
土壤干旱对转基因甘薯光合曲线的响应
2015-03-13 01:27陆燕元马焕成李昊民邓西平
生态学报 2015年7期
陆燕元,马焕成,李昊民,邓西平
1 西南林业大学, 西南生物多样性保育国家林业局重点实验室, 昆明 650224 2 西南林业大学, 云南生物多样性研究院, 昆明 650224 3 黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100
土壤干旱对转基因甘薯光合曲线的响应
陆燕元1,马焕成1,李昊民2,邓西平3,*
1 西南林业大学, 西南生物多样性保育国家林业局重点实验室, 昆明 650224 2 西南林业大学, 云南生物多样性研究院, 昆明 650224 3 黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100
采用盆栽试验的方法,设置适宜水分、中度干旱和重度干旱(土壤田间持水量的75%—80%、60%—65%和35%—40%) 等3个土壤水分条件,以叶片同时转入Cu/Zn SOD和APX基因的甘薯和未转基因对照植株为材料,研究它们在不同水分条件下的光合响应特性。结果表明:在不同的土壤水分条件下,转基因(TS)和未转基因(NS)植株光响应曲线的变化趋势均基本一致,但转基因甘薯叶片的光合速率都高于未转基因植株,尤其是在严重的干旱胁迫下两者的差异最为显著,表明同时转入的Cu/Zn SOD和APX抗氧化酶基因有利于甘薯在干旱逆境下维持较高的光合能力。正常的水分水平及中度干旱胁迫下,TS和NS植株叶片的Pmax无明显差别,但在严重干旱条件下TS叶片的Pmax显著高于NS。在本试验中,与适宜水分处理相比,NS植株叶片的表观量子效率(φ)在中度干旱胁迫下就开始显著下降,而TS植株只是轻微下降,到严重干旱时,两者都显著下降,但是TS仍略高于NS植株。随着土壤干旱程度增加,甘薯叶片光补偿点(LCP)增高,光饱和点(LSP)、表观量子效率(φ)、暗呼吸速率(Rd)下降,表明干旱胁迫使气孔关闭,光能利用率降低,同时利用CO2的能力降低,光合产物消耗增加,不利于同化产物的积累,在干旱胁迫条件下,转基因甘薯叶片光补偿点(LCP)、光呼吸增高以及光饱和点下降的幅度均明显小于未转基因植株,这可能是因为转入的抗氧化酶基因提高干旱胁迫下叶片中酶促防御系统的活性,适度缓解了干旱逆境引起的膜脂过氧化伤害。
转基因; 甘薯; 干旱; 光合响应曲线
光合作用是作物生长发育以及产量形成的基础。提高叶片的光合性能是实现作物高产的重要途径[1]。植物的光响应曲线反映的是光合速率随着光照强度的变化特性,是判定植物光合能力的重要指标[2]。植物的光响应曲线不仅受外界条件的影响,更主要受自身遗传特性的影响。已有的研究表明,转基因甘薯在水分胁迫条件下的超氧化物歧化酶(SOD)、抗坏血酸过氧化物酶(APX)及过氧化氢酶(CAT)等抗氧化酶活性显著高于未转基因植株,膜系统受到的损伤较小,表明转入的外源基因有效提高了甘薯抗氧化系统的防御能力,抗旱性增强[3- 6],但是否对植物在干旱等逆境条件下光合特性,尤其是其潜在的光合能力有影响尚未见报道。本试验旨在通过对转基因和未转基因甘薯的光响应曲线及其特征参数的研究,探讨转入的抗氧化酶基因对甘薯在水分处理条件下光响应曲线参数的变化特征及其生理机制,从而为转基因甘薯耐旱性研究提供了理论基础。
1 材料与方法
1.1 供试材料
在逆境诱导型启动子SWPA2诱导下在叶片叶绿体中同时表达Cu/ZnSOD和APX抗氧化酶基因甘薯(IpomoeabatatasLam.)植株(TS)和未转基因植株(NS)。基因的筛选、遗传转化以及再生体系的建立均在韩国生命科学与生物技术研究所完成[7- 8]。随后材料的移栽、胁迫试验均在中国科学院水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室中完成。

图1 表达载体SSA的构建Fig.1 Construction of expression vector SSASWPA2pro: 甘薯过氧化物酶启动子sweet potato peroxidase promoter;TEV: 烟草蚀刻病毒tobacco etch virus;TP: 叶绿体转运缩氨酸chloroplast-targeted transit peptide;SOD: 木薯Cu/Zn SOD cassava Cu/Zn SOD;35Sp: 花椰菜花叶病毒35s启动子CaMV 35s promoter;APX: 豌豆抗坏血酸过氧化物酶pea ascorbate peroxidase;35ster: 花椰菜花叶病毒35s终止子35ster, CaMV 35s terminator
1.2 试验设计及取样方法
试验在陕西杨凌中国科学院水土保持研究所干旱棚进行。试验采用随机区组设计。将在室内培养好、大小基本一致的转基因和未转基因甘薯移栽到装好土的塑料桶(长×宽×高=60 cm×20 cm×40 cm)中。盆栽土用3∶1的耕层土和沙子进行混合,每盆装土12 kg,并施入8.63 g 化肥(2.8 g N∶1.43 g P∶4.4 g K=1∶0.51∶1.57)作为底肥,在每桶土壤表面覆盖50 g 的珍珠岩,防止土壤水分的蒸发。每盆2 株,每个水分处理4 盆。待苗子成活后(移栽30 d后)开始设3个水分处理:充足的水分处理(对照,CK),整个生育期土壤含水量一直维持在田间持水量的75%—80%;中度干旱处理:土壤含水量达到田间持水量的60%—65%;严重干旱:土壤含水量达到田间持水量的35%—40%。到生长后期(移栽121 d后),除对照以外,对中度干旱和重度干旱处理进行复水。试验采用称重的方法控制土壤含水量:即每天下午进行称重,补充消耗的水分,使其一直维持在该水分水平。
取样方法
对处理后的甘薯第4片完全展开叶进行光合曲线测定。
1.3 光合曲线的测定
测定选择在9月份的晴天进行,每天从9:30开始至15:00结束,采用Li- 6400型便携式光合测定系统测定,开放式气路,温度控制在(28±0.5) ℃,空气相对湿度为50%—70%,应用Li- 6400-02B内置式红蓝光光源提供不同的光合有效辐射强度(PAR),光照强度梯度设置为0、40、80、100、200、600、1000、1500、2000 μmol m-2s-1,测定各水分条件下转基因和未转基因植株甘薯叶片Pn。测定Pn前使用500 μmol m-2s-1冷光源诱导10 min,在每个光照强度下平衡60—120 s后开始测定。同时,利用光合仪同步测出气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、相对湿度(RH)等光合参数。测定时每处理随机选3个典型植株,结果取平均值。
1.4 光响应曲线特征参数提取
作物的光响应曲线采用经典的Farquhar模型来拟合,它由Prioul和Chartier首次提出[9],随后得到广泛的应用[9- 15],该模型的理论公式为:
式中,Pn为净光合速率(μmol m-2s-1),I为光量子通量密度即光合有效辐射强度(μmol m-2s-1),φ为表观量子效率,Pmax为光饱和时的最大净光合速率(μmol m-2s-1),Rd为暗呼吸速率(μmol m-2s-1),θ为非直角双曲线的凸度。其中φ与Pmax是描述光合作用光响应特征的参数,φ是光响应曲线的初始斜率,表示植物在光合作用对光的利用效率,而Pmax是衡量叶片光合能力的一个主要参数。
本研究运用SPSS13.0统计软件中的非线性统计分析模块等技术做统计分析,具体步骤参照刘宇锋的方法[16],运用其方程对转基因与未转基因甘薯片的光合速率与光强的关系进行拟合分析。由图2可见,该公式可以较好地模拟光合速率随光强的变化,转基因和未转基因甘薯各水分处理的光响应曲线的决定系数(R)可以有效控制在0.98以上,模拟结果能反映实际情况。利用该公式对测定数据进行拟合,可直接求出转基因和未转基因甘薯叶片的θ、Pmax、Rd等参量。在低光强下,Pn随光合辐射强度的增加呈线性增高。通过对200 μmol m-2s-1以下所采集数据进行直线回归数分析(图3),运用所得出的直线方程与前述中得出的最大光合速率(Pmax)值,可计算出光补偿点(LCP)、光饱和点(LSP)和表观量子效率(φ)等参数。200 μmol m-2s-1以下的线性方程,与非直线双曲线模型计算出的Pmax值和X轴这两条平行直线相交,可得出交点。其与X轴的交点数值即为LCP,而与Pmax的交点在X轴上的数值则为LSP,其中表观量子效率可表示为该直线方程的斜率。
2 结果与分析
2.1 土壤干旱对转基因和未转基因甘薯叶片光响应曲线的影响
不同土壤干旱程度下转基因和未转基因甘薯光合响应曲线响应变化如图4。由该图可以看出,光强PAR在0—400 μmol m-2s-1左右时,净光合速率Pn快速上升,当光强达到400 μmol m-2s-1左右时上升的幅度减弱。进一步分析各处理的光合速率随光强的变化趋势可以发现,转基因和未转基因植株在充足的水分条件下光合响应曲线无明显差异。严重干旱下TS植株叶片的净光合速率明显高于NS,两者差异非常显著;中度干旱条件下TS光合速率略高于NS。

图2 Farquhar模型拟合光合曲线响应Fig.2 Photosynthetic response curve fitting by Farquhar model

图3 0—200 μmol m-2 s-1 Pn-PAR 直线回归Fig.3 Linear regression of 0—200 μmol m-2 s-1 Pn-PAR

图4 不同土壤干旱程度下转基因(TS)和未转基因甘薯(NS)光合响应曲线响应Fig.4 Light response curves of photosynthesis of transferred genes sweet potato (TS) and non- transferred control under different soil moistureTS: 转基因甘薯,NS: 未转基因甘薯,CK: 充足水分处理,MS: 中度水分胁迫,SS: 重度水分胁迫
2.2 土壤干旱对转基因和未转基因甘薯叶片光响应特征参数的影响
在一定环境条件下,叶片的最大光合速率表示了叶片的最大光合能力[17- 18]。由表1可以看出,在水分充足条件下,TS和NS植株的Pmax基本处于同一水平,中度干旱胁迫下都略有下降,TS下降的幅度较小,但与NS的差异不明显。在重度水分胁迫下,TS和NS的最大光合速率均显著下降,其中TS相对于对照降低了28%,而NS则下降了44%,两者差异较大。表观量子效率(φ),是反映植物在弱光下吸收转换和利用光能能力的指标,是光合作用中光能转化效率的指标之一,φ值较高则可解释为叶片具有较高光能转化效率,通常值在0.03—0.07之间,呼吸速率(Rd)则与叶片的生理活性有关[1, 19- 20]。由表1可以看出这两个指标的变化趋势趋同,均为随土壤水分胁迫程度的增加而逐渐降低,但是TS下降的幅度始终小于NS植株。光饱和点和光补偿点分别表示植物利用强光和弱光的能力,两点差距越大说明植物利用光的能力越强。从表1中可知,光饱和点是随土壤胁迫增强而逐渐降低,而光补偿点则呈相反的趋势,说明甘薯在水分胁迫下对光的利用受到限制,但是NS受限制的程度更大些。
3 结论与讨论
光照是影响植物生长和生物量积累的重要环境因子之一。光合作用光响应曲线反映了植物光合速率随光照强度增减的变化规律[18],而由此获得光合作用的最大光合速率、表观量子效率、光饱和点、光补偿点和暗呼吸速率等生理参数,有助于判定植物光合机构运转状况、光合作用能力和光合作用效率及其受环境变化的影响程度,因此光响应曲线的测定是植物光合生理生态学研究的重要手段之一[16, 21]。水分是影响植物生长的主要环境因子之一,干旱胁迫通过各种途径降低植物的光合能力[21]。本课题组之前的研究表明,在甘薯叶片中同时转入外源Cu/ZnSOD和APX抗氧化酶基因后,甘薯在水分胁迫下的SOD、APX、CAT等抗氧化酶活性均显著增强,膜系统受到的损伤也较轻,脯氨酸等渗透调节物质的积累升高[3- 4, 6]。在室内的水培试验中,转基因甘薯在水分胁迫条件下光合速率均高于对照植株,在随后的复水过程中恢复也较快[5]。在本试验中,在大田盆栽试验条件下也表现出类似的变化趋势,即在不同土壤水分条件下转基因甘薯叶片的光合速率均高于未转基因植株,尤其是在严重的干旱胁迫下两者的差异最为显著,表明同时转入的Cu/ZnSOD和APX抗氧化酶基因有利于甘薯在干旱逆境下维持较高的光合能力,结合之前的研究结果,可以推断出转入的抗氧化酶基因可能通过提高植株的抗氧化酶活性,及时有效的清除了逆境下生成过量的活性氧自由基,一定程度上保护了光合机构,使其维持较好的生理功能。
表1 不同土壤干旱程度下转基因(TS)和未转基因甘薯(NS)光合响应特征参数
Table 1 Light response characteristics of photosynthesis of transferred genes sweet potato (TS) and non-transferred control under different soil moisture
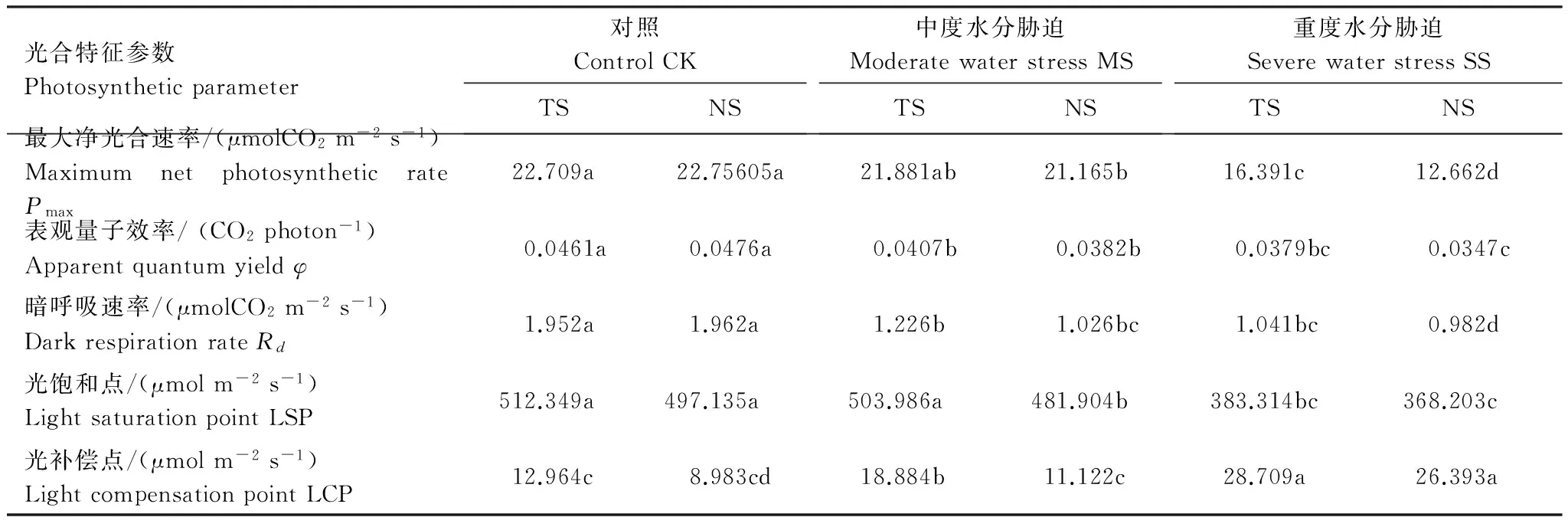
光合特征参数Photosyntheticparameter对照ControlCKTSNS中度水分胁迫ModeratewaterstressMSTSNS重度水分胁迫SeverewaterstressSSTSNS最大净光合速率/(μmolCO2m-2s-1)MaximumnetphotosyntheticratePmax22.709a22.75605a21.881ab21.165b16.391c12.662d表观量子效率/(CO2photon-1)Apparentquantumyieldφ0.0461a0.0476a0.0407b0.0382b0.0379bc0.0347c暗呼吸速率/(μmolCO2m-2s-1)DarkrespirationrateRd1.952a1.962a1.226b1.026bc1.041bc0.982d光饱和点/(μmolm-2s-1)LightsaturationpointLSP512.349a497.135a503.986a481.904b383.314bc368.203c光补偿点/(μmolm-2s-1)LightcompensationpointLCP12.964c8.983cd18.884b11.122c28.709a26.393a
一定的环境条件下,叶片的最大净光合速率(Pmax) 反映了植物叶片的最大光合能力[22- 23]。由表1可以看出,TS植株叶片的Pmax在严重干旱条件下也显著高于NS植株,但在正常的水分水平及中度干旱胁迫下二者无明显差别。由于干旱胁迫会影响有关酶的活性及光合电子传递,而Pmax决定于Rusbico 活性和电子传递速率[1,13, 24]。因此该研究结果表明在严重的干旱胁迫下转入的抗氧化酶基因有可能使甘薯叶片维持较高的PSⅡ电子传递速率及叶肉细胞Rusbico 活性,从而使TS植株叶片的Pmax保持了较高的水平。光能转化效率的高低直接决定叶片光合作用的高低,低光能转化效率是光合作用的重要限制因子[25- 26]。表观量子效率(φ),是反映植物在弱光下吸收转换和利用光能能力的指标,是光合作用中光能转化效率的指标之一,该值高,说明叶片光能转化效率高[24]。在本试验中,与对照相比,NS植株在中度干旱胁迫下就开始显著下降,而TS植株只是轻微下降,到严重干旱时,两者都显著下降,但是TS仍略高于NS植株,表明转入抗氧化酶基因可以有效的抑制甘薯在一定程度干旱胁迫条件下对弱光光能转化效率的降低,从而使其维持较高的光合能力。植物在干旱胁迫下能够维持其Pmax和φ在较高水平,表明其能有效地利用弱光及强光进行光合作用,对光的生态适应性强,并在一定程度上反映了植物光合作用对干旱的适应性较强[25]。由此可以推测,在叶片中同时转入SOD和APX抗氧化酶基因后,甘薯对光的适应性增强,对干旱的适应性也有所增强,这有助于拓展甘薯的种植地理范围。
不同植物的光响应曲线不同,光补偿点和光饱和点有很大的差异,同一种植物的光补偿点和光饱和点也不是固定数值,它们会随外界条件的变化而变动[20, 26]。在本试验中,随着土壤干旱程度增加,甘薯叶片光补偿点、光呼吸增高,光饱和点下降,表明干旱胁迫使气孔关闭,光能利用率降低,同时利用CO2的能力降低,光合产物消耗增加,不利于同化产物的积累,这与其他的研究结果类似[27- 31]。在严重缺水条件下,可能由于植物体内活性氧自由基代谢失调而引发生物膜结构和功能的破坏,导致光合作用的非气孔抑制。在干旱胁迫条件下,转基因甘薯叶片光补偿点、光呼吸增高以及光饱和点下降的幅度均明显小于未转基因植株,这可能是因为转入的抗氧化酶基因提高干旱胁迫下叶片中酶促防御系统的活性,加强的渗透调节物质的积累,适度缓解了干旱逆境引起的膜脂过氧化伤害,从而维护了细胞膜结构和功能的稳定性,改善了转基因植株在干旱逆境条件下的光合效率。
[1] 郭江, 郭新宇, 王纪华, 张凤路. 不同株型玉米光响应曲线的特征参数研究. 西北植物学报, 2005, 25(8): 1612- 1617.
[2] Jiang G M, He W M. A quick new method for determining light response curves of photosynthesis under field light conditions. Chinese Bulletin of Botany, 1999, 16(6): 712- 718.
[3] 李筠, 邓西平, 郭尚洙, 田中净. 转铜/锌超氧化物歧化酶和抗坏血酸过氧化物酶基因甘薯的耐旱性. 植物生理与分子生物学学报, 2006, 32(4): 451- 457.
[4] 李建梅, 邓西平. 干旱及复水条件下转基因甘薯抗氧化防御系统特性. 中国农学通报, 2007, 23(6): 616- 621.
[5] 李建梅, 邓西平. 干旱和复水条件下转基因甘薯的光合特性. 水土保持学报, 2007, 4(4): 193- 196.
[6] 陆燕元, 邓西平. 转入Cu/ZnSOD和APX基因对甘薯旱后复水的恢复作用. 西北农林科技大学学报: 自然科学版, 2010, 38(1): 67- 74.
[7] Kim K Y, Kwon S Y, Lee H S, Hur Y, Bang J W, Kwak S S. A novel oxidative stress-inducible peroxidase promoter from sweet potato: molecular cloning and characterization in transgenic tobacco plants and cultured cells. Plant Molecular Biology, 2003, 51(6): 831- 838.
[8] Li Y H, Zhou H J, Yang M S, Wang G Y, Lan H. Studies on drought-resistance of popular Langfang. Journal Northwest Forestry University, 2004, 19(1): 27- 31.
[9] Prioul J L, Chartier P. Partitioning of transfer and carboxylation components of intracellular resistance to photosynthetic CO2 fixation: a critical analysis of the methods used. Annals of Botany, 1977, 41(4): 789- 795.
[10] Pons T L, Anten N P R. Is plasticity in partitioning of photosynthetic resources between and within leaves important for whole-plant carbon gain in canopies? Ecology, 2004, 18: 802- 811.
[11] 叶子飘. 光响应模型在超级杂交稻组合-Ⅱ优明86中的应用. 生态学杂志, 2007, 26(8): 1323- 1326
[12] Lu P L, Luo Y, Liu J D, Yu Q. Characteristic parameters of light response curves of photosynthesis of winter wheat in north China. Quarterly Journal of Applied Meteorology, 2000, 3(2): 254- 260.
[13] 郎莹, 张光灿, 张征坤, 刘顺生, 刘德虎, 胡小兰. 不同土壤水分下山杏光合作用光响应过程及其模拟. 生态学报, 2011, 31(16): 4499- 4508.
[14] 吴芹, 张光灿, 裴斌, 徐志强, 赵瑜, 方立东. 不同土壤水分下山杏光合作用CO2响应过程及其模拟. 生态应用学报, 2013, 24(6): 1517- 1524.
[15] 王荣荣, 夏江宝, 杨吉华, 赵艳云, 刘京涛, 孙景宽. 贝壳砂生境干旱胁迫下杠柳叶片光合光响应模型比较. 植物生态学报, 2013, 37(2): 111- 121.
[16] 刘宇锋, 萧浪涛, 童建华, 李晓波. 非直线双曲线模型在光合光响应曲线数据分析中的应用. 中国农学通报, 2005, 21(8): 76- 79.
[17] Watling J R, Press M C, Quick W P. Elevated CO2induces biochemical and ultrastructural changes in leaves of the C4cereal sorghum. Plant Physiology, 2000, 123(3): 1143- 1149.
[18] 许大全. 光合作用效率. 上海: 上海科学技术出版社, 2002: 356- 358.
[19] 韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性. 生态学报, 2010, 30(15): 4019- 4026.
[20] 卜令铎, 张仁和, 常宇, 薛吉全, 韩苗苗. 苗期玉米叶片光合特性对水分胁迫的响应. 生态学报, 2010, 30(2): 5- 11.
[21] 付士磊, 周永斌, 何兴元, 陈玮. 干旱胁迫对杨树光合生理指标的影响. 应用生态学报, 2006, 17(11): 4- 9.
[22] 张劲松, 孟平, 高峻. 板蓝根光合及水分生理生态特性. 东北林业大学学报, 2004, 32(3): 26- 28.
[23] 林叶春, 曾昭海, 任长忠, 李志坚, 郭来春, 杨学超. 局部根区灌溉对裸燕麦光合特征曲线及叶绿素荧光特性的影响. 作物学报, 2012, 38(6): 1062- 1070.
[24] 伍维模, 李志军, 罗青红, 韩路. 土壤水分胁迫对胡杨、灰叶胡杨光合作用光响应特性的影响. 林业科学, 2007, 43(5): 6- 10.
[25] 夏江宝, 张光灿, 孙景宽, 刘霞. 山杏叶片光合生理参数对土壤水分和光照强度的阈值效应. 植物生态学报, 2011, 35(3): 322- 329.
[26] 罗维成, 曾凡江, 刘波, 张利刚, 刘镇, 宋聪, 彭守兰. 绿洲-沙漠过渡带引种植物光合生理特征研究. 草业学报, 2013, 2(1): 36- 36.
[27] 吴芹, 张光灿, 裴斌, 方立东. 3个树种对不同程度土壤干旱的生理生化响应. 生态学报, 2013, 33(12): 3648- 3656.
[28] 陈凯利, 李建明, 贺会强, 胡晓辉, 姚勇哲, 孙三杰. 水分对番茄不同叶龄叶片光合作用的影响. 生态学报, 2013, 33(16): 4919- 4929.
[29] 张海娜, 苏培玺, 李善家, 周紫鹃, 解婷婷, 赵庆芳. 荒漠区植物光合器官解剖结构对水分利用效率的指示作用. 生态学报, 2013, 33(16): 4909- 4918.
[30] 刘慧民, 车艳双, 车代弟, 闫永庆, 吴凤芝. 干旱胁迫对华北绣线菊和金山绣线菊光合能力的影响. 应用生态学报, 2010, 21(8): 2004- 2009.
[31] 郭春芳, 孙云, 张木清. 土壤水分胁迫对茶树光合作用-光响应特性的影响. 中国生态农业学报, 2008, 16(6): 1413- 1418.
Light response characteristics of photosynthetic of transgenic sweet potato under drought stress
LÜ Yanyuan1, MA Huancheng1, LI Haomin2, DENG Xiping3,*
1KeyLaboratoryofStateForestryAdministrationforBiodiversityConservationinSouthwestChina,SouthwestForestryUniversity,Kunming650224,China2YunnanAcademyofBiodiversity,SouthwestForestryUniversit,Kunming650224,China3StateKeyLaboratoryofSoilErosionandDrylandFarmingonLoessPlateau,NorthwestA&FUniversity,XinongRoadNo.26,Yangling712100,China
Water deficit is one of the major limiting photosynthesis of plants in many regions. A comparative study on photosynthetic characteristics between the genetic modified sweet potato transferred with the Cu/Zn SOD and APX genes (TS) and non-GM sweet potato (NS) were carried out with pot experiments. Three controls, namely the favorable soil moisture (75%—80% of field water capacity, FWC), the moderate drought (60%—65% of FWC) and the severe drought (35%—40% of FWC) groups were used. Result showed that light response curves of the TS and NS have no significant difference under the three soil moisture conditions. However, the photosynthetic rates of the TS were all higher than that of the NS. This was especially significant for the severe drought condition, which proved that the transcription of the Cu/Zn SOD and APX genes can help in maintaining a higher photosynthetic capacity under drought stress. To be specific, difference of the net photosynthetic rate (Pmax) between TS and NS was not notable under favorable and moderate drought conditions. But thePmaxof TS was obviously superior under severe drought stress. The apparent quantum efficiency (φ) of NS declined significantly under moderate drought. Comparatively, theφof TS only decreased slightly. Although theφvalues were both significantly decreased under severe drought stress, TS could still maintain a higher level ofφthan NS. This indicated that the transcription of antioxidant enzyme genes in sweet potato has promoted its overall photosynthetic capacity by inhibit the decrement of weak-light energy conversion efficiency under drought stress. The Light compensation point (LCP) would increase; and the light saturation point (LSP), apparent quantum efficiency (φ) and dark respiration rate (Rd) etc. would decrease when the degree of drought rose. Here, drought stress has compelled a comprehensive stomatal closure, reduced the utilization efficiency of light and CO2, raised the consumption of photosynthates. This is not conducive to the accumulation of anabolites. As the drought intensified, the increment of LCP, photorespiration and the decrement of LSP of TS were all smaller than that of NS. It′s possible that the transferred antioxidant enzyme genes have promoted the leaves′ bioactivity of the enzyme-recovery-system under drought stress. And strengthen of the osmolytes accumulation has alleviated the membrane lipid peroxidation injuries by drought stress.
transgenic gene; sweet potato; drought; photosynthetic response curves
国家自然科学基金地区科学基金(31260175)
2013- 06- 04;
日期:2014- 05- 08
10.5846/stxb201306041308
*通讯作者Corresponding author.E-mail: dengxp@ms.iswc.ac.cn
陆燕元,马焕成,李昊民,邓西平.土壤干旱对转基因甘薯光合曲线的响应.生态学报,2015,35(7):2155- 2160.
Lü Y Y, Ma H C, Li H M, Deng X P.Light response characteristics of photosynthetic of transgenic sweet potato under drought stress.Acta Ecologica Sinica,2015,35(7):2155- 2160.
猜你喜欢
科学(2022年4期)2022-10-25
江苏农业科学(2020年12期)2020-07-20
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
今日农业(2019年16期)2019-01-03
Coco薇(2016年5期)2016-06-03
西南农业学报(2016年5期)2016-05-17
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
新课程·中旬(2014年10期)2014-12-26
当代畜禽养殖业(2014年7期)2014-02-27
