
四明山脉溪流区水产经济动物棘胸蛙(Paa spinosa)成体形态性状对体质量和净体质量影响效应的性别差异*
2015-03-08 08:48:52斯烈钢李鸿鹏朱卫东申屠基康王志铮
海洋与湖沼 2015年3期
斯烈钢 富 裕 李鸿鹏 朱卫东 申屠基康 王志铮①
(1. 浙江海洋学院 舟山 316022; 2. 宁波市海洋与渔业研究院 宁波 315012; 3. 余姚市水产技术推广中心 余姚 315400)
棘胸蛙(Paa spinosa)俗称石蛙、石鸡等, 隶属于两栖纲、无尾目、蛙科、棘蛙属, 系分布于我国南方丘陵山区的特有大型食用蛙, 多生活于海拔 500—600m的溪流区中(林光华等, 1990), 具滴水产卵的繁育特性(丁松林等, 2009), 其肉嫩味美、营养丰富, 食用具滋补强身、清心润肺、健肝胃、补虚损, 以及解热毒、治疳疾等功效, 素有“百蛙之王”和“山珍”之美誉, 历来为我国居民喜食的名贵佳肴和高档滋补品。近年来, 受栖息地生境的持续恶化和山区农民酷采滥捕的影响, 我国棘胸蛙野生资源日趋衰退, 现已被中国物种红色名录列为易危(VU)等级。加强棘胸蛙野生种质资源保护, 积极发展以人工定向选育和优质亲体科学饲育为基础的棘胸蛙增养殖业, 无疑对促进山区农民创收致富和满足市场对棘胸蛙日益高涨的消费需求具重要现实意义。
体质量和净体质量既是决定水产养殖动物市售价格的重要变量, 也是实施水产养殖动物人工定向选育的重要目标性状, 但直观性明显弱于形态性状。研究表明, 水产养殖动物可借助通径分析找寻到用以标记体质量的关键形态性状组合(刘小林等, 2002),但两性个体在标记体质量的关键形态性状组成上却存在一定的差异(王志铮等, 2011)。因此, 弄清决定目标水产养殖动物两性成熟个体在体质量和净体质量形态标记上的差异, 并以此探析进而揭示两者在体质量和净体质量决定机制上的本质区别, 无疑对指导目标水产养殖动物两性成熟个体的人工定向选育和科学饲育具重要应用价值。鉴于棘胸蛙两性成熟个体具明显的异形特征(刘春涛等, 2007; 路庆芳等,2008), 且目前国内外涉及该蛙的研究主要集中于种质生物学(李炳华等, 1983; 胡知渊等, 2005)、繁殖生物学(路庆芳等, 2008; 俞宝根等, 2008)、生态生理学(张盛周等, 2003; 凌云等, 2012)、发育生物学(虞鹏程,1995)、人工养殖技术(谢海妹等, 2007)及营养成分分析(朱炳全, 2000)等方面, 而有关该蛙形态性状对体质量和净体质量的影响效应研究迄今尚未见报道之实际,本文作者于2013年6月16日以位于四明山脉溪流区的浙江省余姚市下鲁石蛙养殖场养成的3年龄棘胸蛙同生群养殖群体中的成熟个体为研究材料, 采用多元分析方法, 较系统地开展了棘胸蛙形态性状对体质量和净体质量影响效应的性别差异研究, 以期为提高该蛙优质亲体的选择精度和饲育效率提供科学依据。
1 材料与方法
1.1 供试蛙来源及数量
2013年6月16日于浙江省余姚市下鲁石蛙养殖场亲蛙培育池中, 随机选取产卵时间为 2010年6月20日的 3年龄余姚本地品系棘胸蛙同生群雌、雄成熟个体各84只作为本研究测定群体。
1.2 形态表型性状的测定
实验样品逐一编号后, 采用电子数显游标卡尺(精度 0.02mm)分别测量鼻间距(左右鼻孔间的水平距离, INS)、眼间距(左右上眼睑内侧缘间的水平距离,IS)、眼径(与体轴平行的眼之直径, ED)、吻长(吻端至眼前角的水平距离, SL)、头长(吻端至颈褶的水平距离, HL)、头宽(头部左右两侧的最大水平距离, HW)、肩宽(左右肩点间的水平距离, SW)、前肢长(肩关节至第三指末端的直线距离, FLL)、前肢臂长(肘关节至第三指末端的直线距离, FAL)、前足长(前肢基部至第三指末端的直线距离, FFL)、后肢长(体后端正中部位至第四趾末端的直线距离, HLL)、后足长(后肢基部至最长趾末端的的直线距离, HFL)、肩肛距(颈褶至肛孔后缘的的水平距离, SAL)、体长(吻端至泄殖腔孔后缘的水平距离, SUL)等 14项表型尺寸性状; 采用低温麻醉法测量实验蛙的体积(BV), 具体方法同日本鳗鲡(王志铮等, 2012); 采用CS1-JJ500Y型高精度电子天平(精度 0.01g)分别称量经滤纸吸净体表水分后的体质量(BW)和去除内脏后的净体质量(NW)。
1.3 数据处理
整理所测结果, 采用SPSS17.0软件分别计算实验蛙雌、雄测定群体各表型性状的均值、标准差以及变异系数, 以形态性状对BW、NW的相关分析和通径分析为基础, 剖析那些通径系数达到显著水平(P<0.05)的形态性状对BW、NW的直接作用和间接作用, 进而计算决定系数和复相关指数, 并以此确定影响BW、NW的关键形态性状组合; 通过偏回归分析, 保留那些偏回归系数达到显著水平(P<0.05)的形态性状, 建立这些性状对BW、NW的多元回归方程, 并进行拟合度检验。
2 结果与分析
2.1 参数估计值
由表1可见, 除IS、ED、SL等3项形态性状测量值无性别差异外, 其余12项形态性状及BW、NW测量值均呈雄性显著大于雌性(P<0.05); 除INS、ED、HL等3项形态性状测量值的变异系数呈雄性略小于雌性外, 其余12项形态性状及BW、NW测量值的变异系数均呈雄性大于雌性, 表明棘胸蛙在生长速度和体型可塑性上均存在较为明显的性别差异。
2.2 相关性分析
由表2可见, 棘胸蛙雌、雄测定群体所测形态性状与BW和NW间的相关性存在较为明显的异同, 即:(1) 雄性测定群体除FAL与BW和NW均仅呈显著相关(0.01<P<0.05)外, 其余形态性状均与BW和NW呈极显著相关(P<0.01); (2) 雌性测定群体 BW 除与ED、SL、FFL未呈显著相关(P>0.05), 与HL仅呈显著相关(0.01<P<0.05)外, 其余形态性状均与其呈极显著相关(P<0.01); (3) 雌性测定群体的NW除与ED、SL和HL未呈显著相关(P>0.05), 与INS、FFL仅呈显著相关(0.01<P<0.05)外, 其余形态性状均与其呈极显著相关(P<0.01); (4) 与雌、雄测定群体BW和NW相关系数达到0.85以上的形态性状均仅为BV。综上可知,棘胸蛙雄性成熟个体在形态性状对BW和NW的影响上较雌性更具一致性, 体型综合参数BV系影响棘胸蛙两性成熟个体BW和NW的关键公共变量。
2.3 通径分析
根据通径分析原理, 计算雌、雄测定群体各形态性状对BW和NW的通径系数。经显著性检验, 保留那些达到显著水平的形态性状(P<0.05), 并按相关系数组成效应, 将这些形态性状与BW和NW的相关系数(rij)剖分为直接作用(通径系数 P1)和通过其它性状的间接作用(P2), 并将结果列于表3。

表1 棘胸蛙两性个体生物学测定指标的统计量描述Tab.1 The statistics of various phenotypic and morphological traits for P. spinosa
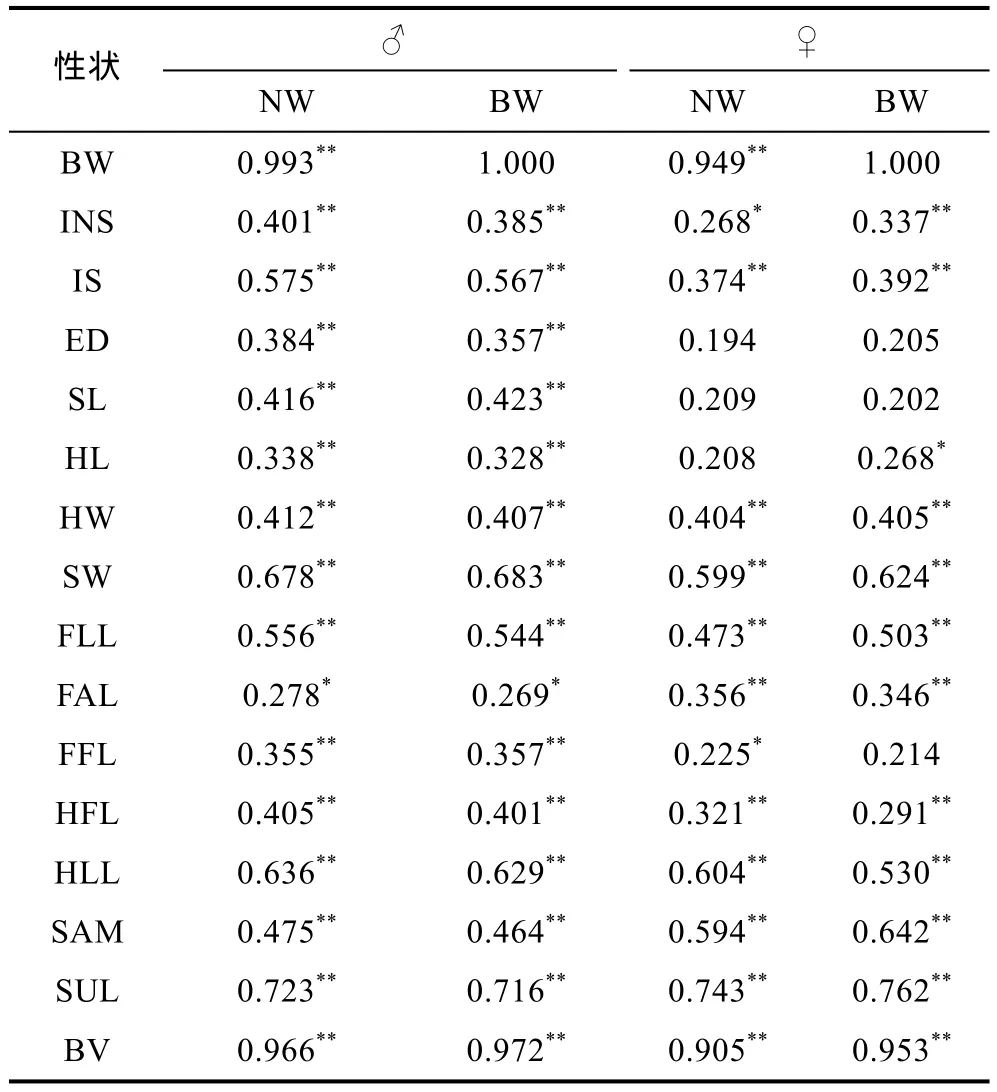
表2 棘胸蛙两性个体形态性状与体质量、净体质量间的相关系数(df=82)Tab.2 The correlation coefficients in phenotype among traits of P. spinosa
由表3可见, 雄性测定群体被保留的形态性状对BW和NW的直接作用依次呈BV(0.903) > SW(0.108)和BV(0.898) > SW(0.106), 间接作用依次呈SW(0.575) >BV(0.069)和 SW(0.572) > BV(0.068); 雌性测定群体被保留的形态性状对 BW和 NW的直接作用依次呈BV(0.887) > HLL(0.106) > HL(0.079)和 BV(0.796) >LHL(0.231), 间接作用依次呈HLL(0.425) > HL(0.189) >BV(0.066)和 HLL(0.373) > BV(0.108); 且上述 SW、LHL、HL的间接作用均主要通过BV来实现。综上,进一步表明 BV为影响棘胸蛙两性成熟个体 BW 和NW的决定性公共变量。
经计算, BV-SW组合对雄性测定群体BW和NW的相关指数R2分别为0.952和0.939; BV-HLL-HL组合和BV-HLL组合对雌性测定群体BW和NW的相关指数R2分别为0.922和0.860(表3, 表6), 均大于已找寻到影响目标变量的关键自变量组合的相关指数临界值0.85(刘小林等, 2002); 复共线性诊断也显示, 各形态性状的方差膨胀因子(VIF) 均远小于经验值(VIF=10),即各自变量间无显著相互干扰作用(表3)。表明BV-SW组合为影响雄性测定群体BW和NW的关键形态性状组合, BV-HLL-HL组合和BV-HLL组合分别为影响雌性测定群体BW和NW的关键形态性状组合。

表3 棘胸蛙形态性状对体质量、净体质量影响的通径分析Tab.3 The effects of morphologic traits on the weight of P. spinosa
2.4 决定程度分析
依表2、表3及表4所列数据, 分别计算棘胸蛙单个性状对BW和NW的决定系数(di=Pi2, Pi为性状对体重的通径系数)以及性状两两交互对BW和NW的共同决定系数(dij=2rijPiPj, rij为两性状间的相关系数, Pi、Pj分别为两性状对BW或NW的通径系数), 得表5。

表4 棘胸蛙被保留部分形态性状间的相关系数(df=82)Tab.4 Correlation coefficients of selected morphometric traits of P. spinosa
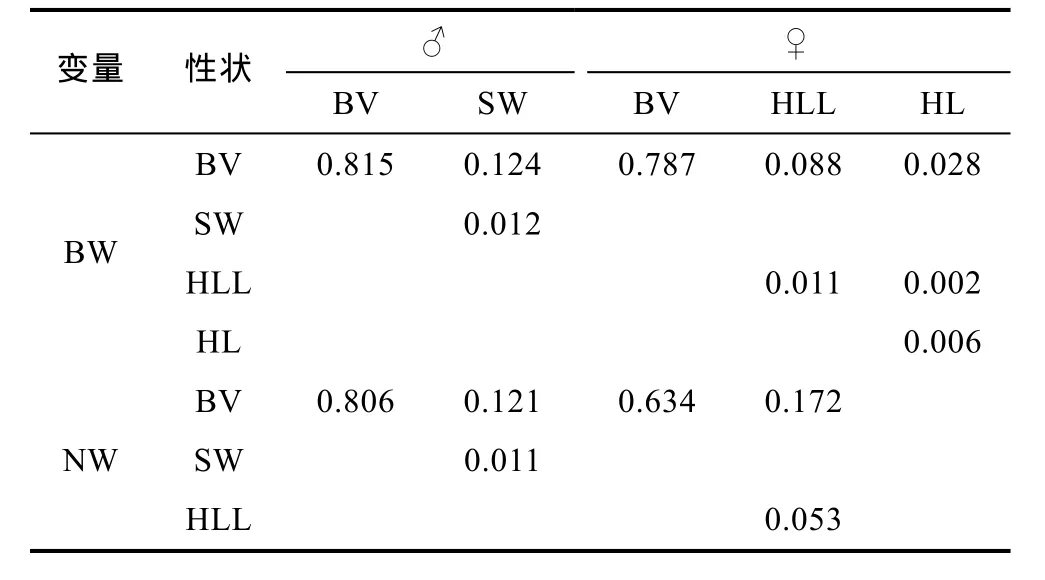
表5 棘胸蛙形态性状对体质量、净体质量的决定系数Tab.5 The determinant coefficients of morphometric traits on the weight of P. spinosa
由表5可见, BV、SW及两者交互对雄性测定群体BW和NW的决定系数加和值分别为0.952和0.942;BV、HLL、HL及其两两交互对雌性测定群体BW的决定系数加和值为0.923, BV、HLL及两者交互对雌性测定群体NW的决定系数加和值为 0.860, 均与其对应的相关指数R2值基本相等(表6), 且 BV对雄性测定群体BW 和NW 的决定程度分别达到 81.5%和80.6%, 对雌性测定群体BW和NW的决定程度分别达到 78.7%和 63.4%。上述结果, 进一步明确了BV-SW组合为决定雄性测定群体BW、NW的关键形态性状组合, BV-HLL-HL和BV-HLL依次为决定雌性测定群体BW、NW的关键形态性状组合, 从而再次确立了BV在决定棘胸蛙两性成熟个体BW和NW上的关键核心作用。
2.5 多元回归方程的建立
对经通径分析被保留的形态性状进行复相关分析和回归分析, 得表6、表7和表8。由表6可见, 棘胸蛙雌、雄测定群体中那些被保留的形态性状组合与其BW和NW间的复相关系数均达到极显著水平(R>r0.01[0]), 表明它们与BW和NW间存在极为密切的内在联系, 在对BW和NW的影响上较其它未被保留的形态性状更具重要性。
由表7可见, 用于估算雄性成熟个体BW和NW的回归方程分别为 BW(♂)= 0.894BV+0.750SW–16.481 和 WN(♂)= 0.727BV+0.600SW–10.184, 用于估算雌性成熟个体体质量和净体质量的回归方程分别为BW(♀)=0.841BV+0.220HLL+0.344HL–20.950和WN(♀)=0.543BV+0.346HLL–24.916, 且上述回归方程的回归截距及所涉形态性状的偏回归系数均达到显著水平(P<0.05), 另复共线性诊断也显示, 各形态性状的方差膨胀因子(VIF) 均远小于经验值。经方差分析和回归预测(表 8), 所建各回归方程的回归系数均达到极显著水平(P<0.01), 且估计值和实测值间无显著差异(P>0.05)。以上均表明, 所建方程能精确反映棘胸蛙形态性状与BW和NW间的相互关系。
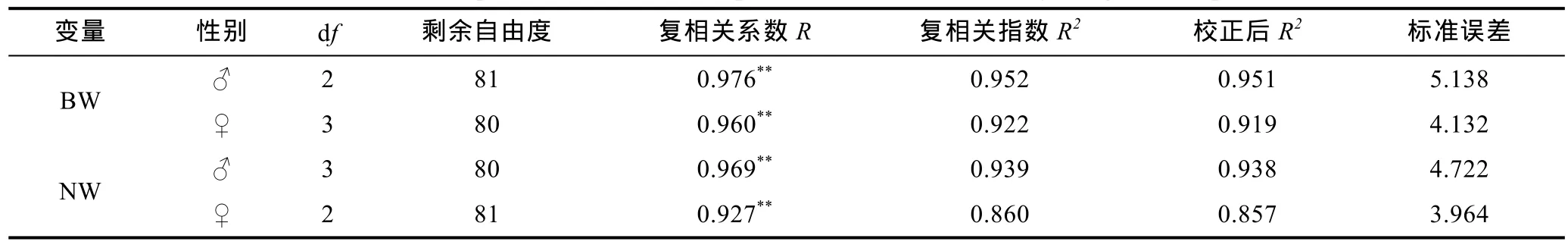
表6 棘胸蛙形态性状与体质量、净体质量间的复相关分析Tab.6 The multiple-coefficients of morphometric traits on the body weight of P. spinosa

表7 棘胸蛙形态性状与体质量、净体质量间的偏回归系数检验Tab.7 Test of partial regression and constants of P. spinosa
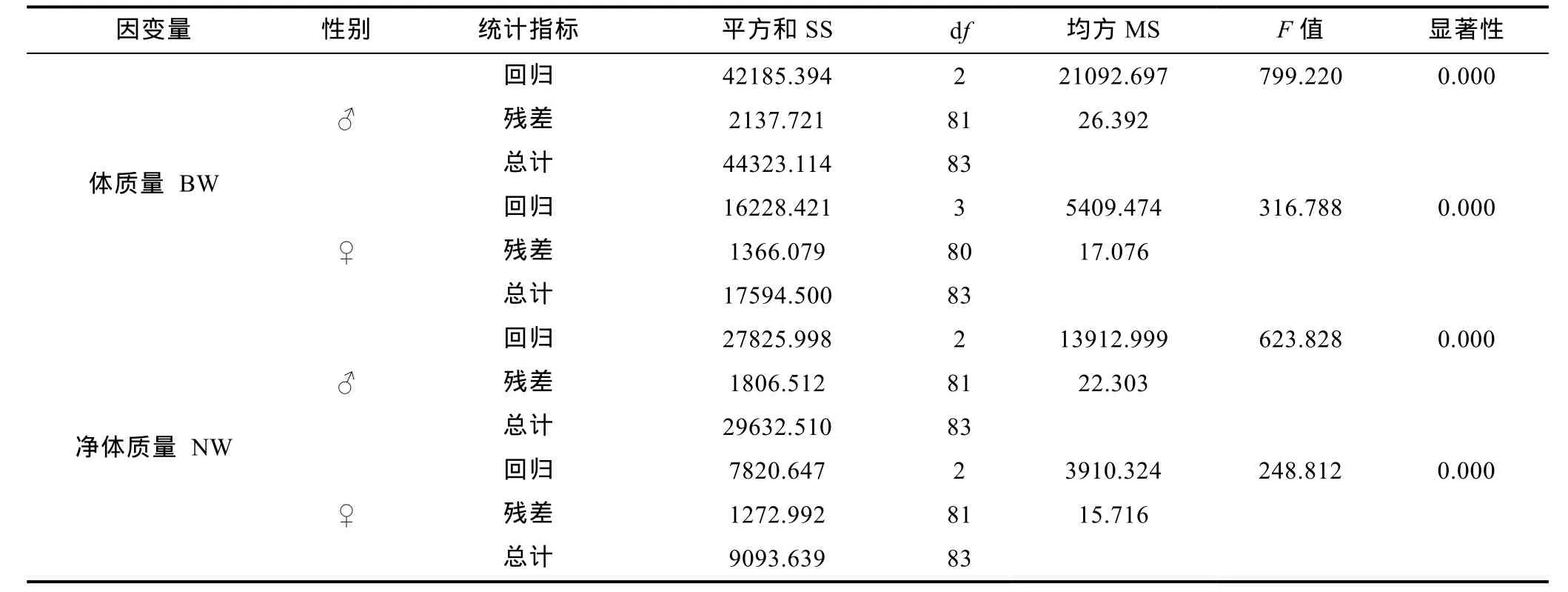
表8 棘胸蛙形态性状与体质量、净体质量间多元回归方程的方差分析Tab.8 Analysis of variance in multiple regression for P. spinosa
3 讨论
3.1 BV成为影响棘胸蛙同生群两性成体BW和NW核心公共变量的原因分析
BV作为表征同生群动物体型大小和体质强壮程度的综合性形态测量指标, 往往与BW、NW均有着极为密切的关系, 但表型生长、性选择“嗜好”及生育力选择压力的形态特征取向, 则是导致其能否成为影响BW、NW决定性变量的主因。本文作者认为, BV成为影响棘胸蛙同生群两性成熟个体BW、NW核心公共变量的原因, 主缘于两性成熟个体形态性状的等速生长以及体型对其繁殖适合度所具的重要影响力, 即: (1) 棘胸蛙两性个体的NW、HL、HW和FLL均随其 SUL的增长而呈等速增长(路庆芳等, 2008),以及与棘胸蛙同生群两性成熟个体BW和NW的相关系数达到0.90以上的形态性状均仅为BV, 且SUL与BW和NW的相关系数也均低于0.77(表2)的研究结果, 表明BV作为综合性形态测量指标在表征BW、NW上较其它形态测量指标更显全面和准确, 与相关报道认为影响水产动物体重的主要因素是使个体具有较大空间几何的性状的观点(耿绪云等, 2007; 王新安, 2008, 2013)相吻; (2) 棘胸蛙雌性的繁殖输出与个体大小呈正相关(路庆芳等, 2008), 生长速度较雄性个体慢(表 1), 到达初届性成熟年龄明显滞后于雄性(林光华等, 1990), 个体生殖力远低于林蛙(Rana temporovia)、牛蛙(Rana catesheiana)等其它常见经济蛙类(刘元楷等, 1990), 以及BV变异系数均大于其它测量指标, 而与BW和NW则均相近(表2)的特点, 既揭示了棘胸蛙雌性个体的性选择“嗜好”及生育力选择压力有驱使其体形朝大型化方向进化的倾向, 也反映了棘胸蛙与海蟾蜍(Bufo marinus) (Lee, 2001)、黑斑侧褶蛙(Pelopylax nigranaculata) (郑荣泉等, 2002)和多疣狭口蛙(Kaloula verrucosa) (贺佳飞等, 2006)等其它无尾类相似, 均具通过增大体型以增加腹腔怀卵容纳量来提高繁殖适合度的生殖对策, 表明BV既为表征棘胸蛙雌性成熟个体繁殖适合度的关键形态指标, 又是决定BW和NW的关键形态性状; (3) 无尾两栖类的外部形态进化保守(Marshall, 2005), 本研究中棘胸蛙同生群成体所表露的雄大雌小的体型特征(表1), 与浙江金华产(刘春涛等, 2007)和湖南平江产(路庆芳等, 2008)棘胸蛙两性异形特征的结果均完全相同, 表明地域分布的差异并不会改变该蛙属无尾两栖类两性异形中第二种类型(Shine, 1979)这一特质。在性选择中, 雄性应该是进攻的, 雌性则应该是挑剔的(孙儒泳等, 2002), 与体形较大的雄蛙交配获得的后代具更高的适合度(Sullivan et al, 1992), 体形较大的雄性棘胸蛙在争夺配偶中占有明显优势(路庆芳等, 2008)。Shine(1979)和 Davies等(1977)均指出,通过增大个体以在争斗中处于有利地位, 从而获得更多的交配机会, 是第二种两性异形类型中雄性常采取的策略。施林强等(2011)也认为, 该两性异形类型通常与促使雄性向较大体形方向进化的性选择压力较强有关。因此可将BV标定为指示雄蛙性成熟个体繁殖适合度的关键形态指标和决定BW和NW的关键形态性状。
3.2 SW和HLL在影响棘胸蛙同生群雌雄成体BW和NW上所起的作用
动物两性异形进化主要基于性选择(Shine, 1979;Anderson et al, 1990; Parker, 1992)。动物性选择的实现途径可能通过基于配偶竞争的性内选择或通过偏爱异性“奢侈”特征的性间选择, 或两者兼而有之(孙儒泳等, 2002)。因此, 本研究中除综合性形态测量性状BV外, 其它被保留的形态性状对BW和NW的直接影响作用均较小(波动于0.059—0.232之间)的情形(表 3), 无疑揭示了棘胸蛙同生群雌、雄成熟个体为谋求繁殖利益的最大化, 在性选择压力的作用下强化并保留了各自具一定“奢侈”特征的形态性状, 致使雄蛙的SW和雌蛙的LHL由此分别成为影响各自BW和NW的公共变量。
抱对是两栖类动物求偶、交配和完成繁殖过程极为重要的环节(尚玉昌, 1998)。棘胸蛙的抱对行为常表露为雄蛙追逐雌蛙, 并用前肢抱住雌蛙的后肢或躯体背部, 使两者泄殖腔相接近(俞宝根等, 2008);由此, 刘春涛等(2007)认为棘胸蛙雄蛙前肢显著长于雌蛙的原因是有利于其更好地抱对。本文作者认为上述直观认识并未能从本质上揭示对雄蛙抱对起关键作用的形态性状。本研究中, 棘胸蛙雄蛙SW与BW和NW的相关系数均分别大于对应的FLL, 以及FLL对雄蛙 BW和NW的通径系数均未达到被保留水平的结果(表2, 表3), 无疑真实反映了SW在对雄蛙增重和抱对成功率的贡献上较FLL更具重要性的事实,即雄蛙SW值越大, 则其捕食成功率和抱对成功率也就越高。
棘胸蛙具雌小、雄大的异形特征(刘春涛等, 2007;路庆芳等, 2008)和雌性背负雄性的抱对交配特性(林光华等, 1990; 俞宝根等, 2008)。路庆芳等(2008)研究发现, 棘胸蛙两性个体在局部形态上均无显著差异(P>0.05), 但雌性后肢长的增长速率则显著大于雄性(P<0.05)。上述情形, 无疑为本研究中HLL成为影响雌蛙BW和NW的公共变量的结果(表3)提供了极为重要的证据支持, 即较长而结实的后肢既有利于其弹跳取食和遇害逃逸, 也有助于其选择反应更为敏捷和体质更为强壮的配偶, 无疑对提高棘胸蛙雌性个体的存活率和繁殖效率具重要作用。至于, 本研究中HL未能被选入决定棘胸蛙雌性个体NW的形态性状组合(表 3)的原因, 则应跟其与 BW 呈显著相关(P<0.05), 而与NW未达到显著相关水平(P>0.05) (表2)有关。
丁松林, 郑宝成, 2009. 棘胸蛙繁育特性研究. 四川动物,28(4): 602—604
王志铮, 吴一挺, 杨 磊等, 2011. 日本沼虾(Macrobrachium nipponensis)形态性状对体重的影响效应. 海洋与湖沼,42(4): 612—618
王新安, 马爱军, 庄志猛等, 2013. 红鳍东方鲀(Takifugu rubripes)形态性状对体重的影响效果. 海洋与湖沼, 44(1):135—140
王新安, 马爱军, 许 可等, 2008. 大菱鲆幼鱼表型形态性状与体重之间的关系. 动物学报, 54(3): 540—545
朱炳全, 2000. 棘胸蛙营养成分的分析. 动物学杂志, 35(3):31—32
刘小林, 常亚青, 相建海等, 2002. 栉孔扇贝壳尺寸性状对活体重的影响效果分析. 海洋与湖沼, 33(6): 673—678
刘元楷, 苏向阳, 李荣招等, 1990. 棘胸蛙生殖腺发育的研究.福建水产, (2): 15—24
刘春涛, 郑荣泉, 路庆芳等, 2007. 棘胸蛙的两性异形和雌性繁殖特征. 见: 计翔. 两栖爬行动物学研究(第十一辑).南京: 东南大学出版社, 160—165
孙儒泳, 李庆芬, 牛翠娟等, 2002. 基础生态学. 北京: 高等教育出版社, 114
李炳华, 汪尊德, 1983. 棘胸蛙的染色体组型分析. 遗传, 5(5):39—41
张盛周, 黄徐根, 吴孝兵, 2003. 棘胸蛙消化道内分泌细胞的免疫组织化学定位. 动物学报, 49(6): 858—864
林光华, 虞鹏程, 1990. 棘胸蛙的繁殖生物学研究. 江西大学学报(自然科学版), 14(4): 64—70
尚玉昌, 1998. 动物行为学. 北京: 北京大学出版社, 58—100
郑荣泉, 杜卫国, 寿 鹿, 2002. 黑斑侧褶蛙的两性异形和雌性繁殖特征. 动物学杂志, 37(4): 14—18
胡知渊, 来雅萍, 陈文静, 2005. 棘胸蛙、虎纹蛙、黑斑蛙血细胞的比较. 四川动物, 24(1): 5—8
俞宝根, 叶容晖, 郑荣泉等, 2008. 人工环境下棘胸蛙(Paa spinosa)繁殖期的行为谱及活动节律. 生态学报, 28(12):6371—6378
施林强, 张秀琴, 马小梅, 2011. 泽陆蛙(Fejervarya limnocharis)两性异形的个体发育和雌体繁殖. 生态学杂志, 30(4): 717—723
贺佳飞, 周 伟, 李明会等, 2006. 多疣狭口蛙的两性异形.中南林学院学报, 26(1): 77—81
耿绪云, 王雪慧, 孙金孙等, 2007. 中华绒螯蟹(Eriocheir sinensis)一龄幼蟹外部形态性状对体重的影响效果分析.海洋与湖沼, 38(1): 49—54
凌 云, 邵 晨, 颉志刚等, 2012. 低温暴露和恢复对棘胸蛙雌性亚成体生存力及能量物质消耗的影响. 生态学报,32(18): 5763—5769
谢海妹, 袁久尧, 2007. 棘胸蛙养殖关键技术研究. 浙江海洋学院学报(自然科学版), 26(4): 457—460
虞鹏程, 林光华, 1995. 棘胸蛙早期胚胎发育的初步观察. 水生生物学报, 19(3): 216—222
路庆芳, 郑荣泉, 刘春涛等, 2008. 湖南平江棘胸蛙两性异形和雌性个体生育力. 浙江师范大学学报(自然科学版), 31(2): 220—224
Anderson R A, Vitt L J, 1990. Sexual selection versus alternative causes of sexual dimorphism in teiid lizards. Oecologia,84(2): 145—157
Davies N B, Halliday T R, 1977. Optimal mate selection in the toad Bufo bufo. Nature, 269(5623): 56—58
Lee J C, 2001. Evolution of a secondary sexual dimorphism in the toad, Bufo marinus. Copeia, 2001(4): 928—935
Marshall E, 2005. Will DNA bar codes breathe life into classification?. Science, 307(5712): 1037
Parker G A, 1992. The evolution of sexual size dimorphism in fish. Journal of Fish Biology, 41(supplement), http://dx.doi.org/10.1111/j.1095-8649.1992.tb03864.x
Shine R, 1979. Sexual selection and sexual dimorphism in the Amphibia. Copeia, 1979(2): 297—306
Sullivan B K, Hinshaw S H, 1992. Female choice and selection on male calling behaviour in the grey treefrog Hyla versicolor. Animal Behaviour, 44(4): 733—744
猜你喜欢
黑龙江气象(2021年2期)2021-11-05 07:06:58
世界科学技术-中医药现代化(2021年5期)2021-11-05 06:56:18
机械工程与自动化(2021年4期)2021-07-30 09:37:02
机械工程与自动化(2020年6期)2020-12-28 11:58:26
现代园艺(2018年2期)2018-03-15 08:00:10
数学理论与应用(2017年2期)2017-06-27 07:38:54
湖南中医药大学学报(2015年1期)2016-01-06 01:06:39
商洛学院学报(2015年3期)2015-12-21 03:36:22
化工进展(2015年3期)2015-11-11 09:06:57
材料研究与应用(2015年4期)2015-08-23 11:39:36
