
鸡腺胃型传染性支气管炎病毒山东分离株S基因的克隆及序列分析
2015-06-11 02:21王玉东刘俊辉郑增忍王君玮王永玲王晓燕王树双牛钟相
动物医学进展 2015年6期
王玉东,刘俊辉,郑增忍,王君玮,王永玲,王晓燕,王树双,牛钟相
(1.中国动物卫生与流行病学中心,山东青岛266032;2.中国农业大学国际合作与交流处,北京100083;3.山东农业大学动物科技学院,山东泰安271018)
传染性支气管炎病毒(Infectious bronchitis virus,IBV)粒子主要包括3种主要结构蛋白,分别为纤突蛋白(S)、膜糖蛋白(M)、内部核衣壳蛋白(N),此外还有少量的第4种蛋白(小膜蛋白,E)[1-2]。病毒的纤突蛋白S蛋白由S1和S2两种糖蛋白组成,血凝抑制抗体和大部分病毒的中和抗体都是由S1诱导产生,具有免疫保护作用;据谢景伟等[3]报道,S1基因变异最大,是该病毒主要的抗原基因。因此本研究设计了一对S1和S2引物,对S蛋白基因核苷酸进行克隆及序列分析。1998 年王玉东等[4]首次报道了中国发生了腺胃型IB,并将青岛分离株命名为QXIBV。潘杰彦等[5]报道中国青岛腺胃型IBV 株(QXIBV 株又称SD/97/02株)的S1基因全序列及推导出的氨基酸序列被病毒基因库(Gen-Bank)所收录,收录号为AF193423。张小荣等[6]分离的2株河北分离IBV 株属于QXIBV 基因型。
Geerligs H J等[7]研制了 一 种QX 样IBV 活 疫苗,其病毒毒株与QXIBV 的同源性高达98.1%。近年来亚洲、欧洲和非洲都有大量IBV-QX 样病毒感染和发病的报道。Toffan A 等[8]报道非洲发生QX 样IBV。Abro S H 等[9]对IBV-QX 病毒 血清型按全基因序列分型,除了中国QXIBV 血清型外,欧洲多个国家分离到IBV-QX 样血清型,包括法国、德国、意大利、荷兰、波兰、俄罗斯、西班牙、瑞典和英国等都有相关报道,其分离毒株的S1 基因序列都与QXIBV 接近。Sigrist B 等[10]报道在瑞士发现了冠状病毒IBV-QX 型感染的病例。Ganapathy K 等[11]报道英国从商品肉鸡腺胃炎病例中分离到QX 样IBV。Zhang X R 等[12]报 道 了 来 源 于SPF 鸡IBV纤突蛋白在重组马立克病病毒的表达;Amin O G M等[13]报道了QX 样IBV 毒株在中东的传播,并对许多国家的病毒分离株进行了基因进化分析。Mork A K 等[14]报道了冠状 病毒IBV-QX 株和B1648病毒纤突蛋白特性的比较。QX 样IBV 感染病例呈现世界范围蔓延的趋势。
为研究腺胃型IBV 山东分离株QX 和DY 及FC的病毒S蛋白的核苷酸特征,确认其与肾型IBV和呼吸型IBV 的同源性。采用分子生物学技术(RT-PCR 试验方法),对多株IBV 毒株进行了纤突蛋白S基因的扩增,并对QX 和DY 株的S基因扩增产物核苷酸序列进行分析,表明腺胃型IBV 是不同于呼吸型IBV 和肾型IBV 的一种具有明显腺胃嗜性的IBV 变异株。
1 材料与方法
1.1 材料
1.1.1 病毒毒株 山东省腺胃型IBV 典型分离株青 岛 株(QXIBV)、东 营 株(DYIBV)、肥 城 株(FCIBV)、肾型IB分离株(HD),均由中国动物卫生与流行病学中心分离鉴定、保存。IBV H52标准株、H120株、M41标准株、肾型IBV-T 株,均购自中国兽医药品监察所。
1.1.2 主要仪器及试剂 台式高速(12 000r/min)离心机为德国Eppendorf公司产品;DNA 扩增仪为美国MJ Research 公司产品;稳压稳流电泳仪和水平电泳槽和凝胶成像分析系统为美国伯乐公司产品;振荡器购自江苏常州国华公司;恒温水浴锅为天津泰斯特仪器公司产品;冰箱为青岛海尔公司产品。
NaCl,MgCl2,NaH2PO4,Na2HPO4,KH2PO4,K2HPO4,KCl,NaOH,HCl,三 羟 甲 基 氨 基 甲 烷(Tris),EDTA,异丙醇,无水乙醇,氯仿,总RNA 抽提剂(Trizol),琼脂糖,溴化乙锭,硼酸,分别为山东赛恩斯科技公司和青岛爱普科生物工程公司产品。
Taq DNA 聚合酶为上海生工生物工程技术服务有限公司产品;M-MLV 反转录酶、RNA 酶抑制剂、脱氧核苷三磷酸(dNTP)、DEPC(焦碳酸二乙酯)为Promega公司产品;DNA Marker DL 2 000购自宝生物工程(大连)有限公司。
1.1.3 引物 参照GenBank注册的IBV S通用基因序列,自行设计了一对跨幅约0.9kb的引物,该区段包含了S1基因C 末端的400bp和S2基因N末端的500bp。该基因片段具有良好的保守性和遗传特异性,是IBV S基因研究者选择较多的合成引物。引物预期扩增片段大小为750bp~950bp,引物的使用浓度为10pmol/μL。
引物序列如下:S1:5′CAAGGTTTTATTACTAATGT 3′;S2:5′AGCAAACCATTATATTCACGA 3′。以上引物由上海博亚生物技术有限公司合成。
1.2 方法
1.2.1 RNA 的提取 分别取典型腺胃型IBV 分离株(QX 株、DY 株、FC 株)的 病 毒 尿 囊 液 各100 μL,放入1.5mL的离心管中,加500μL Trizol,上下颠倒5次,混匀,室温放置15 min。加入100μL氯仿,涡旋混匀,室温放置8 min,期间混匀2 次。12 000r/min离心15min,取上清200μL加200μL异丙醇混匀,在-20℃放置过夜。12 000r/min离心15min,缓慢倒掉上清,沿管壁缓慢加750mL/L乙醇0.5mL进行洗涤一次,缓慢倒掉750mL/L乙醇,倒扣于吸水纸上,置超净工作台晾干,加入20 μL DEPC处理的无菌双蒸水溶解核酸,置-20℃保存备用。
1.2.2 反转录(RT) 在干净的1.5mL 的离心管中依次加入以下试剂:5×M-MLV RT buffer 4μL,40 U RNA 酶 抑 制 剂1 μL,2.5 mmol/L dNTP 3μL,10pmol/μL 混合引物(S1+S2)1μL,RNA 提取样品5μL,200 U M-MLV 反转录酶1 μL,双蒸水5μL,总体积为20μL。混匀后置42℃30min,37℃30min,95℃3min,冷却至4℃。
1.2.3 目的基因的PCR 扩增 在0.2mL 的反应管中依次加入以下试剂:10×PCR buffer 3μL,25 mmol/L MgCl22μL,2.5 mmol/L dNTP 3μL,10 pmol/μL的混合引物(S1+S2)1μL,反转录产物5μL,5UTaq DNA 聚合酶0.5μL,双蒸水15.5 μL,总体积30μL。PCR 反应条件为:94℃5min;35个循环,每个循环包括94℃1 min,55℃1 min,72℃2min;最后72℃延伸10min。
1.2.4 PCR 产 物 检 测 获 得 的PCR 产 物 于10 g/L琼脂糖凝胶(已含有0.5μg/mL 的溴化乙锭),在电压100 V~120 V 条件下,电泳30 min~60 min。当电泳结束后,取出凝胶反应板,放置于凝胶成像分析系统中进行观察,并照相记录。
1.2.5 S基因序列测定及分析 将PCR 产物纯化后,用引物S1 进行测序。使用DNAstar序列分析软件,对克隆的S基因核苷酸同源性进行比较。为探讨不同分析方法的优势和特点,消除因不同方法产生的误差,保证分析结果的准确性和科学性,选择国际通用的Clustal V method、Jotun Hein method和Clustal W method三种方法,分别对2株腺胃型QX 株、DY 株、1株 肾 型IB HD 株 和 标 准M41株 的S基因核苷酸序列进行比对,对其同源性进行分析。由上海博亚生物技术有限公司协助完成。
1.2.6 S基因序列进化关系分析 使用DNAStar对腺胃型IBV(QX 株、DY 株)与标准株(M41株)和肾型IBV(HD 株)同源性进行进化关系分析。由上海博亚生物技术有限公司协助完成。
2 结果
2.1 RT-PCR 扩增产物检测结果
用自行设计的一对S基因引物,通过RT-PCR扩增产物进行检测(图1、图2)。山东分离腺胃型QX 株、DY 株、肾型HD 株均与标准株(H52、H120、M41)产物的电泳条带相似,大小约为900bp(在789bp~857bp之间,标准株M41株为857bp),表明设计引物对IBV 具有良好的特异性。其中QX 株和DY 株与标准呼吸型IB M41、H52、H120、肾型HD 株出现的条带明显;FC 株产物有较大差异(仅为500bp),未对其第2次基因扩增,也未分析比对;肾型IB-T 株未形成明显的产物条带。对M41标准株、QX 株、DY 株、肾 型IB 分 离 株HD 株 四 株 病 毒进行第2次RT-PCR,其扩增产物的电泳条带与第一次的结果相同(约为900pb)。
2.2 S基因序列测定结果
将M41株、HD 株、QX 株、DY 株4 株IBV 病毒的PCR 扩增产物进行测序,测序结果见图3~图6。4株病毒S 基因均为IBV 的部分S 基因,其中M41株病毒S基因产物为857bp,S基因序列为多数基因序列;腺胃型QX 株为837bp,DY 株为852 bp,肾型HD 株为789bp。各病毒被扩增出的基因片段均含有编码部分S蛋白的阅读框架(ORF)。
2.3 S基因核苷酸同源性分析
2.3.1 Clustal V method、Jotun Hein method 和Clustal W method三种方法分析图 对2株腺胃型分 离株(QX 株、DY 株),1株 肾 型IB HD 株 和 标 准株M41株的S基因核苷酸序列进行比对,对其同源性进行了分析(图7~图9)。对QX 株、DY 株、HD株和M41标准株进行S基因核苷酸序列分析及其同源性比较结果发现,三种方法之间略有差异,核苷酸分析同源性数值Jotun Hein 法较高,Clustal W法次之,Clustal V 法最低。

图1 9种IBV 的S基因扩增产物条带Fig.1 The amplification products of S genes of 9IBV strains
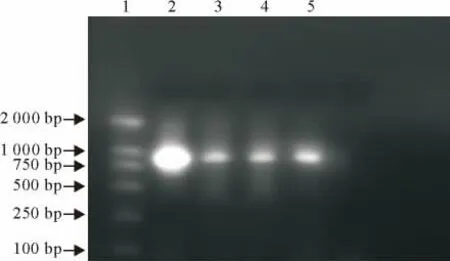
图2 4种IBV 的S基因扩增产物条带Fig.2 The amplification products of S genes of 4IBV strains

图3 病毒标准株(M41株)的S基因测序结果Fig.3 S gene sequencing of reference IBV M41strain

图4 肾型IB分离株(HD 株)的S基因测序结果Fig.4 S gene sequencing of nephric type IBV HD strain

图5 腺胃型IB病毒青岛株(QX 株)的S基因测序结果Fig.5 S gene sequencing of proventriculus type IBV Qingdao QX strain

图6 腺胃型IBV 东营株(DY 株)的S基因测序结果Fig.6 S gene sequencing of proventriculus type IBV DY strain
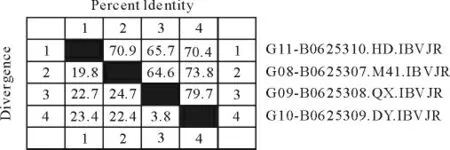
图7 用Clustal V 方法分析结果Fig.7 Analysis result by clustal V method
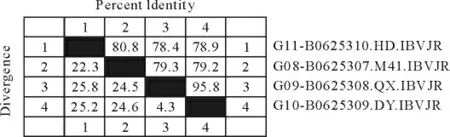
图8 用Jotun Hein 方法分析结果Fig.8 Analysis result by Jotun Hein method
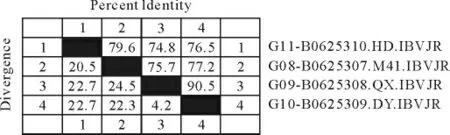
图9 用Clustal W 方法分析结果Fig.9 Analysis result by Clustal W method
2.3.2 S基因核酸的同源性分析 按Clustal V 法QX 株和标准M41株的同源性达到64.6%;DY 株与标准M41株的同源性为73.8%;肾型IB HD 株与M41株的同源性为70.9%;QX 株与HD 株的同源性为65.7%;DY 株与HD 株的同源性为70.4%;QX 株与DY 株的同源性为79.7%。QX 株与DY株的差异性为4.3%。QX 株和DY 株与M41株的差异性分别为24.7%和22.4%。QX 株和DY 株与HD 株的差异性为22.7%和23.4%。HD 株与M41株的差异性为19.8%。
按Jotun Hein法 同源性比较的数值均略高于Clustal V 法。QX 株和标准M41株的同源性为79.3%;DY 株与标准M41株的同源性为79.2%;HD 株与M41株的同源性为80.8%;QX 株与HD株的同源性为78.4%;DY 株与HD 株的同源性为78.9%;QX 株 与DY 株 之 间 的 同 源 性 为95.8%,QX株与DY 株的差异性为3.8%;QX 株和DY 株与M41株的差异性分别为24.5%和24.6%;QX 株和DY 株与HD 株的差异性分别为25.8%和25.2%;HD 株与M41株的差异性为22.3%。QX株和DY 株同源性极高,为95.8%。
按Clustal W method 法 QX 株和标准M41株的同源性为75.7%;DY 株与标准M41株的同源性为77.2%;HD 株与M41株的同源性为79.6%;QX 株与HD 株的 同源性为74.8%;DY 株与HD株的同源性为76.5%;QX 株与DY 株的同源性为90.5%,QX 株与DY 株的差异性为4.2%;QX 株和DY 株与M41 株的差异性分别为24.5%和22.3%。QX 株和DY 株与HD 株的差异性均为22.7%,HD 株与M41株的差异性为20.5%。
归纳三种分析方法结果,QX 株和DY 株的S基因的核苷酸同源性最高达到95.8%,最小差异性为3.8%(张小荣[4]分离的QXIBV 基因型2分离株的同源性仅为77.7%)。QX 株和DY 株病毒与标准病毒M41株的同源性最高达79.3%和79.2%,最小差异性为24.5%和22.3%。QX 株和DY 株与HD 株的同源性最高达78.4%和78.9%,最小差异性均为22.7%。肾型HD 株与M41株的同源性最高达到80.8%。腺胃型QX 株和DY 株分别与呼吸型M41株及肾型HD 株的同源性差异不明显。结果表明,腺胃型IBV 与呼吸型IBV 和肾型IBV 是不同组织嗜性的传染性支气管炎病毒变异株,与呼吸型和肾型IBV 具有同等重要地位的进化阶段。
2.4 S基因系列进化关系分析
对腺胃型QX 株、DY 株和肾型HD 株与标准株M41株进化关系分析见图10。QX和DY 株、M41和肾型HD株各为一个分支;4株病毒分成2个群,Ⅰ群包含M41 株和肾型HD 株两株病毒,遗传距离为19.8%~22.3%,同源性在70.9%~80.8%之间。Ⅱ群包含了QX 株和DY 株,遗传距离在3.8%~4.3%之间,同源性在79.7%以上,QX 株和DY 株之间同源性最高达95.8%。腺胃型QX 和DY 株的遗传距离很小,按Jotun Hein 法分析仅为3.8%。通过进化分析说明,腺胃型IBV 与肾型IBV 和呼吸型IBV 是不同的IBV 病毒的变异株。
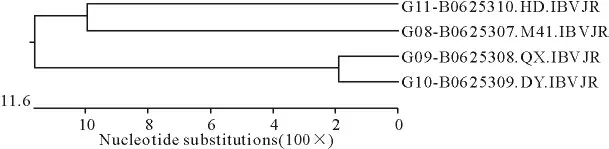
图10 4株病毒S基因系列进化关系分析图Fig.10 The phylogenetic analysis of S genes of 4viral strains
3 讨论
3.1 引物设计和基因序列分析
由于IBV 变异性强,引物的设计必须在基因的高度保守区内进行,以便能获得一对能广泛应用于所有IBV 的通用引物。但为了能对分离的病毒进行分类,又必须选择能代表病毒基因变异的区域进行研究,S纤突蛋白基因被当作IBV 分型的主要研究对象。
在S纤突蛋白中S2蛋白相对保守,但同样含有两个抗原决定簇。S1 基因的3/4 处属于多变区[15]。参考基因库IBV S基因序列,自行设计了一对引物,对9株IBV 进行了RT-PCR,其中7株均形成了特异的目的基因片段,获得了理想的扩增产物。QX 株、DY 株、肾 型HD 株 及M41 株 四 株 病 毒 的RT-PCR 扩增产物大小范围为787bp~857bp,对其进行基因序列测定及同源性分析,表明腺胃型IBV 分离株QX 株和DY 株是一种不同于呼吸型和肾型IBV 的组织嗜性的新的病毒变异株。QX 和DY 株两株病毒的同源性高达95.8%。QX 和DY株与M41标准株及肾型HD 株的同源性较低。因FC株的PCR 扩增产物与标准株有较大差异,未进行DNA 序列分析和同源性比较。
3.2 腺胃型IBV 分子流行病学分析
从临床上表现典型症状的病鸡分离到2株病毒QX 株和DY 株均有嗜腺胃特性,说明我国流行的IBV 与疫苗毒株和国际上许多IBV 毒株发生了明显的变异。按进化关系分析QX 株和DY 株为一群,这两株腺胃型IBV S 基因序列有高度的同源性。
腺胃型IBV 分离株QX 株和DY 株都与国内流行的肾型IB HD 株和标准毒株M41株具有较远的亲缘关系。近年来嗜腺胃型IBV 的研究也不断有所发展。王玉东等[16]报道,腺胃型IBV 是与呼吸型和肾型IBV 不同血清型的变异株。王玉东等[17]进行的山东分离株(QX 株、DY 株、FC株)SPF鸡致病回归试验获得了成功,外源病毒的检测均为阴性。对山东3个分离株进行了病毒病理学和超微结构的研究,表明本病的病原为冠状病毒属的嗜腺胃型IBV 病毒[18]。
Beato M S等[19]报道了IBV 中国分离株QXIBV 在意大利传播流行的证据。Pohuang T 等[20]报道中将泰国分离的IBV 进行S1基因的序列分析。近年来该病呈世界性流行,Abro S H 等[9]的报道表明欧洲发病病例最多。
3.3 对引起腺胃炎病变的腺胃型IBV 病原学研究和防疫的思考
1995年-1996年发生于江苏省和山东省的腺胃型IB,国外最初未见此类疾病的报道,是一种新的IBV 变异株。Saif Y M 主编的禽病学第11版和第12版均引用了王玉东的报道[1-2]。10余年来,中国各地呈现蔓延趋势,使用中国青岛分离株(QXIBV 株)和山东分离株病毒制备的油乳剂灭活苗,在中国大面积范围内使用表明,免疫效果好,保护率高[4,16]。国外近年来也陆续有疫苗研制的报道,Terregino C 等[21]报 道,意 大 利 分 离 株IBV-QX 株对SPF鸡和商品肉鸡的致病性,在蛋鸡和肉鸡场1日龄和14日龄时免疫Ma5和4/91变异株疫苗,对降低发生IBV-QX 感染时引起的经济损失具有帮助作用。Geerligs H J等[7]报道的一种与QXIBV 基因同源性高达98.1%的IB-QX 类病毒疫苗,主要用于鸡肾型IB的免疫,目前未进行大面积推广,免疫效果如何,尚未可知。
近年来国内对本病的病原学研究一直争论不休,包括传染性支气管炎病毒、网状内皮组织增殖病病毒、新城疫病毒、细小病毒、呼肠病毒等病原在有关腺胃炎的研究中都有分离到。我们认为该病的病原为一种IBV 变异株-即腺胃型IBV 变异株。IBV的多变异特性对IBV 引起的传染病的诊断和防疫带来了严重的困难,所以如何快速、及时地对所分离的IBV 进行分析和研究,对IBV 的防控十分重要。对该病病原的分子流行病学分析研究,仍然是未来一段时间研究的重要内容。
[1] Saif Y M.禽病学[M].11版.苏敬良,高 福,索 勋,主译.北京:中国农业出版社,2005:108-130.
[2] Saif Y M.禽病学[M].12版.苏敬良,高 福,索 勋,主译.北京:中国农业出版社,2012:129-151.
[3] 谢景伟,陈 峰,谢青梅,等.鸡传染性支气管炎病毒分子流行病学研究进展[J].动物医学进展,2008,29(8):75-78.
[4] 王玉东,王永玲,张子春,等.鸡腺胃型传染性支气管病毒(QXIBV)的分离和鉴定[J].中国动物检疫,1998,15(1):1-3.
[5] 潘杰彦,陈德胜,戴亚斌,等.IBV 青岛腺胃分离株(SD/97/02)S1蛋白基因的序列测定和分析[J].中国病毒学,2001,16(4):377-381.
[6] 张小荣,程靖华,陈启稳,等.鸡腺胃分离的传染性支气管炎病毒基因型与致病性研究[J].扬州大学学报:农业与生命科学版,2012,33(4):6-9.
[7] Geerligs H J,Boelm G J,Meinders C A,et al.Efficacy and safety of an attenuated live QX-like infectious bronchitis virus strain as a vaccine for chickens[J].Avian Pathol,2011,40(1):93-102.
[8] Toffan A,Monne I,Terregino C,et al.QX-like infectious bronchitis virus in Africa[J].Vet Rec,2011,169(22):589.
[9] Abro S H,Rrenstrom L H,Ullman K,et al.Characterization and analysis of the full-length genome of a strain of the european qx-like genotype of infectious bronchitis virus[J].Arch Virol,2012,157(6):1211-1215.
[10] Sigrist B,Tobler K,Schybli M,et al.Detection of avian coronavirus infectious bronchitis virus type qx infection in switzerland[J].J Vet Diagn Invest,2012,24(6):1180-1183.
[11] Ganapathy K,Wilkins M,Forrester A,et al.QX-like infectious bronchitis virus isolated from proventriculitis in commercial broilers in England[J].Vet Rec,2012,doi:10.1136/vr.101005:1-2.
[12] Zhang X R,Wu Y T,Huang Y Z,et al.Protection conferred by a recombinant Marek's disease virus that expresses the spike protein from infectious bronchitis virus in specific pathogen-free chicken[J].Virol J,2012,9:85.
[13] Amin O G M,Valastro V,Salviato A,et al.Circulation of QX-like infectious bronchitis virus in the Middle East[J].Vet Rec,2012,171:530.
[14] Mork A K,Hesse M.Differences in the tissue tropism to chicken oviduct epithelial cells between avian coronavirus IBV strains QX and B1648are not related to the sialic acid binding properties of their spike proteins[J].Vet Res,2014,45(1):67.
[15] 于圣青,译.与传染性支气管炎病毒血清型有关的S1亚基氨基酸差异的定位[J].中国家禽,1994(4):30-31.
[16] 王玉东,王永玲,范根成,等.鸡腺胃型传染性支气管炎(综述)[J].畜牧兽医学报,2001,32(1):64-72.
[17] 王玉东,王永玲,张国中,等.三株腺胃型传染性支气管炎病毒分离毒株对鸡的致病性试验[J].中国兽医科学,2015,45(2):195-201.
[18] 王玉东,王永玲,王君玮,等.山东分离株鸡腺胃型传染性支气管炎病毒病理组织学和电镜观察[J].中国兽医杂志,2015年,待发表.
[19] Beato M S,De Battisti C,Terregino C,et al.Evidence of circulation of a Chinese strain of infectious bronchitis virus(QXIBV)in Italy[J].Vet Rec:J Bri Vet Asso,2005,156(22):720.
[20] Pohuang T,Chansiripornchai N,Tawatsin A,et al.Sequence analysis of S1genes of infectious bronchitis virus isolated in Thailand during 2008-2009:identification of natural recombination in the field isolates[J].Virus Genes,2011,43(2):254-260.
[21] Terregino C,Toffan A.Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens,and evaluation of protection induced by a vaccination programme based on the Ma5and 4/91serotypes[J].Avian Pathol,2008,37(5):487-493.
猜你喜欢
中国畜禽种业(2021年1期)2021-12-02
现代畜牧科技(2021年8期)2021-10-13
中国动物保健(2021年9期)2021-09-22
粉末冶金技术(2021年3期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
天然产物研究与开发(2019年10期)2019-11-05
兽医导刊(2019年10期)2019-02-12
兽医导刊(2018年21期)2018-03-18
中国民族医药杂志(2016年2期)2016-05-14
养殖与饲料(2015年12期)2015-02-10
