
IL-22重组慢病毒促进肝细胞再生的作用研究*
2015-03-18 01:46张红宾罗红春曾爱中辛小娟
重庆医学 2015年25期
张红宾,罗红春,曾爱中,辛小娟
(1.重庆医科大学附属第一医院:1.血液科;2.感染科 400016)
肝衰竭患者病死率高,积极深入研究促肝细胞的再生机制,探索新的治疗方法是征服这一严重疾病的关键措施。近年来,学者们发现一种新的细胞因子IL-22,其在调节细胞增生、分化、凋亡的过程中发挥重要的作用,并且具有抗感染,诱导急性炎症介质反应,抵御及修复损伤等作用[1-5]。研究发现IL-22受体由2个跨膜的异二聚体IL-22R1和IL-10R2组成。IL-10R2分布广泛,几乎在所有细胞表面均有表达,而IL-22R1的分布具有一定选择性,主要分布于尿路黏膜上皮细胞、消化道和呼吸道黏膜上皮细胞[6-7]。Brand等[8]报道,在肝细胞中IL-22R1和IL-10R2均有分布。为进一步证实IL-22对肝细胞的作用,本课题组拟采用IL-22重组慢病毒感染人正常肝细胞L02,同时设置感染空慢病毒及未感染对照。进行体外培养后,RT-PCT检测L02细胞中结合珠蛋白 mRNA的表达,MMT法检测L02细胞增殖活性,免疫细胞化学法检测L02细胞增殖核抗原(PCNA)的表达。以此评估IL-22重组慢病毒对肝细胞再生的影响,为后续进行的体内实验研究IL-22/REG生物轴与肝脏再生的关系提供细胞实验数据。
1 材料与方法
1.1 材料 IL-22重组慢病毒由本院血液科实验室构建并保存。L02细胞由重庆医科大学病毒性肝炎研究所提供。DNA maker DL2000和PrimerStar PCR试剂盒购自大连宝生物Takara公司。胎牛血清和DMEM培养基购自Hyclone公司。MTT购自美国Sigma公司。鼠抗PCNA单克隆抗体和免疫细胞化学试剂盒购自北京中杉金桥生物技术有限公司。
1.2 方法
1.2.1 IL-22重组慢病毒感染LD2细胞感染效率及IL-22表达测定 调整正常人肝细胞L02密度为2×105个/mL,加入24孔板中培养,待细胞生长至60%融合时进行感染。感染后1、3、5d,荧光显微镜下观察感染效率,并采用ELISA检测感染IL-22重组慢病毒后L02细胞中IL-22的表达情况。
1.2.2 IL-22重组慢病毒对人正常肝细胞L02中结合珠蛋白mRNA表达的影响 将L02细胞接种于24孔板中,接种细胞数为5×105个/孔。设3个实验组:感染IL-22重组慢病毒组(A组)、感染空慢病毒组(B组)、未感染组(C组)。每组设3个复孔,加入DMED细胞培养液(含10%胎牛血清)进行培养。待细胞生长至70%融合时,将IL-22重组慢病毒及空慢病毒储存液稀释100倍后,分别感染A组、B组。C组仍然使用DMED细胞培养液进行培养。在37℃,5%CO2的孵箱中培养48h后,收集各组细胞提取总RNA。
采用RT-PCR检测结合珠蛋白mRNA的表达。设计引物分别为:结合珠蛋白上游引物:5′-TGG CTG CTG ACC ACG GCT AA-3′,下游 引物:5′-CGC ATC GCC ATA GCA GGT GT-3′;内参β-actin上游引物:5′-CAA AGA CCT GTA CGC CAA CA-3′;下游引物:5′-GAA GCA TTT GCG GTG GAC-3′。反应条件:95℃5min;95℃ 1min,55℃ 45s,72℃ 45 s,共30个反应体系;72℃7min。取PCR扩增产物5μL,加6×Buffer 1μL,行1%琼脂糖凝胶电泳。在凝胶成像仪中电泳结果进行分析。
1.2.3 MTT法检测IL-22重组慢病毒对L02细胞增殖的影响 实验分组同前。L02细胞以5×103/孔的密度接种于96孔板,每组设定3个复孔。将96孔板放入37℃,5%CO2的孵箱中培养,分别于感染后2、3、4d后弃上清液,加入MTT溶液20μL,无酚红DMED细胞培养液180μL,继续孵箱中培养4h后检测各孔吸光度值。
1.2.4 免疫细胞化学法检测IL-22重组慢病毒对L02细胞PCNA的影响 L02细胞以5×105个/孔的密度接种于6孔板,放入37℃,5%CO2的孵箱中培养24h。实验分组同前,感染重组慢病毒1d后,进行免疫细胞化学检测。弃上清液后,用0.01mol PBS漂洗3次,每次5min。4%多聚甲醛固定30min,再用0.01mol PBS漂洗3次,每次5min。3%H2O2封闭30min,0.01mol PBS漂洗3次。山羊血清室温孵育10 min。去血清,加鼠抗增殖细胞核抗原PCNA单克隆抗体1∶500(一抗),置湿盒内4℃过夜。MPBS漂洗3次,每次5min。加羊抗小鼠抗体(二抗)。置湿盒内37℃孵育15min。加辣根过氧化物酶标记链酶卵白素(S-A/HRP)。置湿盒内37℃,15 min。再用0.01mol PBS漂洗3次,DAB镜下显色5min,光镜下观察图像。每个孔随机计数5个视野,每个高倍镜视野(×400)计数100个细胞,计算呈阳性染色的细胞百分比。
1.3 统计学处理 采用SPSS17.0统计软件进行数据分析。计量资料以±s表示,采用单因素方差分析及两因素析因设计资料的方差分析,以P<0.05为差异有统计学意义。
2 结 果
2.1 IL-22重组慢病毒感染L02细胞的效率及IL-22表达情况 荧光显微镜下观测,IL-22重组慢病毒感染L02细胞1、3、5d的效率分别是(26.21±2.73)%、(90.12±3.45)%、(61.21±2.48)%,见图1。感染后第3天的感染效率明显高于第1天和第5天(P<0.05)。同时ELISA检测感染后第3天,IL-22在L02细胞中的表达(7.81±0.22)μg/mL显著高于感染后第1天(1.01±0.18)μg/mL和第5天(4.29±0.14)μg/mL(P<0.05)。
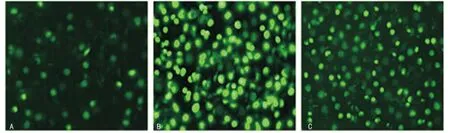
图1 荧光显微镜下观察IL-22重组慢病毒对L02细胞的感染效率
2.2 IL-22重组慢病毒促进正常人肝细胞L02结合珠蛋白mRNA的表达 L02细胞感染重组慢病毒2d后提取总RNA,行RT-PCR扩增后,1%琼脂糖凝胶电泳分析显示,3个实验组的L02细胞均有β-actin及结合珠蛋白的表达,其条带大小分别为255、480bp,和实验预期结果一致,见图2。L02细胞结合珠蛋白mRNA的半定量分析以结合珠蛋白条带/βactin条带的光密度值表示,见图3。经统计学分析,A组L02细胞结合珠蛋白mRNA的表达明显高于B组及C组(P<0.01),B组和C组之间,差异无统计学意义(P>0.05)。
2.3 MTT法检测出IL-22重组慢病毒对L02细胞有明显的促再生作用 MTT法检测各孔吸光度值显示,A组L02细胞各时间点的再生活性均明显高于B组及C组(P<0.05)。而B组和C组L02细胞各时间点的再生活性差异无统计学意义(P>0.05)。IL-22重组慢病毒对L02细胞的促再生作用于感染3d最强,感染4d后有所减弱。见图4。
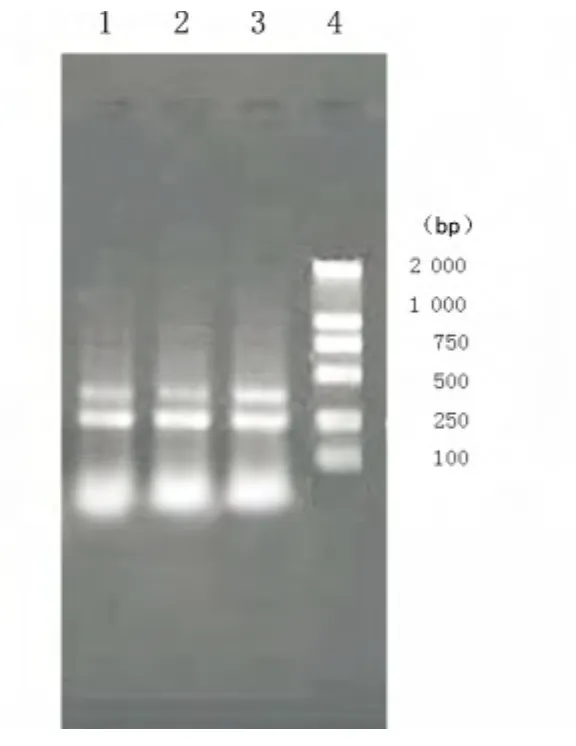
图2 结合珠蛋白凝胶电泳分析结果
2.4 免疫细胞学检测出IL-22重组慢病毒能促L02细胞PCNA表达 光镜下,细胞质着色,而细胞核无色或细胞质细胞核均无色记为PCNA阴性细胞,细胞核染为棕黄色记为PC-NA阳性细胞。A组有较多PCNA阳性细胞,细胞核染色深,可见呈双核或分叶状,且细胞体积大,为再生肝细胞。B组仅有少数几个PCNA阳性细胞。C组几乎无PCNA阳性细胞,见图5。计算各组标记指数(%),经统计分析后结果示:与B组及C组相比,A组显著促进PCNA的表达(P<0.01)。B组和C组差异无统计学意义(P>0.05),见图6。

图3 结合珠蛋白条带与β-actin条带的光密度比值

图4 MTT法检测IL-22重组慢病毒对L02细胞促再生作用

图5 免疫细胞化学检测各处理组L02细胞PCNA表达情况(×400)
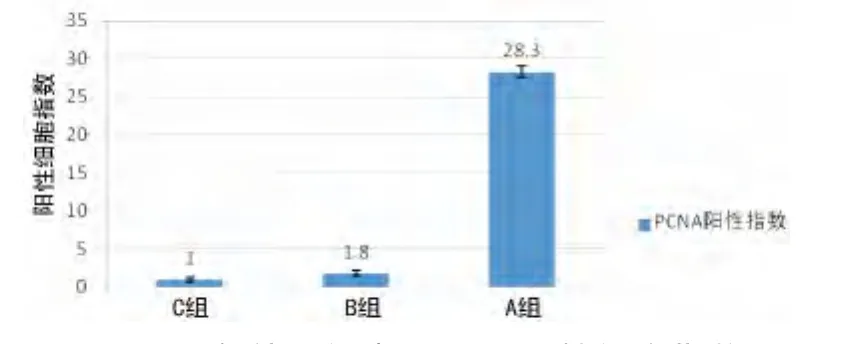
图6 各处理组中PCNA阳性细胞指数
3 讨 论
目前肝衰竭的治疗,是临床上非常棘手的问题。因为该病肝细胞坏死多,再生能力差,缺乏特效治疗方法,以致病死率高。因此,积极深入研究促肝细胞再生的相关机制,探索新的促肝细胞再生的有效治疗方法势在必行。据研究发现,人和哺乳动物的肝脏具有强大的再生功能,2/3肝切除后的动物在10~15d左右残存肝脏可恢复到术前体积。肝细胞再生涉及到复杂的网络信号通路:一是细胞因子通路,与肝细胞再生启动有关;二是生长因子通路,与细胞周期进程有关[9]。现在已经证实的能促进肝细胞再生的相关因子包括IL-6、肿瘤坏死因子α(TNF-α)、肝细胞生长因子(HGF)、转化生长因子α(TGF-α)、表皮细胞生长因子(EGF)、干细胞因子等[9-11]。2000年,有研究者发现了一种新的细胞因子IL-22,属于IL-10家族成员,由6个螺旋构成,反平行排列形成束状单体蛋白,分子量为16.7~17.0×103[12]。其在调节细胞再生、分化、凋亡、抗感染、诱导急性炎性介质反应、抵御及修复损伤等方面起重要作用。研究者们发现在肝脏有IL-22受体分布,并对IL-22在肝脏损伤修复中的作用进行了初步研究[13-14],认为IL-22有利于肝脏的再生。为进一步证实IL-22与肝脏再生修复的关系,本课题组进行了上述实验。
将IL-22重组慢病毒感染正常肝细胞L02后,采用RTPCR检测结合珠蛋白mRNA的表达量。结合珠蛋白绝大部分在肝脏中合成,是一种急性时相蛋白,具有重要的抗炎作用。实验结果发现IL-22重组慢病毒能大幅度刺激结合珠蛋白mRNA的表达,而感染空慢病毒组和未感染组中结合珠蛋白mRNA的表达量差异无统计学意义(P>0.05)。MTT法检测发现感染IL-22重组慢病毒组在感染后2、3、4d,L02细胞的再生活性均明显高于感染空慢病毒组及未感染组。但IL-22对L02细胞的促再生活性在感染后3d最强,之后有所减弱。这可能与细胞本身的老化有关。还采用细胞免疫化学方法,以L02细胞为对象,研究IL-22重组慢病毒对L02细胞的PCNA表达的影响,从另一个方面探讨IL-22对肝细胞再生的促进作用。PCNA是存在于细胞核内的一种蛋白质,相对分子质量约为36KD。其表达的高峰在细胞周期的G/S期,是细胞DNA合成所必需的蛋白质,是判断再生细胞生物活性及所处生长状态的重要指标。本研究结果显示,感染IL-22重组慢病毒组有较多PCNA阳性细胞,经统计分析该组PCNA阳性细胞显著高于感染空慢病毒组及未感染组。虽然感染空慢病毒组也有少数PCNA阳性细胞,但与未感染组相比,差异无统计学意义(P>0.05),故认为慢病毒本身不会引起PCNA大量表达。
综上所述,IL-22可以促进L02细胞结合珠蛋白mRNA及PCNA的表达,并能增强L02细胞的再生活性,故认为IL-22可能对肝细胞再生有促进作用。这一实验结果为本课题组后续进行的体内实验研究IL-22/REG生物轴与肝脏再生的关系提供了细胞实验数据。
[1]Dumoutier LJ.IL-TIF,a novel cytokine structurally related to IL-10and inducible by IL-9[J].J Immunol,2000,164(4):1814-1819.
[2]Wolk K,Kunz S,Witte E,et al.IL-22increases the innate immunity of tissues[J].Immunity,2004,21(2):241-254.
[3]Wolk K,Sabat R.Interleukin-22:a novel T-and NK-cell derived cytokine that regulates the biology of tissue cells[J].Cytokine Growth Factor Rev,2006,17(5):367-380.
[4]Andoh A,Zhang Z,Inatomi O,et al.Interleukin-22,a member of the IL-10subfamily,induces inflammatory responses in colonic subepithelial myofibroblasts[J].Gastroenterology,2005,129(3):969-984.
[5]Wolk K,Witte E,Wallace E,et al.IL-22regulates the expression of genes responsible for antimicrobial defense,cellular differentiation,and mobility in keratinocytes:a potential role in psoriasis[J].Eur J Immunol,2006,36(5):1309-1323.
[6]Dumoutier L,Lejeune D,Colau D,et al.Cloning and characterization of IL-22binding protein,a natural antagonist of IL-10-related T cell-derived inducible factor/IL-22[J].J Immunol,2001,166(12):7090-7095.
[7]Weiss B,Wolk K,Grünberg BH,et al.Cloning of murine IL-22receptor alpha 2and comparison with its human counterpart[J].Genes Immun,2004,5(5):330-336.
[8]Brand S,Dambacher J,Beigel F,et al.IL-22-mediated liver cell regeneration is abrogated by SOCS-1/3overexpression in vitro[J].Am J Physiol Gastrointest Liver Physiol,2007,292(4):G1019-1028.
[9]Fausto N,Riehle KJ.Mechanisms of liver regeneration and their clinical implications[J].J Hepatobiliary Pancreat Surg,2005,12(3):181-189.
[10]Koniaris LG,Mckillop IH,Schwartz SI,et al.Liver regeneration[J].J Am Coll Surg,2003,197:634-659.
[11]Ren XD,Hu B,Colletti L.Stem cell factor and its receptor,c-kit,are important for hepatocyte proliferation in wild-type and tumor necrosis factor receptor-1knockout mice after 70%hepatectomy[J].Surgery,2008,143(6):790-802.
[12]Dumoutier L,Van Roost E,Ameye G,et al.IL-TIF/IL-22:genomic organization and mapping of the human and mouse genes[J].Genes Immun,2000,1(8):488-494.
[13]Ren X,Hu B,Colletti LM.IL-22is involved in liver regeneration after hepatectomy[J].Am J Physiol Gastrointest Liver Physiol,2010,298(1):G74-80.
[14]Ki SH,Park O,Zheng M,et al.Interleukin-22treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding:role of signal transducer and activator of transcription 3[J].Hepatology,2010,52(4):1291-1300.
猜你喜欢
检验医学与临床(2022年18期)2022-09-27
现代诊断与治疗(2022年7期)2022-08-13
转化医学杂志(2022年1期)2022-03-07
医药与保健(2021年4期)2021-04-07
野生动物学报(2020年1期)2020-02-21
中华神经外科疾病研究杂志(2018年4期)2018-08-20
检验医学与临床(2018年13期)2018-07-12
中华肺部疾病杂志(电子版)(2018年6期)2018-03-11
中国高原医学与生物学杂志(2017年4期)2017-03-08
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
