
不同食用木薯品种木薯棒孢霉叶斑病空间分布型研究初报*
2015-05-26 01:21段春芳娄予强周迎春熊贤坤易怀峰李月仙刘倩严炜沈绍斌宋记明张林辉刘光华
西部林业科学 2015年5期
段春芳,娄予强,周迎春,熊贤坤,易怀峰,李月仙,刘倩,严炜,沈绍斌,宋记明,张林辉,刘光华
(云南省农业科学院热带亚热带经济作物研究所,云南 保山 678000)
木薯 (Manihot esculenta Crantz),为大戟科(Euphorbiaceae)木薯属灌木状多年生作物,其块根肉质,富含淀粉,是木薯、马铃薯 (Solanum tuberosum)、甘薯 (Ipomoea batatas Lam)世界三大薯类之一。木薯起源于热带美洲,于19世纪20年代引入中国,主要分布在海南、广东、广西、云南和福建等省,而湖南、四川和贵州也有零散分布[1],种植面积一直保持在 40×104hm2以上[2]。目前,鲜薯主要被用作加工淀粉和酒精的生产原料,较少直接食用[3]。随着人们生活水平的提高,吃粗粮和杂粮已成为一种健康的生活方式,加上近几年来不断有优良食用木薯品种的选育和推广,木薯越来越受人们的关注和喜爱。然而,木薯在生产中易受病害的侵染而影响木薯产量[4]。木薯棒孢霉叶斑病是由山扁豆生棒孢 (Corynespora cassiicola)引起的病害[5],在海南的儋州、白沙,广西的武鸣、北海以及广东的茂名、湛江均有发生[6~7],棒孢霉叶斑病在田间常表现为:发病初期叶片上出现黄褐色圆形小点,其周围具有明显的黄色晕圈;病斑扩大后成近圆形病斑,病斑深褐色,中心略呈白色纸质状,边缘黑褐色,周围具有很明显的黄色晕圈。近年来,棒孢霉叶斑病在云南保山木薯种植区普遍发生,从田间观察到,不同木薯品种上棒孢霉叶斑病发病有差异。
空间分布型是病虫在空间相对静止时的散布状况,它揭示了病虫个体某一时刻的行为习性、环境因子对叠加影响,以及种群选择环境的内禀特性和空间结构异质性程度。在估计病虫密度的抽样调查、确定试验数据的统计分析方法及考察受害形成过程和决定必需防治密度时,空间分布型的确定是必不可少的[8]。目前,国内关于木薯棒孢霉叶斑病只是在病原鉴定上有所报道[7],关于空间分布方面的研究未见报道。该病害在云南木薯种植区从无到有,从轻到重,从该调查地观察得到,木薯棒孢霉叶斑病从木薯种植3个月后开始发病,比其他叶部病害发病时间早,病害流行受阴雨天气影响较大,为了解木薯棒孢霉叶斑病的发病规律和空间分布特征,2014年10月10日和11月24日两次对该病害在不同食用木薯品种上的空间分布型进行了初步研究,以期为该病害监测预报及综合防治工作提供依据。
1 材料与方法
1.1 样地概况
调查样地设在云南省农业科学院热带亚热带经济作物研究所木薯科技示范园,位于云南省保山市隆阳区潞江坝,东经 98°52′53″,北纬 24°58′52″,海拔为704 m,年平均温为21.5℃,年降雨量755 mm,年蒸发量2 111.5 mm,绝对最高气温40.4℃,≥10℃活动积温7 800℃,年日照数2 325.75 h,相对湿度为70%,属于南亚热带向热带过渡的干热河谷气候,土壤为冲积母质发育的沙壤土,有机质含量 1.3%,pH 值 6.5~7.0。木薯面积为 8 hm2,调查木薯品种有 SC9、SC201、GR891和 E1424,以及对照品种SC124(CK),所调查木薯均为1年生木薯。2014年9月下旬,笔者在田间病虫害调查时发现,木薯棒孢霉叶斑病对木薯危害较其他年份严重,在品种SC9上的发病率高达51.57%,平均病斑数为2.44个/叶,因此,本项目重点对几个食用木薯品种做了病害调查研究,为病害监测提供基础性数据。
1.2 方法
1.2.1 调查时间
在木薯发病中期和发病后期,选择2014年10月10日和2014年11月24日进行调查。
1.2.2 调查方法
参考玉米 (Zea mays)[9~10]和梨树 (Pyrus sorotina)[11]等作物的调查方法:在试验地内随机选取5块样地,采用随机抽样调查法,在每块样地随机选取6株木薯,每株的上部和中部各取5个叶片,每株共取10个叶片,总共300片木薯叶,记录每个叶片的病斑总数,采用空间分布型指标确定病害的空间分布型。
1.3 数据分析
用Excel 2003和DPS v 7.0处理田间调查的数据。
1.4 空间格局的测定方法
1.4.1 聚集度指标[12~15]
(1)扩散系数 (C) C=S2/x,式中,S2为样本方差;x为样本均数。当C>1时为聚集分布;C<1时为均匀分布;C=1时为随机分布。
(2)Cassie 指标 (Ca) Ca=(S2-x)/x2,当Ca<0时为均匀分布;当Ca>0时为聚集分布;Ca=0时为随机分布。
(3)David-Moore 丛生指标 (I) I=S2/x-1,当I>0时为聚集分布;I<0时为均匀分布;I=0时为随机分布。
(4)平均拥挤度 (M*) M*=M+S2/M-1,式中,M*为样本中个体平均值;M为种群的平均密度;S2为方差。
(5)聚集性指数 (M*/M) M*/M=S2-M/M2+1,当M*/M<1时,个体在种群中为均匀分布;当M*/M=1时,个体在种群中为随机分布;当M*/M>1时,个体在种群中为聚集分布,其值越大,个体间聚集度越大。
(6)负二项参数 (k) k值愈小,聚集度愈大;若k值趋于无穷大 (一般为8以上),则接近随机分布。
1.4.2 兰星平[16]的 Lα-m 回归模型
Lα-m回归模型 Lα=θ+ηm,式中,m为样本平均值,θ、η为引入的参数。当 θ>0,η≥1时,为聚集分布;θ=0,η>1时,为聚集分布;θ=0,η=1时,为随机分布;θ=0,η<1时,为均匀分布;θ<0,η≤1时,为均匀分布;θ>0,η<1或 θ<0,η>1时,θ/m+η<1为均匀分布,θ/m+η=1,为随机分布,θ/m+η>1为聚集分布。
1.4.3 张连翔[17]Z-V 模型
Z-V模型 Z=A+BV,式中,v为样本方差。A=0,B<1,A<0,B≤1,A>0,B<1,V>Vα 〔Vα=A/(1-B)〕,A<0,B>1,V<Vα 时,为均匀分布,A=0,B>1,A>0,B≥1,A>0,B<1,V<Vα,A<0,B>1,V>Vα 时,为聚集分布;A=0,B=1,A>0,B<1,V=Vα,A<0,B>1,V=Vα 时,为随机分布。
1.5 数据处理
采用方差分析比较不同食用木薯品种上棒孢霉叶斑病发病情况。
2 结果与分析
2.1 空间分布型
不同食用木薯品种,11月24日调查的木薯棒孢霉叶斑病田间聚集度各项指标见表1。

表1 不同食用木薯品种上棒孢霉叶斑病的集聚度指标Tab.1 Spatial distribution indices of Corynespora leaf spot disaese of cassava in different edible cassava species
9种指标判定木薯棒孢霉叶斑病的空间分布型为聚集分布,其中SC124(CK)的负二项参数k>8,说明木薯棒孢霉叶斑病在SC124(CK)上呈聚集分布,但接近于随机分布;Cassie指标Ca值均大于0,但平均病斑数高的SC124(CK)和GR891以及平均病斑数最少的E1424较其他品种接近0,这说明木薯棒孢霉叶斑病随着病斑密度较高或者较低时趋向于随机分布;在SC124(CK)、GR891和E1424上聚集程度不高,接近于随机分布,数值上的表现同于Cassie指标Ca值。木薯棒孢霉叶斑病在田间各品种上分布总体呈聚集分布状态,其原因主要由病害的发生规律和外界环境因子共同作用。木薯棒孢霉叶斑病首先在病株上形成初侵染源,病原菌的孢子随着风雨等传播行进扩散,从而形成病株为发病中心的聚集块,同时,由于不同品种抗病力不一致,导致各个木薯品种之间聚集度存在着差异性。
2.2 线性回归模型
用Lα-m回归模型和Z-V模型得到的不同木薯食用品种上棒孢霉叶斑病回归方程见表2。从采用的4种回归模型结果来看,Lα-m回归模型和Z-V模型得到的结果一致,与聚集度指标测定结果相一致。从Lα-m回归模型相关系数r大于Z-V模型相关系数r值,说明木薯棒孢霉叶斑病的空间分布型以Lα-m回归模型为佳。
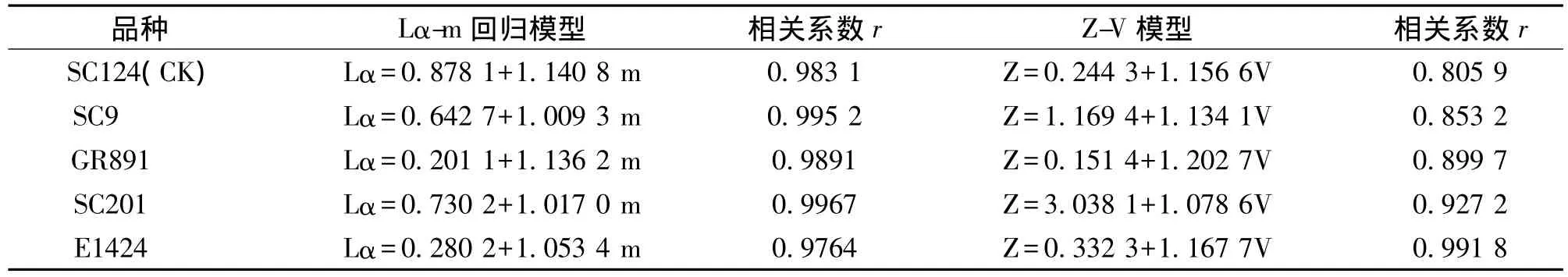
表2 不同食用木薯品种棒孢霉叶斑病聚集度指标的回归模型Tab.2 The regressive model for aggregation indexes of Corynespora leaf spot disaese of cassava in different edible cassava species
2.3 不同食用木薯品种的棒孢霉叶斑病发病情况
不同食用木薯品种的棒孢霉叶斑病病斑的平均数如图1所示,4个木薯品种病斑数均比对照SC124(CK)低,其中新选育的品种E1424病斑数最少,且第二次调查的株病斑数大于第一次。从图1还可知,木薯棒孢霉叶斑病在发病初期,4个木薯品种之间平均病斑数差异不显著,与SC124(CK)差异显著。到11月下旬,5个木薯品种之间平均病斑数差异显著。该研究结果表明,SC124(CK)、SC201、SC9和GR891发病较E1424严重。除了木薯品种E1424以外,其余木薯品种已选育和种植多年,导致品种抗病力严重退化,因此,选育出高抗、高产、优质的木薯品种将是今后中国木薯产业发展的一项重点工作。

图1 棒孢霉叶斑病在不同食用木薯品种上的平均病斑数Fig.1 The biomass of Acrocarpus fraxinifolius plantation
3 结论与讨论
有研究报道表明,病害的空间分布随着田间发病率的轻重而有所不同:百日草 (Zinnia elegans Jacp.)[18]病害的空间分布型在每叶病斑数均值小于70个时为聚集分布,随着每叶病斑数均值增大,病害的空间分布型逐渐变为均匀分布;油菜菌核病(Sclerotinia sclerotiorum)[8]在发病轻的条件下,总体上呈聚集分布,但聚集程度不高,少量田块呈随机、均匀分布。轻发生时油菜菌核病的空间分布型趋向随机和均匀分布;玉米褐斑病 (Physoderma maydis Miyabe)[10]在发病率相对低时,空间分布型基本为随机分布,当发病率相对较高时,空间分布型基本为均匀分布。木薯棒孢霉叶斑病空间分布同样存在着类似的结论。在棒孢霉叶斑病田间发病较高的年份是否又出现从聚集分布到随机分布或均匀分布,还有待于进一步调查研究。
针对病虫害的空间分布型,应用最广的是Iwao 的 M*-m 回归分析和 Taylor幂法则[19~20],在分析过程中,常常会出现m*与m不为线性关系,以及v与m不为幂函数关系的情况,为了克服模型出现这样的不足之处[21~22],本研究采用的 Lα-m模型和Z-V模型来分析木薯棒孢霉叶斑病空间分布型。Lα-m模型和Z-V模型计算简便,易于掌握应用,而且模型拟合的效果较好,本文作者曾用M*-m 回归分析、Taylor幂法则、Lα-m 模型和 Z-V模型分析木薯烟粉虱 (Bemisia tabaci)成虫的空间分布,结果 Lα-m模型和 Z-V模型拟合效果好于M*-m回归分析、Taylor幂法则 (待发表)。在调查方法上,参考了梨树、玉米、油菜 (Brassica campestris)和百日草等作物的空间分布研究报道,报道中统计斑病数时,上述作物玉米病害病斑数采用了以株为单位,梨树锈病采用了以叶为单位。在木薯棒孢霉叶斑病调查期间,各个木薯品种之间株高和分枝数差异大,结合木薯的实际情况,调查时采用每株上部和中部共取10个叶片,以每片木薯叶片上的病斑平均数来分析病害空间分布型。本研究所参考的作物种植密度与木薯有所不同,今后,在木薯病害空间分布型的调查方法上,还有待于作进一步研究和改进。
[1]刘光华,刘倩,张林辉,等.云南木薯高效栽培技术[M].昆明:云南科技出版社,2010.
[2]李开绵,林雄,黄洁.国内外木薯科研发展概况[J].热带农业科学,2001,89(1):56-60.
[3]单荣芝,黄洁.我国食用木薯的历史、现状与发展对策[J].农业科技管理,2013,32(5):73-76.
[4]李开绵,黄贵修.木薯主要病虫害[M].北京:中国农业科学技术出版社,2008.
[5]Liu X.B,Shi T,Li C.P,et al.First Report of Corynespora cassiicola causing leaf spot of cassava in China[J].plant disease,2010,94(7):916.
[6]李超萍,时涛,刘先宝,等.国内木薯病害普查及细菌性萎蔫病安全性评估[J].热带作物学报,2011,32(1):116-121.
[7]裴月令,时涛,蔡吉苗,等.木薯棒孢霉叶斑病病原鉴定及其生物学特性测定[J].热带作物学报,2011,32(4):728-733.
[8]朱金良,陈跃,王光华,等.油菜菌核病轻发生条件下的空间分布格局与抽样技术[J].浙江农业学报,2012,24(3):464-468.
[9]王艳红,王晓梅,张建春,等.长春地区玉米大斑病、灰斑病空间分布型研究[J].玉米科学,2009,17(5):148-151.
[10]段显德,王晓梅,马腾达,等.玉米褐斑病空间分布型研究[J].辽东学院学报,2011,18(4):290-292.
[11]郑义,段显德,崔欣,等.梨树锈病病斑在叶片上的空间分布型及抽样技术[J].东北林业大学学报,2013,41(12):80-83.
[12]徐汝梅.昆虫种群生态学[M].北京:北京师范大学出版社,1987.
[13]陈小飞,熊仁次,王兰,等.南疆骏枣疮痂病空间分布型测定及抽样技术研究[J].塔里木大学学报,2013,25(3):9-13.
[14]陈观浩,梁盛铭,任惠,等.南方水稻黑条矮缩病空间分布型及抽样技术[J].植物保护,2014,40(1):131-133.
[15]茹水江,王汉荣,王连平,等.西兰花菌核病空间分布格局及抽样技术研究[J].浙江农业报,2006,18(2):63-66.
[16]兰星平.Lα-m模型在检验昆虫种群空间分布型与抽样调查中的应用[J].林业科学研究,1996,9(1):41-46.
[17]张连翔,吕尚彬,温豁然,等.种群空间格局研究的Z-V模型及其抽样设计方法[J].西北林学院学报,1997,12(1):75-79.
[18]瞿小杰,段显德,何玉会,等.百日草白粉病病斑在叶片上的空间分布型及抽样技术研究[J].中国植保导刊,2014,34(6):46-50.
[19]张连翔,冯子娟,王伟,等.Iwao和 Taylor模型应用的局限性及补充[J].辽宁林业科技,1997(5):38-41.
[20]徐汝梅.种群数量的时空动态[M].北京:北京师范大学出版社,1990.
[21]兰星平,薛贤清.c′-m回归模型在确定昆虫种群空间分布型中的应用[J].南京林业大学学报,1996,20(2):59-64.
[22]兰星平,魏义贤,朱秀娥.松毒蛾空间分布型的研究[J].贵州林业科技,1992,20(1):8-19.
猜你喜欢
中国农业科学(2022年16期)2022-09-19
土壤学报(2022年1期)2022-03-08
甘肃教育(2021年10期)2021-11-02
食用菌(2019年4期)2019-08-27
中学生数理化·高二版(2017年3期)2017-07-07
现代农业科技(2016年23期)2017-04-06
电脑知识与技术(2016年31期)2017-02-27
天然产物研究与开发(2016年6期)2016-06-05
建材发展导向(2016年3期)2016-05-23
西北园艺(综合)(2016年2期)2016-02-19
