
基于投影寻踪模型的草原蝗虫栖境评价及风险评估
2015-04-15 08:03黄训兵吴惠惠秦兴虎曹广春王广君农向群涂雄兵格希格都仁贺兵额尔登巴图乌亚汗张泽华
草业学报 2015年5期
黄训兵,吴惠惠,秦兴虎,曹广春,王广君,农向群,涂雄兵,格希格都仁,贺兵,额尔登巴图,乌亚汗,张泽华*
(1.植物病虫害生物学国家重点实验室,中国农业科学院植物保护研究所,北京 100193;2.内蒙古自治区锡林郭勒盟镶黄旗草原工作站,内蒙古 锡林郭勒盟 013250)
基于投影寻踪模型的草原蝗虫栖境评价及风险评估
黄训兵1,吴惠惠1,秦兴虎1,曹广春1,王广君1,农向群1,涂雄兵1,格希格都仁2,贺兵2,额尔登巴图2,乌亚汗2,张泽华1*
(1.植物病虫害生物学国家重点实验室,中国农业科学院植物保护研究所,北京 100193;2.内蒙古自治区锡林郭勒盟镶黄旗草原工作站,内蒙古 锡林郭勒盟 013250)
草原蝗虫发生与栖境存在紧密而复杂的关系,二者关系的研究是评估蝗灾发生风险的基础。本文分析了不同栖境内蝗虫种群密度与21个植被特征参数的相关关系,利用投影寻踪模型进行了栖境评价及风险评估,并进行模型验证。结果表明,低优参数植物生物量多样性对蝗虫种群密度影响最大,最佳投影向量a为0.6725;高优参数禾本科生态优势度对亚洲小车蝗密度影响最大,最佳投影向量a为0.6547;样点植被投影特征值Zi与蝗虫种群密度线性相关关系极显著(y=48.861x-18.937,R=0.9509**),Zi越大,栖境内植被越适合蝗虫的发生,蝗灾发生的风险越高,根据Zi值可预测不同栖境草原蝗虫的发生。投影寻踪模型评价不同植被条件下蝗虫的发生风险,可以排除与数据结构和特征无关或关系很小变量的干扰,是一种更稳健实用的方法,对于蝗虫的监测预警具有重要意义。
草原蝗虫;草原植被;投影寻踪模型;风险评估
草原蝗虫发生与栖境内植被紧密相关,国内外学者已从能流模式、资源利用方式、共存机制等不同角度对蝗虫与植物的关系做了研究[1-9]。草原植被生长势受季节及气候条件影响,导致蝗虫生存环境变化及生态位中心转移[10]。典型对应分析(CAA)黑河流域13种蝗虫种群表明,按栖境植被可将蝗虫分为6个类群[11-12],禾本科和菊科植物对蝗虫种群空间分布的影响最大。通过冗余分析(RDA)及双重筛选逐步回归等方法分析内蒙古典型草原蝗虫与植被群落结构的关系表明,植物特征参数变化导致蝗虫特征参数变化,优势种植物决定优势种蝗虫的存在[13]。特别是最近有研究表明亚洲小车蝗(Oedaleusasiaticus)趋向取食含氮量低的植物,喜欢栖居于以大针茅(Stipagrandis)为优势种的低氮草原类型[14],低氮草原类型更利于蝗害的发生。而对亚洲小车蝗食性及营养生态位的研究发现,牧压增加引起的植被群落变化,导致亚洲小车蝗对克氏针茅(Stipakrylovii)的采食降低,增加了对米氏冰草(Agropyroncristatum)和星毛委陵菜(Potentillaacaulis)的采食,亚洲小车蝗生态位变窄[15]。高山草地毒杂草侵入能够改变植被结构和营养价值,以及蝗虫发生区域内的土壤理化特性和栖息生境,从而引起草地蝗虫群落组成、数量和多样性的变化[16]。植被显著影响蝗虫种群[17-18],对蝗虫发生起复杂而重要的生态效应。
蝗虫种群密度、生长和繁殖能力等受植被的显著影响,蝗害的发生与植被紧密相关[19-20]。对亚洲小车蝗的食性和食量的研究表明,亚洲小车蝗喜食禾本科植物,非喜食植物会负向影响其生长生殖状况[21],在禾本科中,与含氮量高的羊草相比,含氮量低的针茅最利于亚洲小车蝗生长发育[14]。对蝗虫生长发育过程中取食植被的研究表明,有些不喜食植被的存在有重要意义,与喜食植被混合能够促进蝗虫的生长发育[22],这可能与营养平衡和营养稀释有关[23-24]。蝗虫对栖境的选择是多方面的,各因子彼此联系,相互影响[11,18]。
以上研究从植被因子与蝗虫发生关系的不同角度进行了深入探索,但缺乏蝗虫栖境植被条件的综合评价模型及不同植被条件下蝗虫发生风险评估的可靠方法。而随着对草原生态环境保护的重视[25],有必要建立栖境风险评价模型来指导蝗害防控。因此,本研究在2011年、2012年草原蝗虫和植被调查的基础上,旨在提出基于遗传算法的投影寻踪模型综合评价方法,并对方法的可靠性进行验证,以期实现植被综合评价及蝗虫发生的风险评估,这一方法的实现对于草原蝗虫的监测预警、宜生区划分、生态治理等具有重要意义。
1 材料与方法
1.1 研究区概况
试验地位于内蒙古锡林郭勒盟镶黄旗,地理坐标:北纬42°15′-42°25′,东经113°45′-113°83′,海拔1300 m。该区域属中温带半干旱大陆性季风气候的典型草原,年降水量267.9 mm,主要降水集中在夏季,平均气温3.1 ℃,无霜期平均为120 d左右,最大冻土深度154 cm,土壤类型为栗钙土,调查区域内的气候背景相同,地势相对平坦,2011和2012年草原蝗虫发生盛期分别选取了9和13块草原样地进行植被和蝗虫调查。
1.2 试验材料
样方框(1 m×1 m)、捕虫网、剪刀、纸袋、 烘箱、电子天平(1/1000)、GPS定位仪等。
1.3 草原蝗虫与植被调查
植被调查采用五点取样法,每样地随机选取5个样点,以样方框取样,样点面积1 m2,调查植物的种类、密度、生物量等,详细方法见参考文献[26-27]。
蝗虫数量调查以扫网的方式进行,避开样地内因微环境形成的不同植物结构,每样地重复5次,每次100复网,已扫网区域不能重复进入,对每100复网中的蝗虫种类及数量(头/100网)进行统计。
1.4 数据处理
1.4.1 植被群落特征参数分析 计算2011年9个样地植被的21个特征参数(表1),计算公式如下:



式中,Pi=ni/N,i:第i个物种;ni:第i个物种的个体数;N:所有物种个体数之和;Pi:群落相对密度;S:物种数(即植被丰富度)。
将蝗虫种群密度及亚洲小车蝗密度同21个植被特征参数作相关性分析(SAS 8.0),找出相关性较好的植被特征参数作为评价指标,与蝗虫密度正相关的指标为高优指标,负相关的为低优指标,利用基于遗传算法的投影寻踪模型[28-32]进行数据处理(Matalab R2009b编程),得到投影特征值后与蝗虫种群密度作线性回归分析(SAS 8.0)。
2012年的实验数据作为模型的验证数据,计算由2011年数据相关性分析得到的高优和低优指标,之后利用基于遗传算法的投影寻踪模型进行综合评价,并分析投影特征值与蝗虫种群密度关系。
1.4.2 基于遗传算法的投影寻踪综合评价方法 投影寻踪是一种处理多因素复杂问题的统计方法, 其基本思路是将高维数据向低维空间进行投影, 通过低维投影数据的散布结构来研究高维数据特征[29-32]。具体数据处理过程为:
I)归格化处理。对相关性分析得到的植被低优指标和高优指标进行规格化处理,设p个指标n个样本集的原始数据为(Xij)n×p,对越大越优的评价指标(高优指标):
(1)
对于越小越优的评价指标(低优指标):
(2)
其中min(Xj)、max(Xj)分别为第j个评价指标的最小值和最大值。
II)构造投影指标函数[Q(a)]。将规格化后的p维数据{Yij|j=1, 2, ...,p},综合成以a={a1,a2,a3, …,ap}为投影向量的一维投影值Zi。
(3)
根据Zi值进行排序,最理想的综合投影值的散布特征是局部投影点尽可能的密集,凝聚成若干点团,在整体上投影点团之间尽可能分散,因此,投影指标函数可以表达成为:
Qz=SzDz
(4)

III)优化投影指标函数。当各植被指标值样本集一定时,投影函数Q只与投影向量a有关,即以下优化问题:
目标函数:maxQ(a)=SZDZ

(5)
这是一个关于p维向量a的非线性的优化问题,本研究使用收敛性好、全局优化性能优和使用性强的基于实数编码的加速遗传算法对该优化问题进行求解,求解过程采用Matalab R2009b编程求解。

表1 植被特征参数与蝗虫总密度、亚洲小车蝗密度相关性分析
A:植物群落种类多样性Plant diversity;B:植物群落优势度Plant community dominance;C:植物垂直层次结构多样性Plant vertical diversity;D:植物生物量多样性Plant biomass diversity;E:植物丰富度Plant richness;F:均匀度Plant evenness;G:植被盖度(100%)Vegetation coverage;H:针茅生态优势度Stipakryloviidominance;I:羊草生态优势度Leymuschinensisdominance;J:禾本科生态优势度Poaceae dominance;K:菊科生态优势度Asteraceae dominance;L:豆科生态优势度Leguminosae dominance;M:百合科生态优势度Liliaceae dominance;N:杂类草生态优势度Fobs dominance;O:针茅生物量Stipacapillatabiomass (g/m2);P:羊草生物量Leymuschinensisbiomass (g/m2);Q:禾本科生物量多样性Poaceae biomass diversity;R:豆科生物量多样性Leguminosae biomass diversity;S:菊科生物量多样性Asteraceae biomass diversity;T:百合科生物量多样性Liliaceae biomass diversity;U:杂类草生物量多样性Fobs biomass diversity;V:蝗虫总密度Grasshoppers density (Individual/100 net);W:亚洲小车蝗密度O.asiaticusdensity (Individual/100 net).“P<0.05” 表示蝗虫总密度(或亚洲小车蝗密度)与植被特征参数显著相关。“P<0.05”indicated that the correlation between grasshopper density (orO.asiaticusdensity) and character parameters of vegetation was significant.
IV)利用得出的最佳投影向量ai求得各个样本点的投影值Zi。若Zi和Zj的取值接近,认为第i个样本和第j个样本趋于同一类。若按Zi值从大到小排序,可以将样本从优到劣进行排序。
2 结果与分析
2.1 蝗虫种群总密度和亚洲小车蝗密度与植被特征参数的相关性分析
蝗虫密度与21个植被特征参数的相关性分析表明(表1),蝗虫总密度(头/100网)与针茅生态优势度显著正相关(P<0.05),与栖境内植被丰富度、羊草生态优势度、植物生物量多样性及菊科生物量多样性显著负相关(P<0.05)。亚洲小车蝗密度与克氏针茅生态优势度及禾本科生态优势度显著正相关(P<0.05),与栖境内植被丰富度、羊草生态优势度、植物生物量多样性及羊草生物量显著负相关(P<0.05)。相关性分析结果能够表明对蝗虫密度存在正负显著影响的植被特征参数,是植被综合评价及风险评估研究的基础。
2.2 蝗虫种群栖境植被的综合评价
由表2可知,在所有植被特征参数中,低优指标植物生物量多样性对蝗虫总密度影响最大(最佳投影向量a=0.6725),即植被生物量多样性越大,蝗虫总密度越低,其他依次为低优指标植被丰富度(a=0.4548)、高优指标针茅生态优势度(a=0.4118)、低优指标菊科生物量多样性(a=0.4081)和低优指标羊草优势度(a=0.3195)。通过表3不同样地投影特征值可知,2号样地投影特征值Zi最大,其次是9号样地,其他依次为样地6,1,5,8,7,4和3,Zi越大越适合蝗虫的发生。各样点蝗虫总密度与植被投影寻踪特征值的线性回归分析表明(图1),二者极显著(P<0.01)线性相关(y=48.861x-18.937,R=0.9509),表明投影特征值能够很好地反映蝗虫栖境植被适合蝗虫发生的程度,按照投影寻踪聚类的方法[31],9个样地可划分为两类,2和9号样地为I类,其余样地为II类,I类区域较II类区域更适合蝗虫发生。

表2 蝗虫栖境内植被参数的最佳投影方向

表3 不同样地植被参数投影特征值
2.3 亚洲小车蝗栖境植被的综合评价
由表4可知,对于亚洲小车蝗,高优指标禾本科生态优势度对其影响最大(投影向量a=0.6547),禾本科优势度越大,亚洲小车蝗密度越大,其他依次为高优指标针茅生态优势度(a=0.4679)、低优指标羊草生物量(a=0.3969)、低优指标羊草生态优势度(a=0.3865)、低优指标植被丰富度(a=0.3371)和低优指标植物生物量多样性(a=0.1601)。由表5不同样地投影特征值Zi可知,样点2投影特征值最大,依次为样地9,5,1,6,4,3,8和7,Zi值越大栖境植被越适合亚洲小车蝗的发生。各样点亚洲小车蝗密度与栖境内植被投影寻踪特征值Zi的线性回归分析表明(图2),二者极显著(P<0.01)线性相关(y=4.3324x-1.3258,R=0.9295),可见投影寻踪特征值Zi能够很好地反映栖境植被对亚洲小车蝗发生的适合度,评价亚洲小车蝗的发生。按照投影寻踪聚类的方法[31],9个调查样地可分为两类,样地2为I类,其余样点为II类,I类植被栖境较II类更适合亚洲小车蝗发生。
2.4 基于投影寻踪模型的栖境风险评估及模型验证

图1 蝗虫总密度与栖境内植被投影特征值关系Fig.1 Relationship between grasshopper density and projection eigenvalue (2011)
可靠性是衡量模型应用价值的重要因素,以2012年调查得到的数据作为验证,计算由2011年数据分析得到的低优和高优指标,利用基于遗传算法的投影寻踪模型进行综合评价,分析投影特征值与蝗虫密度的关系(图3和4),结果表明,蝗虫总密度与综合评价其栖境内植被的特征值Zi线性相关关系极显著(y=26.425x-9.4970,R=0.8585**),与2011年分析结果相同的是,特征值Zi很好地反映了栖境植被适合蝗虫发生的程度,同样,亚洲小车蝗密度与其栖境内植被特征值也表现为极显著线性相关(y=6.6056x-2.8964,R=0.9087**),利用植被投影特征值可以对不同栖境内蝗虫的发生进行风险评估,栖境植被投影特征值越大,蝗虫发生的风险越大;投影特征值越小,蝗虫发生的风险越小,投影寻踪模型表现了很好的适用性和可靠性。

表4 亚洲小车蝗栖境内植被参数的最佳投影方向(2011)

表5 不同样地植被参数投影特征值(2011)
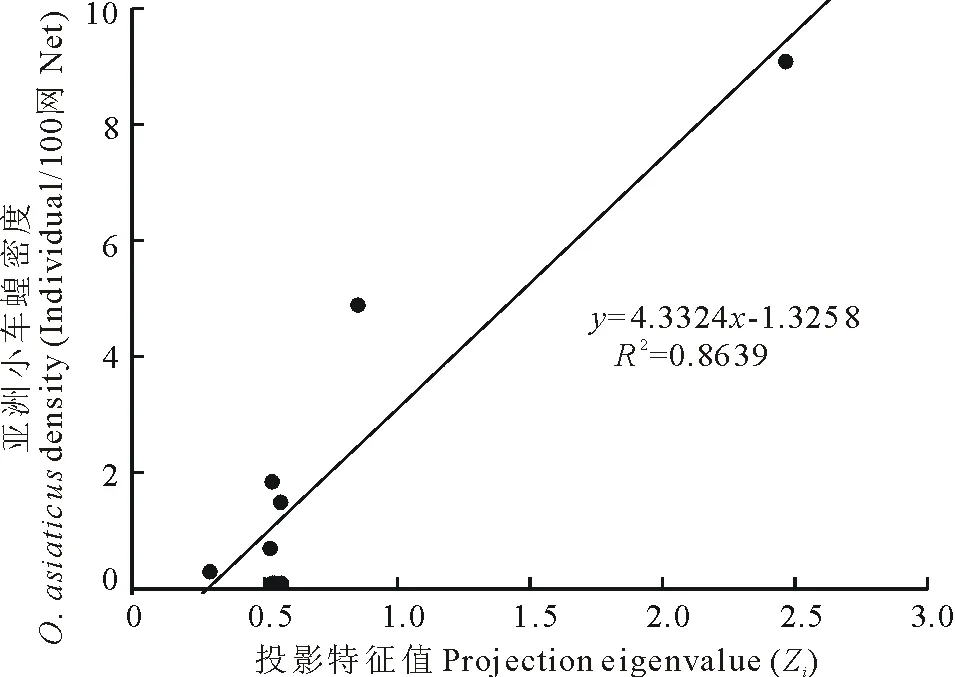
图2 亚洲小车蝗密度与栖境内植被投影特征值关系(2011)Fig.2 Relationship between O. asiaticus density and projection eigenvalue (2011)
3 讨论
投影寻踪模型是处理多因素复杂问题的统计方法,已广泛应用于水质评价、环境监测、灾情评估、水资源评价等许多领域[31-36]。与主成分分析等其他评价模型相比,投影寻踪模型本身对数据和样本容量并无特殊要求,且可以排除与数据结构和特征无关的,或关系很小的变量的干扰,因而是一种更稳健实用的方法[34,36-37],同时,投影寻踪聚类模型能够有效地克服灰色关联度计算值离散性不强、结果趋于均化的弱点,能够有效地解决各类实际问题,是一种客观、准确、可靠的综合评价模型,并表现出很好的适用性和稳定性[34,38-39]。本文利用基于遗传算法的投影寻踪模型,对蝗虫栖境植被综合评价, 分析了影响蝗虫发生的主要植被因子,投影特征值是植被综合评价的指数,与蝗虫种群密度呈显著线性相关,Zi越大,蝗虫种群密度越大,根据Zi的大小能够对栖境内植被适合蝗虫发生的程度进行评估,通过数据验证,基于遗传算法的投影寻踪模型能够可靠的评估栖境植被与蝗虫发生的关系。

图3 蝗虫总密度与栖境内植被投影特征值关系(2012)Fig.3 Relationship between grasshopper density and projection eigenvalue (2012)

图4 亚洲小车蝗密度与栖境内植被投影特征值关系(2012)Fig.4 Relationship between O. asiaticus density and projection eigenvalue (2012)
植被与蝗虫之间存在着紧密而复杂的关系,植物不仅为蝗虫提供食料资源,也为蝗虫提供适宜的栖息地[11,19-20,40]。本研究表明低优指标植物生物量多样性对蝗虫种群的影响最大,主要是因为调查区域内以喜食禾本科的亚洲小车蝗、毛足棒角蝗和宽须蚁蝗为主,植被生物量越分散,蝗虫发生密度越低,而作为高优指标的针茅生态优势度越大,越利于蝗虫的发生。与颜忠诚和陈永林[2]、康乐等[4]研究结果一致的是禾本科对亚洲小车蝗的发生影响最大,但是,在不同的禾本科植物中,针茅生态优势度作为高优指标,羊草生态优势度及羊草生物量作为低优指标影响亚洲小车蝗发生,且二者中以针茅对亚洲小车蝗的影响最大,这与吴惠慧等[13]及Cease等[14]的研究结果一致。
环境因子诸如气象因子、土壤类型、生态地理特征等影响蝗虫的发生[41],本研究只在同一区域内(气象因子、土壤类型等一致)对不同栖境植被与蝗虫发生的关系进行分析与评价,而对不同区域内的研究,还要考虑气象因子、土壤类型、地理环境等多种因素[41],在此基础上利用基于遗传算法的投影寻踪模型对蝗虫发生的不同区域生境条件适合蝗虫发生的程度进行综合评价,对蝗害的发生进行风险评估需要做进一步的研究。另外,本文只通过相关性分析获得影响蝗虫发生的植被低优指标和高优指标,而不同植被因子对蝗虫的发生存在间接或直接的复杂影响,对于这些植被因子的深入研究和利害影响的深度挖掘,是植被条件综合评价及蝗虫发生风险评估进一步研究的基础。
总之,利用基于遗传算法的投影寻踪模型及其得到的投影向量、投影特征值能够综合评价栖境植被适合草原蝗虫发生的程度,能够准确、可靠地评估蝗虫发生风险,具有很好的适用性及应用价值。该综合评价方法的建立,对于草原蝗虫的监测预警、宜生区划分及生态治理等具有重要的意义。
4 结论
基于遗传算法的投影寻踪模型能够对蝗虫栖境植被进行综合评价,评估植被条件适合蝗虫发生的程度。作为低优指标的植物生物量多样性对蝗虫种群密度影响最大,作为最高优指标的禾本科生态优势度对亚洲小车蝗密度影响最大,其中针茅生态优势度作为高优指标,羊草生态优势度为低优指标影响亚洲小车蝗的发生,且二者中以针茅对亚洲小车蝗的影响最大。投影特征值Zi与蝗虫密度极显著线性相关,Zi值反映了植被对蝗虫发生的适合度,根据Zi值可推算、预测蝗虫的发生情况。利用投影寻踪模型对蝗虫栖境内植被进行综合评价方法的建立,能够准确、可靠地评估蝗虫发生风险,具有很好的适用性及应用价值。
[1] Yan Z C, Chen Y L. Composition of grasshoppers species in different habitats in Xilin river basin, Inner Mongolia. Acta Entomologica Sinica, 1997, 40(3): 271-275.
[2] Yan Z C, Chen Y L.Habital selection of grasshoppers: relationship of habital selection and horizontal structure. Wuyi Science Journal, 1998, 14: 251-257.
[3] Qiu X H, Li H C. Energy dynamics ofChorthippusdubius(Zubovsky) populations in three different plant communities in Inner Mongolia grassland ecosystem. Acta Ecologica Sinica, 1993, 13(1): 1-8.
[4] Kang L, Li H C, Chen Y L. Relationship of orthopteran ecological distribution and vegetation types in Xilin river basin, Inner Mongolia. Acta Phytoecologica et Geobotanica Sinica, 1989, 13(4): 341-349.
[5] He D H, Zheng Z M, Liu Y D. Studies of grasshoppers spatiotemporal niche of desert steppe. Journal of Ningxia Agricultural College, 1997, 18(2): 1-9.
[6] Allan E, Crawley M J. Contrasting effects of insect and molluscan herbivores on plant diversity in a long-term field experiment. Ecology Letters, 2011, 14: 1246-1253.
[7] Guo Z W, Li H C, Gan Y L. Grasshopper (Orthoptera: Acrididae) biodiversity and grassland ecosystems. Insect Science, 2006, 13(3): 221-227.
[8] Awmack C S, Leather S R. Host plant quality and fecundity in herbivorous insects. Annual Review of Entomology, 2002, 47: 817-844.
[9] Scherber C, Eisenhauer N, Weisser W W,etal. Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature, 2010, 468: 553-556.
[10] Knop E, Schmid B, Herzog F. Impact of regional species pool on grasshopper restoration in Hay Meadows. Restoration Ecology, 2008, 16(1): 34-38.
[11] Zhao C Z, Zhou W, Wang K M,etal. Relationship between ecological distribution of grasshoppers and their habitats in the middle and upper reaches of Heihe River. Journal of Lanzhou University (Naturnal Sciences), 2009, 45(4): 42-47.
[12] Zhao C Z, Zhou W, Wang K M,etal. The CCA analysis between grasshopper and plant community in upper reaches of Heihe River. Acta Ecologica Sinica, 2011, 31(12): 3384-3390.
[13] Wu H H, Xu Y H, Cao G C,etal. Ecological effects of typical grassland types in Inner Mongolia on grasshopper community. Scientia Agricultura Sinica, 2012, 45(20): 4178-4186.
[14] Cease A J, Elser J J, Ford C F,etal. Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content. Science, 2012, 335: 467-469.
[15] Liu G H, Wang G J, Wang S P,etal. Study on the diet composition and trophic niche of main herbivores in the Inner Mongolia Typical steppe-TakingLeymuschinensiscommunity as an exemple. Acta Prataculturae Sinica, 2013, 22(1): 103-109.
[16] Su T, Chen Q, Zhao Y X,etal. Effects of invasive weeds on relative grasshopper abundance in alpine steppe in the Qilian Mountains. Acta Prataculturae Sinica, 2013, 22(3): 85-91.
[17] Franzke A, Unsicker S B, Specht J,etal. Being a generalist herbivore in a diverse world: How do diets from different grasslands influence food plant selection and fitness of the grasshopperChorthippusparallelus. Ecological Entomology, 2010, 35: 126-138.
[18] Rominger A J, Miller T E X, Collins S L. Relative contributions of neutral and niche-based processes to the structure of a desert grassland grasshopper community. Oecologia, 2009, 161(4): 791-800.
[19] Andersen A N, Ludwig J A, Lowe L M,etal. Grasshopper biodiversity and bioindicators in Australian tropical savannas: Responses to disturbance in Kakadu National Park. Austral Ecology, 2001, 26(3): 213-222.
[20] Belovsky G E, Jennifer B S. Dynamics of two Montana grasshopper populations: relationships among weather, food abandance and intraspecific competion. Oecologia, 1995, 101: 383-396.
[21] Xi R H, Liu J P. Effects onOedaleusasiaticusin growth and fecundity for feeding different foods. Entomological Knowledge, 1984, 4: 153-155.
[22] Kazumi M, Naota M. Diet mixing and its effect on polyphagous grasshopper nymphs. Ecological Research, 2004, 19: 269-274.
[23] Pulliam H R. Diet optimism with nutrient constraints. American Naturalist, 1975, 109: 765-768.
[24] Freeland W J, Janzen D H. Strategies in herbivory by mammals:he role of plant secondary compounds. American Naturalist, 1974, 108: 269-289.
[25] Ma Q Z. Protection of grassland ecology, and construction of beautiful pastoral areas. Acta Prataculture Sinica, 2014, 23(1): 1-2.
[26] Xu K, Li G Q, Li S Z. A comparison study on diversity of plant community of different utilization during recovering of degenerative steppe in Yanchi county, Ningxia. Acta Agriculturae Boreali-occidentalis Sinica, 2007, 16(4): 106-111.
[27] Shang Z H, Yao A X, Guo X S. Evaluation and review on measurement methods of biodiversity at home and abroad. Journal of Ningxia Agricultural College, 2002, 23(3): 68-73.
[28] Gao D W, Yao Y, Wu Y P. Evaluation of energy saving and consumption reduction in different regions based on projection pursuit classification model. Journal of Applied Statistics and Management, 2010, 29(3): 391-399.
[29] Guo J C, Li X L. The application of projection pursuit model in evaluation of enterprise performance. Finance and Accounting Monthly, 2009, 1: 49-51.
[30] Jin J L, Yang X H, Ding J. An improved simple genetic algori thm-accelerating genetic algorithm. Systems Engineering-Theory & Practice, 2001, 4(4): 8-13.
[31] Ni C J, Wang S J, Cui P. New model for projection pursuit dynamic cluster and its application to classifying natural grasslands. Journal of Safety and Environment, 2006, 6(5): 68-71.
[32] Dong X X, Wei X, Song L J. Comprehensive evaluation method for medical quality based on projection pursuit model. Chinese Journal of Health Statistics, 2012, 29(5): 636-638.
[33] Cao Y Q, Xing X S, Yi J M,etal. Application of projection pursuit technique in water security assessment of Wafangdian City. Water Resources Protection, 2012, 26(3): 5-7.
[34] Peng Y G, Luo X P, Wei W. New fuzzy adaptive simulated annealing genetic algorithm. Control and Decision, 2009, 24(6): 843-848.
[35] Jin J L, Wei Y M. Projection pursuit model for comprehensive evaluation of water quality. Acta Scientiae Circumstantiae, 2001, 21(4): 431-434.
[36] Yu G R, Ye H, Xai Z Q,etal. Improvement of projection pursuit classification Model and its application in evaluating water quality. Journal of Sichuan University (Engineering science edition), 2008, 40(6): 24-29.
[37] Jiang L, Li M L. Model of projection pursuit water quality evaluation based on immune evolutionary algorithm. Journal of Sichuan University (Natural Science Edition), 2004, 41(4): 816-819.
[38] Fu Q, Zhao X Y. Principles and Applications of Projection Pursuit Model[M]. Beijing: Science Press, 2006.
[39] Li R Z. Progress and trend analysis of theoretical methodology of water quality assessment. Journal of Hefei University of Technology, 2005, 28(4): 369-373.
[40] Bernays E A, Gonzalez N, Angel J. Food mixing by generalist grasshoppers: plant secondary compounds structure the pattern of feeding. Journal of Insect Behavior, 1995, 8(2): 161-180.
[41] Gong A Q, Wang W J, Zhang S H. Research of correlation between grasshopper and environmental factor in the region around Qinhai lake. Qinghai Prataculture, 2001, 10(2): 38-41.
参考文献:
[1] 颜忠诚, 陈永林. 内蒙古锡林河流域不同生境中蝗虫种类组成的分析. 昆虫学报, 1997, 40(3): 271-275.
[2] 颜忠诚, 陈永林. 草原蝗虫的栖境选择: 栖境选择与水平结构的关系. 武夷科学, 1998, 14: 251-257.
[3] 邱星辉, 李鸿昌. 草原生态系统狭翅雏蝗种群的能量动态. 生态学报, 1993, 13(1): 1-8.
[4] 康乐, 李鸿昌, 陈永林. 内蒙古锡林河流域直翅目昆虫生态分布规律与植被类型关系的研究.植物生态学与地植物学学报, 1989, 13(4): 341-349.
[5] 贺达汉, 郑哲民, 刘颖东. 荒漠草原蝗虫时空生态位的研究. 宁夏农学院学报, 1997, 18(2): 1-9.
[11] 赵成章, 周伟, 王科明, 等. 黑河中上游草原蝗虫生态分布与生境的关系. 兰州大学学报: 自然科学版, 2009, 45(4): 42-47.
[12] 赵成章, 周伟, 王科明, 等. 黑河上游蝗虫与植被关系的CAA分析. 生态学报, 2011, 31(12): 3384-3390.
[13] 吴惠惠, 徐云虎, 曹广春, 等. 内蒙古典型草原草地类型对蝗虫群落优势种群的生态效 应. 中国农业科学, 2012, 45(20): 4178-4186.
[15] 刘贵和, 王国杰, 汪诗平, 等. 内蒙古典型草原主要草食动物食性及其营养生态位研究-以羊草群落为例. 草业学报, 2013, 22(1): 103-109.
[16] 孙涛, 陈强, 赵亚雄, 等. 祁连山高山草地毒杂草侵入对蝗虫相对多度的影响. 草业学报, 2013, 22(3): 85-91.
[21] 席瑞华, 刘举鹏. 不同食料植物对亚洲小车蝗生长和生殖力的影响. 昆虫学报, 1984, 27(4): 153-155.
[25] 马启智. 保护草原生态、建设美丽牧区. 草业学报, 2014, 23(1): 1-2.
[26] 徐坤, 李国旗, 李世忠. 宁夏盐池县退化草地恢复过程中不同利用方式下植物群落多样性的对比研究. 西北农业学报, 2007, 16(4): 106-111.
[27] 尚占环, 姚爱兴, 郭旭生. 国内外生物多样性测度方法的评价与综述. 宁夏农学院学报, 2002, 23(3): 68-73.
[28] 高大伟, 姚奕, 仵雁鹏. 基于投影寻踪分类模型的各地区节能降耗评价研究. 统计与管理, 2010, 29(3): 391-399.
[29] 郭加昌, 李秀丽. 企业绩效评价中投影寻踪模型的应用. 财会月刊, 2009, 1: 49-51.
[30] 金菊良, 杨晓华, 丁晶. 标准遗传算法的改进方案-加速遗传算法. 系统工程理论与实践, 2001, 4(4): 8-13.
[31] 倪长健, 王顺久, 崔鹏. 投影寻踪动态聚类模型及其在天然草地分类中的应用. 安全与环境学报, 2006, 6(5): 68-71.
[32] 董小小, 魏歆, 宋丽娟. 投影寻踪模型在医疗质量综合评价中的应用. 中国卫生统计, 2012, 29(5): 636-638.
[33] 曹永强, 邢晓森, 伊吉美, 等. 投影寻踪技术在瓦房店市水资源安全评价中的应用. 水资源保护, 2012, 26(3): 5-7.
[34] 彭勇刚, 罗小平, 韦魏. 一种新的模糊自适应模拟退火遗传算法. 控制与决策, 2009, 24(6): 843-848.
[35] 金菊良, 魏一鸣. 水质综合评价的投影寻踪模型. 环境科学学报, 2001, 21(4): 431-434.
[36] 于国荣, 叶辉, 夏自强, 等. 投影寻踪分类模型的改进及在水质评价中的应用. 四川大学学报(工程科学版), 2008, 40(6): 24-29.
[37] 姜林, 李梦龙. 基于免疫算法优化的投影寻踪水质评价模型. 四川大学学报(自然科学版), 2004, 41(4): 816-819.
[38] 付强, 赵小勇. 投影寻踪模型原理及其应用[M]. 北京:科学出版社, 2006.
[39] 李如忠. 水质评价理论模式研究进展及趋势分析. 合肥工业大学学报(自然科学版), 2005, 28(4): 369-373.
[41] 巩爱岐, 王薇娟, 张生合. 青海湖滨区草地蝗虫发生与环境因素关联性的初步探讨. 青海草业, 2001, 10(2): 38-41.
Comprehensive evaluation and risk assessment of grasshoppers’habitat based on a projection pursuit model
HUANG Xun-Bing1, WU Hui-Hui1, QIN Xing-Hu1, CAO Guang-Chun1, WANG Guang-Jun1, NONG Xiang-Qun1, TU Xiong-Bing1, Gexigeduren2, HE Bing2, Eerdengbatu2, Wuyahan2, ZHANG Ze-Hua1*
1.StateKeyLaboratoryforBiologyofPlantDiseasesandInsectPests,InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China; 2.GrasslandWorkstationofXianghuangqiXilinguolemeng,Xilinguolemeng013250,China
There are close and complex relationships between grasshopper occurrence and habitat vegetation. A comprehensive analysis of these relationships will provide a stable foundation for risk assessments of grasshopper infection. Grasshopper population density and 21 vegetation parameters were analyzed in the survey on which this paper is based. A projection pursuit model was developed and verified in order to evaluate the risks of grasshopper infection. Results showed that a low index for plant biomass diversity had the greatest influence on grasshopper density, with the best projection directionaat 0.6725. Poaceae dominance with a high index had the greatest influence onOedaleusasiaticusdensity, with the best projection directionaat 0.6547. There was a significant linear relationship (P<0.01) between the projection eigenvalue (Zi) and grasshopper density (y=48.861x-18.937,R=0.9509). The occurrence of grasshoppers can be predicted according to the projection eigenvalue (Zi). The bigger the value ofZi, the higher the risk of grasshopper occurrence. The projection pursuit model can be used to eliminate the effect of irrelevant variables. Its application will play an important role in monitoring and early warning for the ecological management of grasshoppers.
grasshoppers; grassland vegetation; projection pursuit model; risk assessment
10.11686/cyxb20150504
http://cyxb.lzu.edu.cn
2014-05-22;改回日期:2014-06-30
公益性行业(农业)科研专项(201003079)和现代农业产业技术体系(CARS-35-07)资助。
黄训兵 (1990-), 男,山东临沂人,在读硕士。 E-mail: xunbingh@163.com *通讯作者Corresponding author. E-mail: zhangzehua@caas.cn
黄训兵, 吴惠惠, 秦兴虎, 曹广春, 王广君, 农向群, 涂雄兵, 格希格都仁, 贺兵, 额尔登巴图, 乌亚汗, 张泽华. 基于投影寻踪模型的草原蝗虫栖境评价及风险评估. 草业学报, 2015, 24(5): 25-33.
Huang X B, Wu H H, Qin X H, Cao G C, Wang G J, Nong X Q, Tu X B, Gexigeduren, He B, Eerdengbatu, Wuyahan, Zhang Z H. Comprehensive evaluation and risk assessment of grasshoppers’habitat based on a projection pursuit model. Acta Prataculturae Sinica, 2015, 24(5): 25-33.
猜你喜欢
小哥白尼(野生动物)(2022年5期)2022-08-15
数学物理学报(2021年6期)2021-12-21
数学物理学报(2021年5期)2021-11-19
数学物理学报(2021年3期)2021-07-19
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
快乐语文(2020年36期)2021-01-14
科学大众(2020年17期)2020-10-27
科学大众(2020年10期)2020-07-24
故事作文·低年级(2019年6期)2019-06-20
文苑(2018年22期)2018-11-19
