
40周龄母鸡体尺性状的主基因+多基因混合遗传分析
2015-03-15 12:32王克华窦套存沈曼曼
江苏农业学报 2015年5期
马 猛, 王克华, 曲 亮, 窦套存, 沈曼曼
(1.江苏省家禽科学研究所,江苏 扬州 225003;2.扬州翔龙禽业发展有限公司,江苏 扬州 225200)
鸡的体尺性状与众多重要的经济性状关系密切。目前,对鸡体尺的研究多集中在其与屠宰性能、生长发育规律的关系上[1-3]。与其他数量性状一样,影响鸡体尺性状的因素较多,其中遗传因素和环境是主要影响因素。对鸡体尺性状的非遗传因素影响已有较多报道[4-6]。但是,目前关于体尺性状的分子遗传机理研究较少。以往多采用传统的方法进行体尺性状的分子遗传研究,然而传统的方法有一定的局限性,不能解析单个基因座位的遗传效应[7]。盖钧镒等[8]、Zhang 等[9]、Wang 等[10]提出的主基因 +多基因混合遗传分析方法不仅能够鉴别主基因,而且对多基因也可以进行检测,并估计出相应的遗传参数。本研究以绿壳蛋鸡黑羽纯系和白来航鸡资源群体为材料,应用主基因+多基因混合遗传模型分析软件对40周龄母鸡的体尺性状进行研究,探讨鸡40周龄体尺性状的遗传规律,确定最适模型。
1 材料与方法
1.1 试验材料
试验以黑羽绿壳蛋鸡纯系为亲本P1,以白来航鸡为亲本P2,其中P1共109 只,P2共78 只,F1(P1♂ ×P2♀、P2♂×P1♀)代564 只,F2[(P1♂ ×P2♀)♂ ×(P1♂ ×P2♀)♀、(P2♂ ×P1♀)♂ ×(P2♂ ×P1♀)♀]代1 915只。试验动物均饲养于扬州翔龙禽业发展有限公司,饲养管理条件一致,试验过程中每只鸡均单笼饲养。
1.2 测定内容与方法
测定每个世代所有母鸡在40周龄时的体尺性状,包括胫围、胫长、龙骨长、体斜长、胸角和胸宽6项指标。其中胫长、龙骨长、体斜长、胸角和胸宽的测定均按照《家禽生产性能名词术语和度量统计方法》[11]进行测定,胫围的测定方法为用细线绕鸡左胫中间部位3圈,然后用直尺量取3圈的总周长,除以3即为每只鸡的胫围。
1.3 数据的处理
试验所有数据均采用Excel 2003进行统计,用SPSS17.0进行正反交群体间的t检验,用南京农业大学研发的SEA-G4F2软件包进行主基因+多基因混合遗传模型分析。
数据进行分析前对F1代和F2代的数据进行正反交之间的比较,如果正反交之间的差异显著,则需要消除正反交效应。
正交效应=正交群体平均值-群体平均值
反交效应=反交群体平均值-群体平均值
用于分离分析的数据=原数据-正反交效应
1.4 遗传模型简介
利用P1、P2、F1和F23个世代进行分离分析的遗传模型共有5大类24个遗传模型,其具体对应的解释见表1。
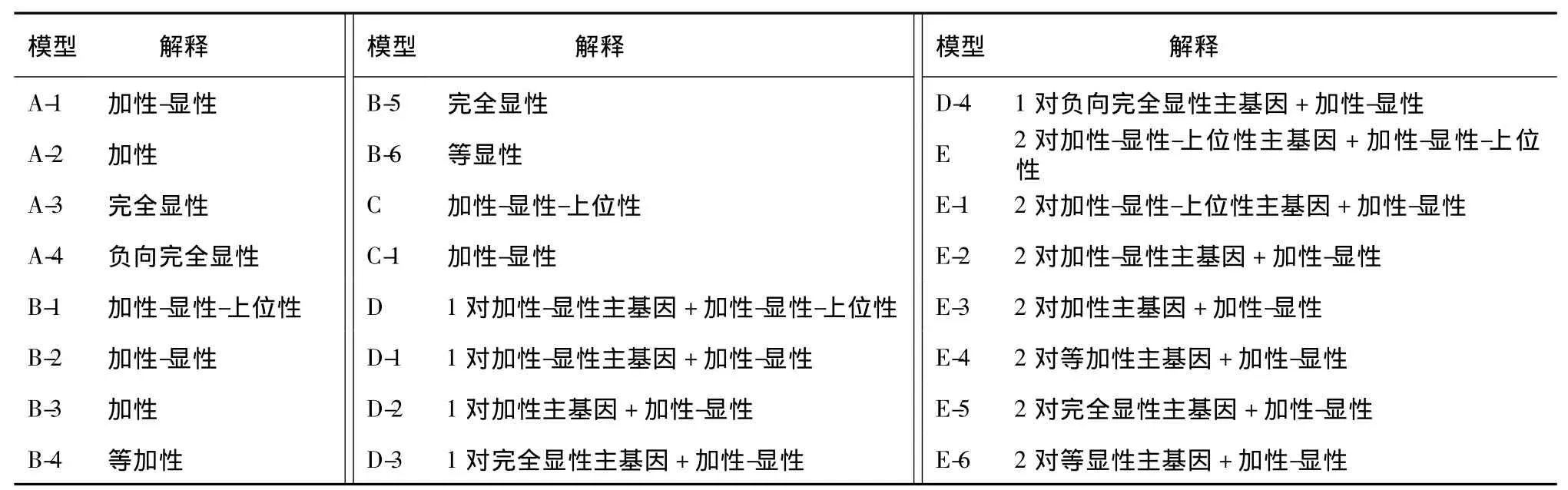
表1 各个遗传模型对应的解释Table 1 Explain ation for of each genetic model
1.5 统计与分析
应用盖钧镒等[8]提出的主基因+多基因混合遗传模型来进行分离分析,通过极大似然法和IECM算法对混合分布中的有关成分分布参数做出估计,然后对24个模型中AIC(Akaike,s information criterion)值进行比较,选择其中AIC值较小的相对应的模型进行适合性检验,然后确定出最适模型,根据最适模型的分析结果,估计其相应的一阶遗传参数和二阶遗传参数。具体的最适模型的选择和遗传参数估计参照文献[8]。适合性检验共有5个统计量,即均匀性检验检验 nW2和Kolmigorov 检验 Dn。

2 结果与分析
2.1 40周龄母鸡体尺性状的表型特征值及F2代的次数分布
6个体尺性状的表型特征值见表2,由表2可知,每个性状的F2代的极差均大于P1、P2和F1代,说明每个性状在F2代都有较好的分离。6个体尺性状中除胫长F1代的均值介于两个亲本之间外,其他5个性状的F1均值均高于或低于亲本,说明这些性状均发生了超亲遗传的现象。6个性状F2代的峰度均大于0,说明各个性状的F2代分布均比正态分布要陡峭,而6个性状中除了胫围F2代的偏度大于0外,其他5个性状的偏度均小于0,说明胫围F2代的分布右边较为分散,其他性状的F2代分布则左边比较分散,且龙骨长F2代的分散程度更高。
图1为40周龄母鸡6个体尺性状F2代的次数分布,由图1可知,胫围F2代的分布呈现单峰的偏态分布,胫长F2代的次数分布呈现单峰分布,其他性状F2代的次数分布则呈现出多峰分布,这也说明每个性状在F2代都出现了较好的分离,且每个性状均属于主基因控制的性状。

表2 40周龄母鸡6个体尺性状每个世代对应的表型特征值Table 2 Phenotypic value of six body size traits in each generation

图1 40周龄母鸡体尺性状F2代的次数分布Fig.1 Frequency distribution of body size traits in F2 at 40-week old
2.2 40周龄母鸡6个体尺性状的最适模型的确定及适合性检验
40周龄母鸡6个体尺性状的各个模型所对应的AIC值见表3,根据候选模型的选择标准,在24个模型中选取AIC值较小的模型作为候选模型进行适合性检验。由表3可知,胫围、胫长、胸角和胸宽的24个模型中E和E-1的AIC值较小,因此选模型E和E-1作为这4个性状的候选模型,龙骨长和体斜长的24个模型中模型E的AIC值均要远远小于其他模型,因此模型E作为龙骨长和体斜长的候选模型。最适模型的确定是根据适合性检验的统计量显著的个数进行,选择达到显著性的统计量个数最少的模型作为最适模型,若候选模型的显著统计量个数相同,则选取AIC值较小的作为最适模型。6个性状的候选模型的适合性检验统计量达到显著性的个数见表4。由表4可知,胫围、胫长、胸角和胸宽的候选模型E和E-1的检验统计量达到显著的个数均相等,因此选择AIC值较小的E-1作为4个性状的最适模型,即2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型。龙骨长和体斜长的候选模型检验统计量显著个数分别为1和6,最适模型均为E模型,即2对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传模型。最适模型的适合性检验见表5。

表3 40周龄母鸡6个体尺性状各个模型所对应的AIC值Table 3 AIC values of six body size traits of 40-year-old hens by each-model

表4 候选模型所对应的适合性检验的统计量的显著个数Table 4 Significant numbers of statistics in adaption test for candidate models

表5 最适模型的适合性检验Table 5 Test of adaption for the best model
2.3 最适模型的遗传参数估计
40周龄母鸡6个性状的最适模型的遗传参数估计见表6。由表6可知,在一阶遗传参数的估计中,胫围、胫长、胸角和胸宽的2对主基因的加性效应均为正向效应,龙骨长和体斜长的2对主基因的加性效应为负向效应,6个性状中除了胸角的2对主基因的显性效应为负向效应外,其他性状的主基因的显性效应均为正向效应。6个性状中除了体斜长外,其他5个性状的主基因的加性效应和显性效应基本可以与加加效应、显显效应和加显效应、显加效应相互抵消,体斜长的|da|>|db|,ha>hb,说明第1对主基因的加性效应和显性效应均大于第2对主基因。在二阶遗传参数的估计中,胫围、胫长、胸角和胸宽的群体方差分别为0.216、0.988、72.356和0.849,主基因所对应的方差分别为0.131、0.605、40.245和0.504,主基因的遗传率分别为 60.89%、61.24%、55.62%和59.43%,龙骨长和体斜长的群体方差为0.461和0.919,主基因的方差为0.076和0.283,主基因遗传率分别为16.52%和30.76%,龙骨长对应的多基因的方差为0.033,多基因遗传率为7.20%。

表6 40周龄母鸡6个性状最适模型的遗传参数估计Table 6 Estimations of genetic parameters of best model for each trait
3 讨论
王春娥等[12]的研究结果表明,运用主基因+多基因混合遗传分析软件检测到的主基因的对数与用QTL定位检测出来的主基因对数是一致的。盖均镒等[13]认为用单个世代的分析效果没有多世代的分析效果好,原因是加性遗传参数体现在两个纯合的亲本均值差异上,而显性遗传参数在亲本与F1代的均值差异中就可以体现出来。目前,对主基因+多基因混合遗传分析软件的应用大多集中在植物数量性状的研究上[14-17],动物数量性状上的研究则有王克华[18]、曲亮[19-20]等应用主基因 +多基因混合遗传模型对鸡12周龄体质量、鸡40周龄蛋质量、冠长、冠高和冠厚的研究,确定鸡12周龄体质量的最适模型为E-6模型,鸡40周龄蛋质量、冠长、冠高和冠厚的最适模型均为E模型。
本研究结果表明,40周龄母鸡体尺性状中的胫围、胫长、胸角和胸宽的最适模型均为E-1模型,即2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型,龙骨长和体斜长的最适模型均为E模型,即2对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传模型,胫围、胫长和胸宽的2对主基因的加性效应值和显性效应值均为正值,龙骨长和体斜长的2对主基因的加性效应值为负值,显性效应值为正值,说明胫围、胫长、胸角和胸宽越大,而龙骨长和体斜长越小的个体对后代的影响越大。本研究结果显示40周龄母鸡的胫围、胫长、胸角和胸宽的主基因遗传率均高于50%,每个性状的主基因效应均远远大于多基因的遗传效应,因此,在对40周龄母鸡的胫围、胫长、胸角和胸宽等性状进行遗传改良的过程中要注重对主基因的选择,40周龄母鸡的龙骨长和体斜长的主基因与多基因的遗传率之和均小于50%,这说明非遗传因素对这2个性状的影响较大,在后期的选育过程中,要注重改善环境条件,降低非遗传因素对龙骨长和体斜长的影响。
虽然主基因+多基因混合遗传模型的应用比较广泛,但是其本身还存在一定的局限性,只能够将主基因的对数为1~3的检测出来。在动物数量性状上开展主基因+多基因的应用,不仅可以丰富动物数量性状的研究方法,而且可以较好地了解性状本身的内在遗传规律,能为与主基因紧密连锁的分子标记的发掘和分子标记辅助选择奠定一定的理论基础。
[1] 李俊营,陈丽园,詹 凯,等.黄山黑鸡体尺、屠宰性能和肉品质测定及相关分析[J].中国家禽,2014,36(14):52-54.
[2] 祝碧琴,曾 涛,李进军,等.白耳黄鸡体尺及屠宰性能的相关性分析[J].浙江农业科学,2012(3):403-404.
[3] 李乃宾,杨芬霞,杜炳旺,等.珍禽贵妃鸡体尺性状与屠宰性能的相关分析[J].南方农业学报,2014,45(7):1281-1285.
[4] 童海兵,王克华,陆俊贤,等.鸡种,日粮能量和日粮蛋白质对体尺性状的影响[J].中国家禽,2004,8(1):96-99.
[5] 孙桂荣,康相涛,李国喜,等.不同饲养方式对卢氏鸡体尺和屠体性状的影响[J].饲料广角,2008(16):40-42.
[6] 潘 文,杜炳旺,王润莲,等.饲粮钙水平对贵妃鸡生长性能、体尺性状及趾骨特性的影响[J].动物营养学报,2014,24(6):1660-1667.
[7] 陈学军,方 荣,周坤华,等.辣椒果实性状主基因+多基因遗传分析[J].西北植物学报,2012,32(2):246-251.
[8] 盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003.
[9] ZHANG Y M,GAI J Y.The EIM algorithm in the joint segregation analysis of quantitative traits[J].Genetical Research,2003,81(2):157-163.
[10] WANG J,FODLIED D W,COOPER M,et al.Power of the joint segregation analysis method for testing mixed major-gene and polygene inheritance models of quantitative traits[J],Theoretical and Applied Genetics,2001,103(5):804-816.
[11]NY/T823-2004 家禽生产性能名词术语和度量统计方法[S].
[12]王春娥,盖钧镒,傅三雄,等.大豆豆腐和豆乳得率的遗传分析与 QTL 定位[J].中国农业科学,2008,41(5):1274-1282.
[13]盖钧镒,王建康.利用回交或F2:3家系世代鉴定数量性状主基因-多基因混合遗传模型[J].作物学报,1998,24(4):402-409.
[14]周清元,崔 翠,阴 涛,等.甘蓝型油菜角果长度的主基因+多基因混合遗传模型[J].作物学报,2014,40(8):1493-1500.
[15]李洪戈,余坤江,郭婷婷,等.甘蓝型油菜无花瓣性状的主基因+多基因遗传分析[J].江苏农业学报,2014,30(2):253-258.
[16]李兰周,刘风珍,万勇善,等.花生荚果和籽仁相关性状的主基因+多基因混合遗传模型分析[J].华北农学报,2013,28(5):116-123.
[17]马 娟.玉米主要株型性状的主基因+多基因遗传模型分析[D].新乡:河南科技学院,2012.
[18]王克华,章元明,曲 亮,等.鸡12周龄体重的主基因+多基因混合遗传分析[J].江西农业大学学报,2011,33(1):123-127.
[19]曲 亮,马 猛,王克华,等.鸡40周龄蛋重主基因-多基因混合遗传模型分析[J].中国家禽,2014,36(7):10-16.
[20]曲 亮,王克华,窦套存,等.鸡第二性征的主基因+多基因混合遗传分析[C]//中国畜牧兽医学会,第十六次全国动物遗传育种学术讨论会论文集.扬州:中国畜牧兽医学会,2011:508.
猜你喜欢
动物营养学报(2022年11期)2022-12-08
农业工程学报(2022年13期)2022-10-09
特产研究(2021年6期)2021-12-08
东北农业大学学报(2021年9期)2021-11-11
今日农业(2020年15期)2020-12-15
安徽农业科学(2020年12期)2020-07-14
安徽科技学院学报(2019年4期)2019-10-28
河南农业科学(2019年9期)2019-09-24
农村百事通(2016年6期)2016-05-14
中国畜禽种业(2015年5期)2015-12-02
