C.ChineseTeaTreeandPollution-FreePestManagement. Hefei: Anhui Science and Technology Publishing House, 2004:155-160. (in Chinese)
[2] 殷坤山,熊兴平.温度对茶尺蠖繁殖力的影响.中国茶叶,1994,16(2):18-19.
Yin K S, Xiong X P. The influence of temperature on the fecundity ofEctropisobliquehypulina.ChinaTea, 1994,16(2):18-19. (in Chinese)
[3] 侯建文.茶园蜘蛛种类的调查.茶叶,1980(1):41-44.
Hou J W. The investigation of spider species on tea garden.Tea, 1980(1):41-44. (in Chinese)
[4] 柯胜兵,周夏芝,毕守东,等.茶园3种害虫与蜘蛛天敌种群动态的关系.华中农业大学学报,2011,30(1):78-83.
Ke S B, Zhou X Z, Bi S D,etal. Relationships among three main pests and their natural enemies of spiders in tea garden.JournalofHuazhongAgriculturalUniversity, 2011,30(1):78-83. (in Chinese with English abstract)
[5] 王国昌,孙晓玲,董文霞,等.不同温度下鞍形花蟹蛛亚成蛛对茶尺蠖3日龄幼虫的捕食功能.茶叶科学,2010,30(3):173-176.
Wang G C, Sun X L, Dong W X,etal. Predatory response of penultimate-instarXysticusephippiatusSimon on 3-day-old larval of tea loopers under different temperatures.JournalofTeaScience, 2010,30(3):173-176. (in Chinese with English abstract)
[6] 胡萃,赵启泉,任翔,等.茶尺蠖绒茧蜂放蜂试验.昆虫天敌,1979(2):43-46.
Hu C, Zhao Q Q, Ren X,etal. The test of released honeybee onApantelessp.NaturalEnemiesofInsects, 1979(2):43-46. (in Chinese)
[7] 许宁,陈宗懋,游小清.引诱茶尺蠖天敌寄生蜂的茶树挥发物的分离与鉴定.昆虫学报,1999,42(2):126-131.
Xu N, Chen Z M, You X Q. Isolation and identification of tea plant volatiles attractive to tea geometrid parasitoids.ActaEntomologicaSinica, 1999,42(2):126-131. (in Chinese with English abstract)
[8] 张正群,孙晓玲,罗宗秀,等.14种植物精油对茶尺蠖行为的影响.茶叶科学,2014,34(5):489-496.
Zhang Z Q, Sun X L, Luo Z X,etal. Effect of 14 plant essential oils on the behavior ofEctropisoblique(Prout).JournalofTeaScience, 2014,34(5):489-496. (in Chinese with English abstract)
[9] 叶恭银,胡萃,朱俊庆,等.茶尺蠖核型多角体病毒对宿主种群的控制作用.植物保护学报,1994,21(3):231-237.
Ye G Y, Hu C, Zhu J Q,etal. The control of the geometrid nuclear polyhedrosis virus on host population.ActaPhytophylacicaSinica, 1994,21(3):231-237. (in Chinese with English abstract)
[10] 曹士先,程曦,蒋正中,等.基于cDNA-AFLP发掘茶树被茶尺蠖取食诱导的相关差异基因及其表达特征.中国农业科学,2013,46(19):4119-4130.
Cao S X, Chen X, Jiang Z Z,etal. Differential genes expression in tea plant (CameillaSinensisL.) induced byEctropisobliquefeeding based on cDNA-AFLP.ScientiaAgriculturaSinica, 2013,46(19):4119-4130. (in Chinese with English abstract)
[11] 任红楼,吕立哲,赵丰华,等.5种新型低毒农药对茶尺蠖的田间防控效果研究.中国茶叶,2013(4):28-29.
Ren H L, Lü L Z, Zhao F H,etal. Five kinds of new low-toxic pesticide on the field control effect of tea geometrid.ChinaTea, 2013(4):28-29. (in Chinese)
[12] 孙钦玉,张家侠,罗仲兴,等.皖南区茶尺蠖发生情况及防治技术措施.中国植保导刊,2014,34(2):38-40.
Sun Q Y, Zhang J X, Luo Z X,etal. The occurrence and control technology measures ofEctropisobliquein the South of Anhui.ChinaPlantProtection, 2014,34(2):38-40. (in Chinese)
[13] 傅伯杰,陈利顶,马克明,等.景观生态学原理及应用.北京:科学出版社,2001:216-217.
Fu B J, Chen L D, Ma K M,etal.ThePrincipleandApplicationofLandscapeEcology. Beijing: Science Press, 2001:216-217. (in Chinese)
[14] 邓聚龙.灰色系统理论教程.武汉:华中科技大学出版社,1990:33-84.
Deng J L.ACourseinGreySystemTheory. Wuhan: Huazhong Science and Engineering University Press, 1990:33-84. (in Chinese)
[15] 邹运鼎,王弘法.农林昆虫生态学.合肥:安徽科学技术出版社,1989:311-327.
Zou Y D, Wang H F.InsectEcologyofAgricultureandForestry. Heifei: Anhui Science and Technology Press, 1989:311-327. (in Chinese)
[16] 邹运鼎.害虫管理中的天敌评价理论与应用.北京:中国林业出版社,1997:27-90.
Zou Y D.TheoryandApplicationofEvaluatingNaturalEnemyinManagementofPests. Beijing: China Forestry Press, 1997:27-90. (in Chinese)
[17] David F N, Moore P G. Notes on contagious distribution in plant population.AnnalsofBotany, 1954,18:47-53.
[18] Arbous A G, Kerrich J E. Accident statistics and the concept of accident-proneness.Biometrics, 1951,7:340-432.
[19] Blackith R E. Nearest-neighbour distance measurements for the estimation of animal populations.Ecology, 1958,39:147-150.
[20] Iwao S. Application of them*-mmethod to the analysis of spatial patterns by changing the quadrat size.ResearchesonPopulationEcology, 1972,14:97-128.
[21] 程鸣珂,周坚.茶小绿叶蝉在茶园中空间分布型和抽样方法的研究.中国茶叶,1981(6):34-37.
Cheng M K, Zhou J. Studies on distribution and sampling ofEmpoascavitis(Gothe) in tea fields.ChinaTea, 1981(6):34-37. (in Chinese)
[22] 刘树生,汪信庚,吴晓晶,等.甘蓝和白菜上桃蚜种群的空间格局及其时序动态.昆虫学报,1996,39(2):158-165.
Liu S S, Wang X G, Wu X J,etal. Spatial pattern and its time series dynamics in the population ofMyzuspersicaeon twoBrassicaspp.ActaEntomologicalSinica, 1996,39(2):158-165. (in Chinese with English abstract)
[23] 石根生,李典谟.不同松林马尾松毛虫蛹及其寄生天敌群子的空间格局分析.生态学报,1997,17(4):386-392.
Shi G S, Li D M. Spatial distribution of masson-pine caterpillar pupae and its parasitic natural enemy guilds for the over wintering generation in the different pine forests.ActaEcologicaSinica, 1997,17(4):386-392. (in Chinese with English abstract)
[24] 邹运鼎,毕守东,周夏芝,等.桃一点叶蝉及草间小黑蛛空间格局的地学统计学研究.应用生态学报,2002,13(12):1645-1648.
Zou Y D, Bi S D, Zhou X Z,etal. Geostatistical analysis on spatial patterns ofErythroneurasudraandErigonidiumgraminicola.ChineseJournalofAppliedEcology, 2002,13(12):1645-1648. (in Chinese with English abstract)
[25] 邹运鼎,毕守东,王祥胜,等.麦长管蚜及蚜茧蜂空间格局的地学统计学研究.应用生态学报,2001,12(6):887-891.
Zou Y D, Bi S D, Wang X S,etal. Geostatistical analysis on spatial patterns ofMacrosiphumgranariumandAphidius.ChineseJournalofAppliedEcology, 2001,12(6):887-891. (in Chinese with English abstract)
[26] 耿继光,邹运鼎,毕守东,等.地理统计学表达的麦二叉蚜及蚜茧蜂空间格局特征.应用生态学报,2002,13(10):1307-1310.
Geng J G, Zou Y D, Bi S D,etal. Geostatistical analysis on spatial patterns ofSchizaphisgranariumandAphidiussp.ChineseJournalofAppliedEcology, 2002,13(10):1307-1310. (in Chinese with English abstract)
[27] 李磊,邹运鼎,毕守东,等.棉蚜和草间小黑蛛种群空间格局的地统计学研究.应用生态学报,2004,15(6):1043-1046.
Li L, Zou Y D, Bi S D,etal. Geostatistics analysis on spatial patterns ofAphisgossypiiandErigonidiumgraminicola.ChineseJournalofAppliedEcology, 2004,15(6):1043-1046. (in Chinese with English abstract)
[28] 丁程成,邹运鼎,毕守东,等.李园桃蚜和草间小黑蛛种群空间格局的地学统计学研究.应用生态学报,2005,16(7):1308-1312.
Ding C C, Zou Y D, Bi S D,etal. Geostatistics analysis on spatial patterns ofMyzuspersicaeandErigonidiumgraminicolain plum orchard.ChineseJournalofAppliedEcology, 2005,16(7):1308-1312. (in Chinese with English abstract)
[29] 邹运鼎,李昌根,周夏芝,等.葡萄跳叶甲和捕食性天敌草间小黑蛛的空间格局及其联系.植物保护学报,2007,34(3):241-246.
Zou Y D, Li C G, Zhou X Z,etal. Spatial construction and relationship betweenHalticachalybeaand its predatory natural enemy,Erigonidiumgraminicola.ActaPhytophylacicaSinica, 2007,34(3):241-246. (in Chinese with English abstract)
[30] 黄保宏,邹运鼎,毕守东,等.朝鲜球坚蚧及黑缘红瓢虫空间格局的地统计学研究.应用生态学报,2003,14(3):413-417.
Huang B H, Zou Y D, Bi S D,etal. Geostatistical analysis on spatial patterns ofDidesmococcuskoreanusBorchs andChilocorusrubidusHope.ChineseJournalofAppliedEcology, 2003,14(3):413-417. (in Chinese with English abstract)
[31] 赵静,赵鑫,王玉军,等.烟盲蝽及其天敌蜘蛛空间格局的地统计学分析.生态学报,2010,30(15):4196-4205.
Zhao J, Zhao X, Wang Y J,etal. Geostatistical analysis of spatial patterns ofNesidiocoristenuis(Reuter) (Hemiptera: Miridae) and its natural enemy spiders.ActaEcologicaSinica, 2010,30(15):4196-4205. (in Chinese with English abstract)
[32] 王瑞,翟保平,胡高,等.基于地统计学方法的稻田灰飞虱与蜘蛛时空动态分析.昆虫学报,2009,52(1):65-73.
Wang R, Zhai B P, Hu G,etal. Analysis of spatio-temporal dynamics ofLaodelphaxstriatellus(Fallen) (Homoptera: Delphacidae) and spiders in paddy fields based on geostatistics.ActaEntomologicaSinica, 2009,52(1):65-73. (in Chinese with English abstract)
[33] 邹运鼎,李磊,毕守东,等.石榴园棉蚜及其天敌之间的关系.应用生态学报,2004,15(12):2325-2329.
Zou Y D, Li L, Bi S D,etal. Relationships betweenAphisgossypiiand its natural enemies in megranate field.ChineseJournalofAppliedEcology, 2004,15(12):2325-2329. (in Chinese with English abstract)
[34] 杨林,郭骅,毕守东,等.合肥秋冬季茶园天敌对假眼小绿叶蝉和茶蚜的空间跟随关系.生态学报,2012,32(13):4215-4227.
Yang L, Guo H, Bi S D,etal. Spatial relationship amongEmpoascavitis(Gothe) andToxopteraaurantii(Boyer) and natural enemies in tea gardens of autumn-winter season in Hefei suburban.ActaEcologicaSinica, 2012,32(13):4215-4227. (in Chinese with English abstract)
Blocked quadrat variance analysis for spatial relation ofEctropisobliquehypulinalarva and spider natural enemies. Journal of Zhejiang University (Agric. & Life Sci.), 2015,41(2):133-146
Liu Feifei1, Ke Shengbing1, Wang Jianpan1, Bi Shoudong1*, Zou Yunding1, Zhou Xiazhi1, Dang Fenghua1, Xu Jinfeng2, Yu Kun1, Zhao Xuejuan1
(1.SchoolofScience,AnhuiAgriculturalUniversity,Hefei230036,China; 2.PlantProtectionStationofQianshanCityinAnhuiProvince,Qianshan246300,Anhui,China)
Ectropisobliquehypulina; spider natural enemy; spatial relationship; blocked quadrat variance analysis; grey correlation degree analysis
国家自然科学基金(30871444);安徽省教育厅重点项目(KJ2008A139).
联系方式:刘飞飞,E-mail:liufeifei805@163.com
2014-09-04;接受日期(Accepted):2015-01-21;网络出版日期(Published online):2015-03-20
Q 968.1
A
*通信作者(Corresponding author):毕守东,E-mail:bishoudong@163.com
URL:http://www.cnki.net/kcms/detail/33.1247.S.20150320.1907.003.html
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
茶叶科学(2021年4期)2021-08-12
浙江农业学报(2020年8期)2020-08-30
落叶果树(2020年2期)2020-04-20
——茶尺蠖和灰茶尺蠖的识别
中国茶叶(2019年11期)2019-12-03
现代园艺(2018年23期)2018-01-18
爱你(2017年13期)2017-11-15
中国新闻周刊(2015年31期)2015-08-28
