
长江口营养盐结构特征及其对浮游植物的限制
2015-04-11 03:26李亚力沈志良线薇微刘素美
海洋科学 2015年4期
李亚力, 沈志良, 线薇微, 刘素美
(1.中国海洋大学 化学化工学院, 山东 青岛 266100; 2.中国科学院 海洋研究所 海洋生态和环境科学重点实验室, 山东 青岛 266071)
日益增长的人类活动使得长江流域和长江三角洲地区的经济发展与长江口生态环境的矛盾日益突出, 长江口邻近海区富营养化逐年加剧[1], 有害赤潮高发, 已成为我国近海水域污染较为严重的地区之一[2]。由于化肥的使用和城市污水的排放, 使得海区DIN、 P O34--P含量增加。水域上游水利工程的修建,截流大量入海泥沙, 使得河口及其邻近海域 S iO32--Si含量减小, 进而导致N/P升高、Si/N下降。水体营养盐含量和结构变化会带来浮游植物群落组成的响应,以及浮游植物生长的区域差异和生物群落演替等一系列生态效应[3]。近年来研究报道, 在长江口外东海赤潮高发区, 硅藻赤潮的比例在逐渐降低, 而甲藻赤潮则呈逐渐增多趋势[4]。因此, 研究长江口营养盐结构特征, 探讨其对浮游植物的限制情况, 对认识水体富营养化的发展和赤潮的防治以及了解人类活动的生态学效应等都具有重要意义。
本文根据2013年春季和秋季两个航次最新调查数据, 分析了长江口营养盐浓度及其结构的分布变化, 并在此基础上探讨了营养盐对浮游植物的限制情况, 为该水域生态学研究提供背景支撑, 为长江口生态系统保护提供科学依据。
1 材料和方法
于 2013年 5月(春季)、11月(秋季)完成长江口及其邻近海域2个季度月的现场调查, 共设置39个调查站位(图1), 其中口门内包括35、36、37、38、39 站。调查内容主要包括硝酸盐(NO3--N)、亚硝酸盐(NO-2-N)、铵盐(NH+4-N)、磷酸盐( P O34--P)、硅酸盐(S iO32--Si)、叶绿素 a(Chl a)等。根据《海洋调查规范》(GB12763-2007), 分别用南森采水器采取表层、5、10、20、30 m、底层水样, 营养盐样品用Whatman GF/F膜(于450℃高温下处理6 h)现场过滤,储于聚乙烯瓶(预先在1∶10HCl溶液中浸泡24 h, 洗净烘干)中, 立即置于冰箱内冷冻保存, 带回实验室分析。
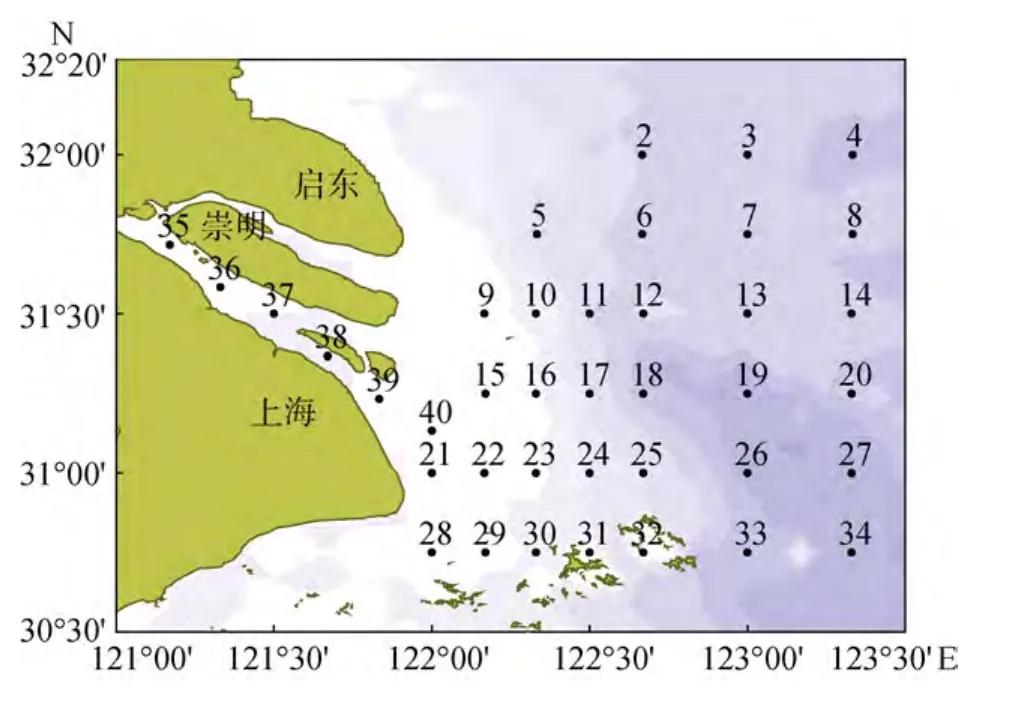
图1 长江口调查站位Fig.1 Sampling stations in the Yangtze River Estuary
叶绿素a采用丙酮萃取荧光法测定, NO3--N采用镉-铜还原法、NO-2-N采用重氮-偶氮法、NH+4-N采用水杨酸钠法、PO34--P采用磷钼蓝法、SiO32--Si采用硅钼蓝法测定, 各项营养盐用德国产Quattro营养盐连续流动分析仪测定, NO3--N、NO-2-N、NH+4-N、PO34--P、SiO32--Si测定的检出限、精密度分别为 0.02 µmol/L、1.6%; 0.02 µmol/L、0.3%;0.03 µmol/L、3.3%; 0.01 µmol/L、1.3%; 0.04 µmol/L 、1.1%。由于调查时间均在月初, 长江营养盐输出通量均是按照4、10月流量以及35号站位(淡水端)浓度进行估算。溶解无机氮(DIN)为NO3--N、NO-2-N、NH+4-N的和。
2 结果与讨论
2.1 营养盐浓度和分布特征
2.1.1 营养盐浓度和平面分布
表1列出了口门内和口门外春、秋季表、底层海水中营养盐的平均浓度及变化范围。由表1可见,口门内 DIN平均浓度秋季较高, 春季丰水期较低。口门外 DIN的平均浓度春季较高, 浓度变化范围也较秋季大, 春、秋季 DIN表层浓度均高于底层。口门外NO3--N、NO-2-N表、底层浓度均为春季高于秋季, 而NH+4-N表、底层浓度秋季远远高于春季, 春、秋季长江NO3--N、NO-2-N、NH+4-N的输出通量分别为37.65、0.12、0.04 kg/s和33.20 、0.08、0.8 kg/s,说明3种DIN季节变化受长江输送影响较大。从图2看出, 春、秋季NO3--N浓度均呈现河口及其附近高、向外海逐渐降低的趋势, 秋季 11月份长江径流量较春季 5月份小(5、11月份径流量分别为905.30×108、334.37×108m3), NO3--N 等值线向南偏移, 高浓度NO3--N分布区向河口收缩, 这可能是受南下黄海沿岸流的影响[5-6]。春、秋季NO-2-N高值区均在口门附近, 春季东南部浓度较低, 秋季东南部浓度较高。NH4+-N的分布除了受长江径流影响外还受生物活动及化学因素的影响, 故其分布较复杂,春季NH4+-N高值区主要在调查区的西南和东北部,其中西南部高值区位于上海市排污口附近, 两高值区之间的大片水域NH4+-N浓度均在0.2~0.3 µmol/L之间, 秋季NH4+-N存在西部、东北、东南三个高值区, 三高值区之间NH4+-N浓度均在1.2~1.6 µmol/L之间。
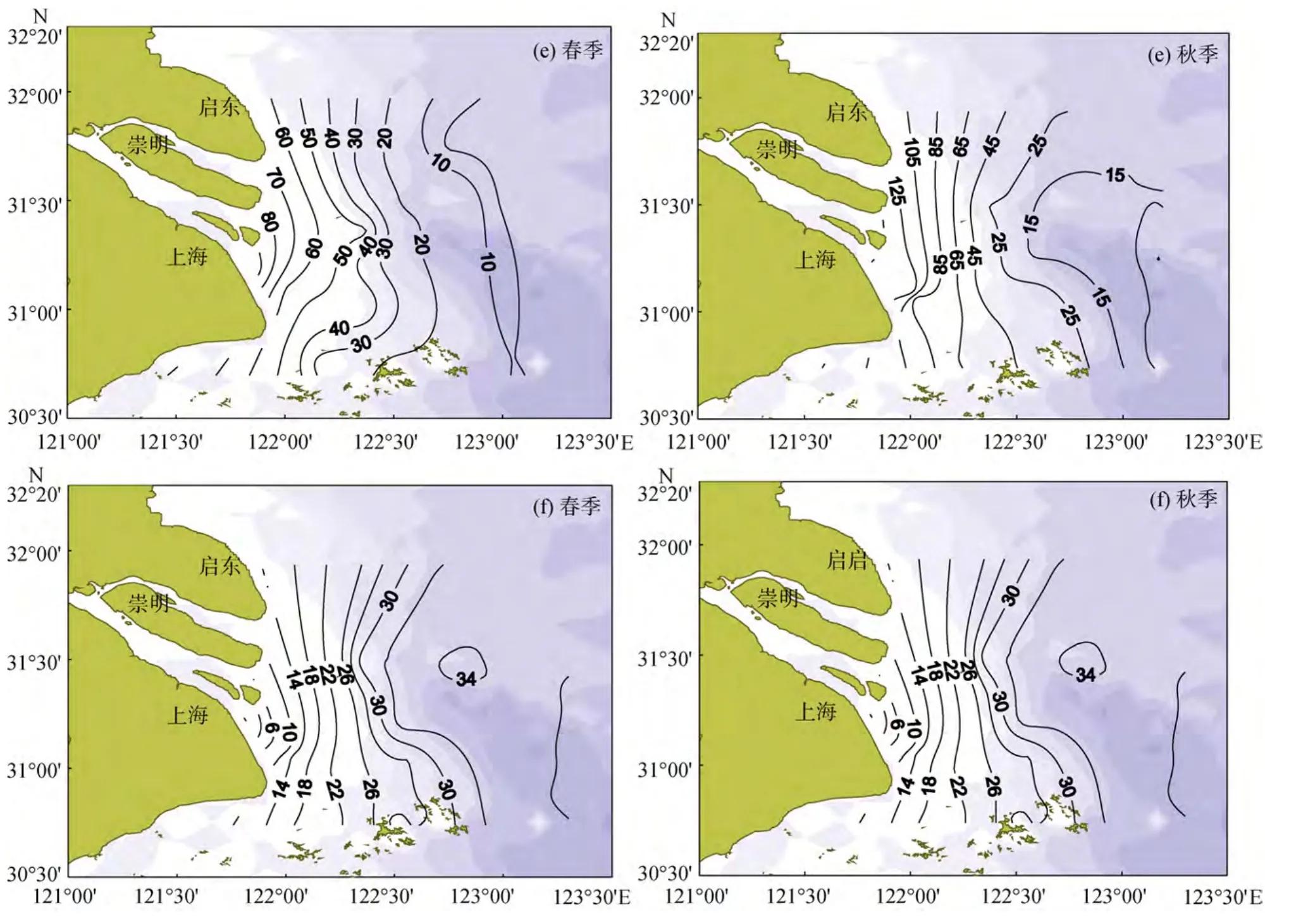
图2 长江口春、秋季表层营养盐和盐度平面分布图Fig.2 The surface horizontal distribution of nutrients and salinity in the Yangtze River Estuary in spring and autumn
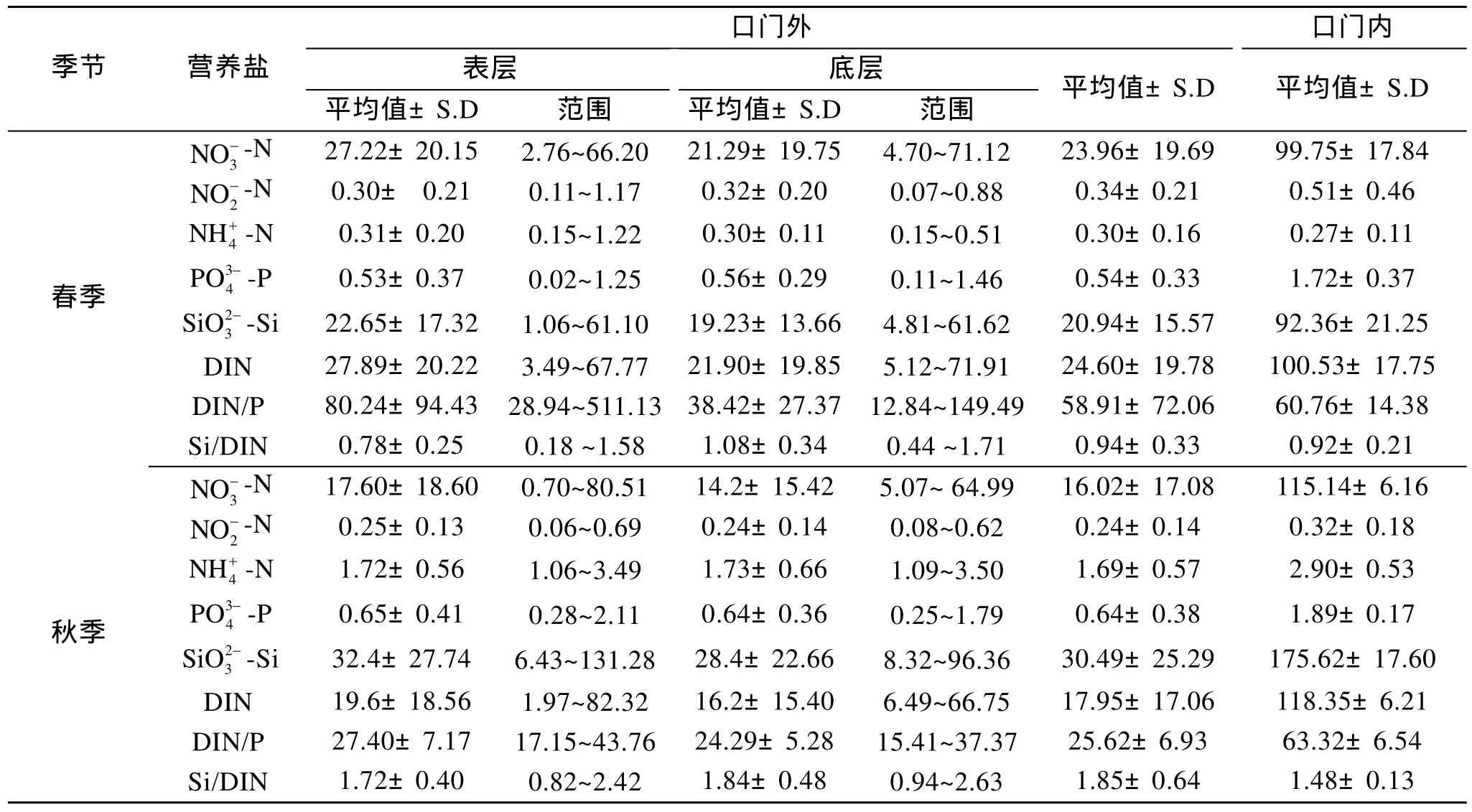
表1 长江口营养盐浓度(µmol/L)Tab.1 Concentrations of nutrients in Yangtze River Estuary(µmol/L)
口门内、外PO34--P浓度均秋季高于春季, 其一方面原因可能与悬浮颗粒物释放出 P O34--P有关, 秋季口门内悬浮颗粒物的平均质量浓度高达 405.26 mg/L,远远高于春季 35.31 mg/L, 因此秋季长江带入的悬浮颗粒物也远多于春季。另一方面可能是夏季大量繁殖的浮游生物在秋季死亡分解, 有机磷转换为PO34--P并在海水中积累。秋季水体垂直交换较好,PO34--P能从底层水体被连续不断的带入表层水体,故 P O34--P各水层分布相对均匀, 表、底层浓度差别较小。春季存在温度跃层[7], 水体交换较差以及生物活动较强, 使PO34--P浓度表层低于底层。PO34--P的分布一般为口门附近高、外海低, 主要受长江冲淡水的影响[8]。但由于PO34--P存在河口缓冲作用, 分布变化较其他营养盐复杂, 与NO3--N和SiO32--Si分布不同的是PO34--P浓度最高值一般不在口门内而是在口门外(如秋季 P O34--P浓度最高值2.11 µmol/L在40号站位)。由于人类活动的影响, 长江口N营养盐浓度逐年升高, 而 P O34--P浓度本身较小, 又由于河口缓冲的作用, 其增长趋势不大明显, 但在该调查区域内, PO34--P浓度与20世纪80年代调查结果相比也有所增加[9]。
春、秋季口门内、外 S iO32--Si浓度均较高, 其等值线分布与海岸线基本平行, 浓度从河口向外海逐渐降低, 表明 S iO32--Si主要受长江径流的影响, 该调查结果与以往的研究结果类似[10]。春季表、底层SiO32--Si浓度在口门内、外都远低于秋季, 其原因除了与径流输送有关外(春、秋季长江 S iO32--Si输出通量分别为 70.93、107.96 kg/s), 还受生物活动影响,春季浮游植物大量繁殖, 消耗较多的 S iO32--Si, 春季表层Chl-a质量浓度平均值为1.35 µg/L, 远高于秋季0.78 µg/L。
2.1.2 DIN的形态特征
海水中的DIN包括NO3--N、NO-2-N、NH+4-N 3种形态, 它们对海洋生物饵料循环起着至关重要的作用[11]。从图3可见在长江口及其邻近海域春、秋季表、底层DIN的主要存在形式都是NO3--N, 春季表、底层NO-2-N和NH+4-N的含量相近, 秋季表、底层NH+4-N的含量大于NO-2-N。春季表、底层NO3--N、NO-2-N、NH+4-N占总溶解无机氮的百分比分别是94.76%、2.77%、2.47%和95.43%、2.21%、2.36%; 秋季表、底层NO3--N、NO-2-N、NH+4-N占总溶解无机氮的百分比分别是 84.07%、2.17%、13.76%和82.51%、2.28%、15.21%。可以看出NO3--N、NO-2-N的相对含量为春季高于秋季, 而NH+4-N相对含量秋季高出春季约5倍。秋季口门内NH+4-N高,外海区亦高, 春季口门内NH+4-N低, 外海区亦低,表明秋季NH+4-N浓度高于春季主要是受长江输送影响。此外, NH+4-N还可能与生物活动有关, 长江口NH+4-N的主要来源是长江沿岸城市工、农业及生活污水排放, 在由河口向外海的输送过程中, 部分NH+4-N会被亚硝化菌和硝化菌氧化为NO-2-N和NO3--N, 这种硝化反应会随着温度的升高而增强[12],秋季水温高于春季(春、秋季海水温度平均值分别为17.61℃、20.69℃), 细菌的活动加强, 加快了有机质的降解[11]。
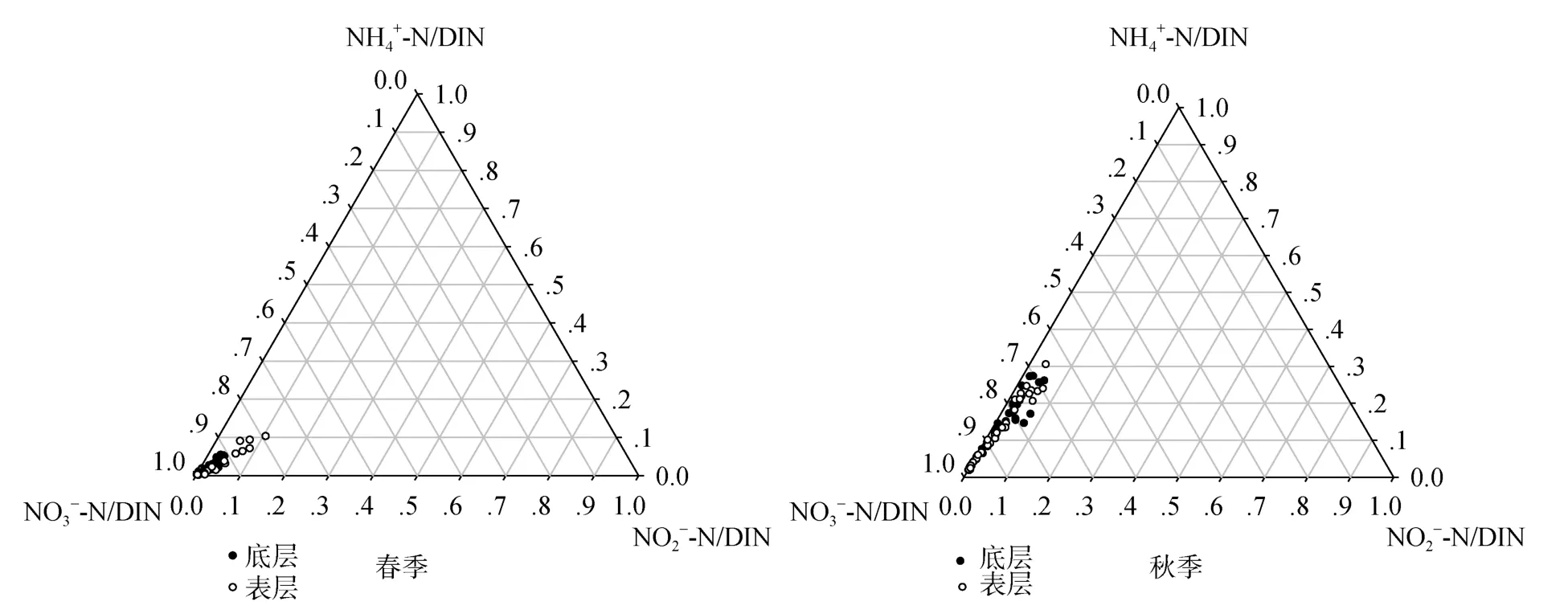
图3 长江口春、秋季NO3--N、NO-2-N、NH+4-N的相对含量Fig.3 The relative content of NO-3-N,NO-2-N andNH+4-N in the Yangtze River Estuary in spring and autumn
2.2 营养盐的结构分布特征
春季, 长江口及其邻近海域 Si/DIN值(S iO32--Si/DIN)大部分小于 1(见图4), 长江河道内其平均值、变化范围分别是0.92和0.81~1.13。海水中DIN主要来自工农业生产和生活废水的排放[13], 而SiO32--Si主要来自河流携带的岩石风化[14], 长期监测数据表明 DIN排放量逐年升高, 而 S iO23--Si溶出量逐年降低, 因此输入海水的长江水体中 Si/DIN值呈降低趋势[15]。此外, 图4中北-西北部存在一舌状Si/DIN高值区, 可能与苏北沿岸流和黄海沿岸流有关[16], 东北部 31.5°~32°N、122.75°~123.25°E 海区存在一 Si/DIN低值区, 最低至 0.18, 此处正位于长江口赤潮高发区[17-18], 也是叶绿素高值区, 7号站位(31.75°N, 123°E)叶绿素值最高, 可达 10.23 µg/L,SiO32--Si浓度则低至 2.05 µmol/L。有些站位(如31.5°N, 123°E)更是低至 1.06 µmol/L, 进而导致异常低的Si/DIN值。春季口门内、外DIN/P(DIN/PO34--P)平均值分别为 60.76和 58.91, 且长江河道内 DIN/P变化范围是46.77~149.49, 暗示河流输入以高DIN/P值为特征, 与以往研究结果相符[19]。此外, 长江口东北部赤潮高发区内的 Si/DIN低值区对应DIN/P的高值区, 最高可达270以上, 叶绿素值最高的 7号站位对应的PO34--P浓度为 0.04 µmol/L,部分站位更是低至 0.02 µmol/L, 这种局部异常应该与赤潮高发区内浮游植物短时间吸收大量PO34--P有关。
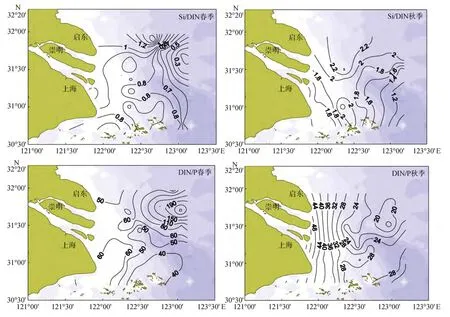
图4 长江口春、秋季表层Si/DIN和DIN/P平面分布Fig.4 The surface horizontal distribution of Si/DIN and DIN/P in the Yangtze River Estuary in spring and autumn

秋季, 口门内、外 DIN/P平均值分别为 63.32和 25.62, 其高值区位于口门附近, 并由近岸向外海递减, 最高值73.27位于长江河道里。因口门外DIN平均浓度比春季低, SiO32--Si平均浓度比春季高,故Si/DIN秋季比春季高, 正如图4所显示在调查区内Si/DIN均大于1, 北部部分区域大于2。长江口春、秋季口门内 Si/DIN的平均值分别是 0.92、1.48, 可看出秋季长江水体以高Si/DIN为特征。在调查区北部Si/DIN呈现一舌状高值区, 可能是受高 S iO32--Si、低DIN的苏北沿岸流影响[20]。
2.3 营养盐限制的判断
2.3.1 潜在相对限制
Redfield[21]提出海洋浮游植物是按一定的比例吸收水体中的碳、氮、磷元素, 即 C∶N∶P=106∶16∶1(此值被称作 Redfield比值), 硅藻一般吸收硅和氮的摩尔比是 1∶1[22], 这是判断浮游植物受哪种营养盐限制的基础[23]。虽不同藻类的限制阈值不同,但对于尺度较大的河口浮游植物, 仍可以依据Redfield比值有效的进行营养盐潜在相对限制的判断[24]。本文采用 Si∶DIN=1∶1、DIN∶P=16∶1作为营养盐潜在相对限制的判别依据。
图5是春、秋季表层营养盐摩尔浓度比的散点分布图。由于受含过量 S iO23--Si和 DIN[13]的长江和钱塘江等陆地径流的影响, 调查区域内春、秋季营养盐皆为PO34--P潜在相对限制。此外, 春季的数据点较秋季分散, 且大部分位于Si∶DIN=1∶1斜线的下方, 虽然长江峰面区 Si/DIN一直在下降(从 1959年的 3.8到 2002年的 0.85[15]), 但相比之下其仍然受PO34--P潜在相对限制。而秋季数据点集中, 数据点几乎全部位于 Si∶DIN=1∶1斜线的上方, 且有向DIN潜在相对限制转变的趋势。
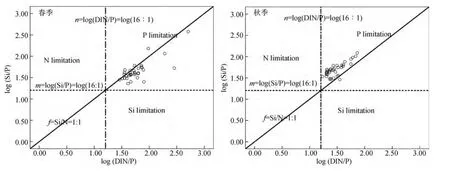
图5 长江口春、秋季表层营养盐浓度比值散点分布Fig.5 The scatter distribution of nutrient ratios in the Yangtze River Estuary in spring and autumn
2.3.2 绝对限制
每一种营养盐化学计量限制因素的成立并不代表实际限制因素就成立, 只有通过比较环境中营养盐的浓度和可能限制营养盐吸收的浓度之后才能确定可能的限制因素[25]。基于对营养盐吸收动力学的研究, Nelson 等[26]提出: DIN:1 μmol/L、P:0.1 μmol/L、Si:2 μmol/L为浮游植物生长所需要的最低阈值。Fisher等[27]也提出营养盐的限制浓度为 DIN:2 μmol/L,P:0.2 μmol/L; 本文在讨论绝对限制浓度时采用前者作为判断标准。
受长江冲淡水的影响, 调查区域内春、秋季均不存在 DIN的绝对限制站位(表2)。春季PO34--P绝对限制的站位除34站位外均位于叶绿素高值区, 即赤潮高发区以内, 说明浮游植物大量生长繁殖导致PO34--P被过量消耗, 从而低于阈值0.1μmol/L。春季SiO32--Si的绝对限制站位亦除34站位均位于赤潮高发区, 与PO34--P一致, 说明浮游植物也吸收了大量的 S iO32--Si。东海赤潮高发区浮游植物优势种呈“硅藻—甲藻—硅藻”的演替态势[28], 且 N/P随硅藻的生长而迅速升高, 并伴随消耗大量的 S iO32--Si[29],因此可判断春季长江口海域浮游植物优势种可能是硅藻。34站位存在PO34--P、SiO32--Si绝对限制, 其原因可能与低营养盐的黑潮表层水入侵有关, 这说明黑潮表层水中 S iO32--Si相对较为缺乏。此外, 因秋季海水的垂直混合作用相对强烈, 营养盐浓度相对较高, 故秋季不存在营养盐绝对限制。

表2 长江口春、秋季表层DIN、 P O43--P和 S iO23--Si绝对限制站位统计Tab.2 The statistics of the stations which appear DIN, P O43--P and SiO23--Si absolute limitations in the Yangtze River Estuary in spring and autumn
2.3.3 营养盐结构对浮游植物生长的影响
近年来随着 DIN和 P O34--P含量的增加,SiO32--Si含量的降低, DIN/P和Si/DIN比例也随之变化[8,30]。从表3可知当同时考虑绝对限制和潜在相对限制时, 20世纪80年代长江口及其邻近海域不存在氮、磷、硅营养盐对浮游植物的限制, 从90年代开始出现 P O34--P限制, 且2003年 P O34--P限制情况较90年代更加突出, 这种变化趋势不得不引起人类对该海区浮游植物生长和群落组成更大的关注。
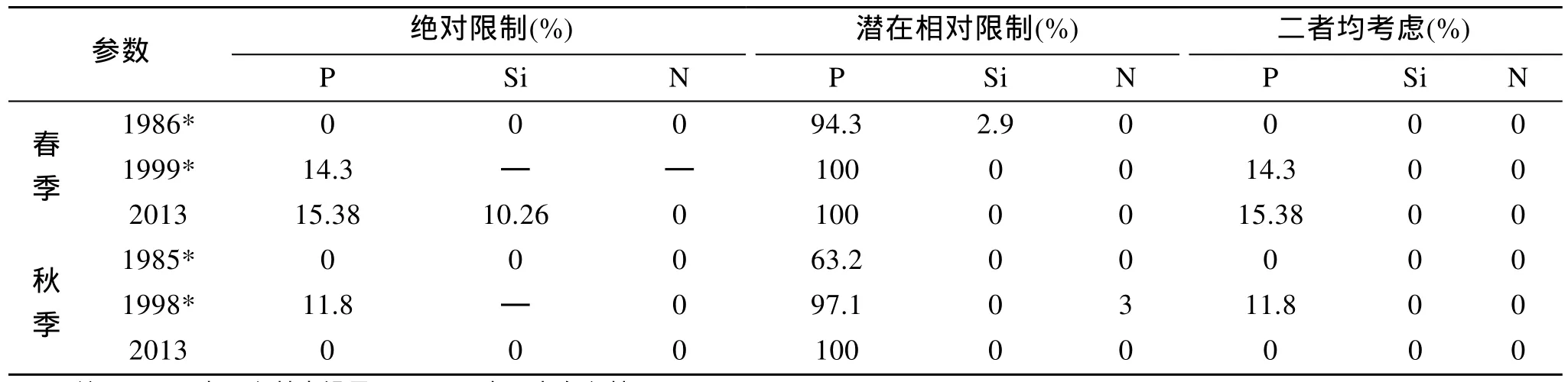
表3 长江口不同年份春、秋季营养盐限制的出现频率Tab.3 The probability of nutrient limitations in the Yangtze River Estuary in spring and autumn of different years
综合考虑营养盐绝对限制和潜在相对限制后,得出调查区域春季存在 P O34--P限制区域, 约占总站位数的 15.38%, 且主要位于叶绿素高值区, 因硅藻的生长使海水中 N/P升高, 并伴随消耗大量的SiO32--Si, 故导致了调查区域较高的 N/P和较低的SiO32--Si、 P O34--P浓度。此外, 低 N/P和高浓度PO34--P有利于硅藻的生长, 而高 N/P和低浓度PO34--P有利于甲藻的生长, 同时PO34--P的相对不足和 DIN/P比例的增大是引起该海域浮游植物群落结构由硅藻向甲藻演替的条件[4]。因此调查海区PO34--P限制区域内硅藻的生长将受到PO34--P的限制, 并且还有可能引起甲藻的大量繁殖。
3 结论
(1) 春季、秋季 DIN的主要形态均为NO3--N,NO3--N、NO-2-N浓度春季高于秋季, NH+4-N则相反。口门内、外PO34--P、SiO32--Si浓度均为秋季高春季低, P O34--P秋季垂直分布均匀, 春季分层明显。NO3--N、 P O34--P、 S iO32--Si浓度从近岸向外海逐渐降低, NO-2-N、NH+4-N分布规律不明显。
(2) 春、秋季长江口营养盐结构均处于高DIN/P状态, 春季Si/DIN大部分小于1, 秋季Si/DIN几乎全部大于1。春季硅藻的局部大量生长使DIN/P值异常升高和 Si/DIN值异常降低, 而秋季存在低 DIN/P和高Si/DIN的分布。
(3) 春、秋季均为PO34--P潜在相对限制, 春季存在局部PO34--P和SiO32--Si的绝对限制, 当同时考虑绝对限制和潜在相对限制时, 春季 15.38%站位受PO34--P限制, 限制情况较20世纪90年代更突出。
[1] 赵卫红, 王江涛.大气湿沉降对营养盐向长江口输入及水域富营养化的影响[J].海洋环境科学, 2007,26(3): 208-216.
[2] 王丽莎, 石晓勇, 祝陈坚.春季长江口邻近海域营养盐分析特征及污染状况研究[J].海洋环境科学,2008 , 27(5): 467-469.
[3] Li M, Xu K, Watanabe M, et al.Long-term variations in dissolved silicate, nitrogen, and phosphorus flux from the Yangtze River into the East China Sea and impacts on estuarine ecosystem [J].Estuarine, Coastal and Shelf Science, 2007, 71(1): 3-12.
[4] 王江涛, 曹婧.长江口海域近 50年来营养盐的变化及其对浮游植物群落演替的影响[J].海洋环境科学,2012, 31(3): 310-315.
[5] 王保栋, 战闰, 臧家业.长江口及邻近海域营养盐的分布特征和输送途径[J].海洋学报, 2002, 24 (1):53-58.
[6] 王奎, 陈建芳, 金海燕, 等.长江口及邻近海域营养盐四季分布特征[J].海洋学研究, 2011, 24(3): 18-35.
[7] 石小勇, 王修林, 韩秀荣, 等.长江口邻近海域营养盐分布特征及其控制过程的初步研究[J].应用生态学报, 2003, 14(7): 1086-1092.
[8] 沈志良, 陆家平, 刘兴俊, 等.长江口区营养盐的分布特征及三峡工程对其影响[C]//中国科学院海洋研究所.海洋科学集刊.北京: 科学出版社,1992.109-129.
[9] 沈志良, 古堂秀.长江口的水化学环境[C]//罗秉征,沈焕庭.三峡工程与河口生态环境.北京: 科学出版社, 1994.141-154.
[10] 潘胜军, 沈志良.长江口及其邻近水域硅酸盐的分布变化特征[J].海洋科学集刊, 2009, 49: 10-18.
[11] 张正斌, 顾宏堪, 刘莲生, 等.海洋化学[M].上海:科学技术出版社, 1984.276-281.
[12] Sierra J.Nitrogen mineralization and nitrification in a tropical soil: effects of fluctuating temperature conditions [J].Soil Biology and Biochemistry, 2002,34(9): 1219-1226.
[13] Zhang J, Zhang Z F, Liu S M, et al.Human impacts onthe large world rivers: Would the Changjiang (Yangtze River) be an illustration?[J].Global Biogeochemical Cycles, 1999, 13(4): 1099-1105.
[14] Zhang J, Ren J L, Liu S M, et al.Dissolved aluminum and silica in the Changjiang (Yangtze River): Impact of weathering in subcontinental scale [J].Global Biogeochemical Cycles, 2003, 17(3): 1-11.
[15] Wang B.Cultural eutrophication in the Changjiang(Yangtze River) plume: History and perspective [J].Estuarine, Coastal and Shelf Science, 2006, 69(3):471-477.
[16] 张平.长江口营养盐结构变化研究[D].青岛, 中国科学院海洋研究所, 2001: 18-22.
[17] 王修林, 孙霞, 韩秀荣, 等.2002年春夏季东海赤潮高发区营养盐结构及分布特征的比较[J].海洋与湖沼, 2004, 35(4): 323-331.
[18] Zhou M J, Shen Z L, Yu R C.Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River [J].Continental Shelf Research, 2008, 28(12): 1483-1489.
[19] Wang B, Wang X, Zhan R.Nutrient conditions in the Yellow Sea and the East China Sea[J].Estuarine,Coastal and Shelf Science, 2003, 58(1): 127-136.
[20] 王奎, 陈建芳, 金海燕, 等.长江口及邻近海区营养盐结构与限制[J].海洋学报, 2013, 35(3): 128-136.
[21] Redfield A C, Ketchum B H, Richards F A.The influence of organisms on the composition of sea water[J].The Sea, 1963: 26-77.
[22] Brzezinski M A.The Si: C: N ratio of marine diatoms interspecific variability and the effect of some environmental variables [J].Journal of Phycology,1985, 21(3): 347-357.
[23] Justic D, Rabalais N N, Turner R E.Stoichiometric nutrient balance and origin of coastal eutrophication[J].Marine Pollution Bulletin, 1995, 30(1): 41-46.
[24] Wong G T F, Gong G C, Liu K K, et al.‘Excess Nitrate’in the East China Sea [J].Estuarine, Coastal and Shelf Science, 1998, 46(3): 411-418.
[25] 张均顺, 沈志良.胶州湾营养盐结构变化的研究[J].海洋与湖沼, 1997, 28(5): 529-535.
[26] Nelson D M, Brzezinski M A.Kinetics of silicic acid uptake by natural diatom assemblages in two Gulf Stream warm-core rings [J].Marine Ecology Progress Series, 1990, 62(3): 283-292.
[27] Fisher T R.Nutrient limitation of phytoplankton in Chesapeake Bay [J].Marine Ecology Progress Series,1992, 82: 51-63.
[28] 李京.东海赤潮高发区营养盐结构及对浮游植物优势种演替的作用研究[D].青岛: 中国海洋大学, 2008:15-32.
[29] 杨登峰, 赵卫红, 李金涛, 等.磷酸盐对中肋骨条藻生长的影响[J].海洋科学集刊, 2004, 46: 165-172.
[30] 李云, 李道季, 唐静亮, 等.长江口及毗邻海域浮游植物的分布与变化[J].环境科学, 2007, 28(4): 719-729.
猜你喜欢
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
海洋通报(2020年2期)2020-09-04
黑龙江水利科技(2020年8期)2020-01-12
应用海洋学学报(2015年3期)2015-11-22
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
棋艺(2001年9期)2001-07-17
棋艺(2001年11期)2001-05-21
棋艺(2001年11期)2001-05-21
