
青山水库轮虫群落结构特征的时空差异对水质变化的影响*
2015-03-08 08:48:55刘成士邵晓阳
海洋与湖沼 2015年3期
刘成士 黄 燕 刘 瑜 刘 忱 邵晓阳①
(1. 杭州师范大学生命与环境科学学院 杭州 310036; 2. 浙江省环境保护科学设计研究院 杭州 310007)
轮虫是淡水浮游生物的重要组成部分之一, 是许多鱼类的主要饵料, 以真核类细菌和原生动物为食, 在水生态系统结构、功能及生物生产力中具有重要意义(Herzig, 1987; Lim et al, 1997)。通常轮虫占浮游动物生产量的10%—40%(Herzig, 1987), 轮虫不仅在淡水生态系统的结构功能、能量传递以及物质转换上具有重要意义(刘建康, 1999), 同时也是环境的重要指示生物(Sládeček, 1983)。公开的研究成果表明,轮虫的群落结构与水体水质有密切关系(谢平等,1996; 温新利等, 2004), 轮虫对水环境变化有较强的敏感性, 轮虫群落结构的生态特征可作为水质评价的依据。国内外有关轮虫生态学的研究已积累了较多的资料(Aare, 1983; Sládeček, 1983; 宗志祥等, 1993;Duggan et al, 2001; 李共国等, 2003; 周淑蝉等, 2006),但针对青山水库轮虫群落结构的研究公开报道的比较少。
杭州市青山水库位于杭嘉湖平原与浙西山区交界处、东苕溪主干南苕溪中下游, 距杭州市40 km, 距临安市8 km。水库集雨面积603 km2, 总库容2.13亿m3,是以防洪为主, 结合灌溉、发电等综合利用的大Ⅱ型水库, 是杭州市重要的饮用水备用水源地。近些年,库区人类活动加剧和区域经济社会发展, 生活、工业等点源污染, 农业、水土流失等面源污染, 以及库区周围的房地产开发等, 对水库的水质影响较大。因此,对水库的水质变动与水生生物群落演替之间的关系进行长期跟踪研究和分析就显得尤为必要。本文调查了 2012—2013年青山水库春、夏、秋、冬四个季节轮虫的密度、生物量、优势种以及生物多样性的动态变化, 并探究了青山水库轮虫群落结构特征与水质的关系, 从水质理化指标、综合营养状态指数、轮虫物种生物多样性指数等多个角度评价了青山水库水质的营养状况, 旨在为青山水库水质富营养化评价和水质保障提供基础资料。
1 材料与方法
1.1 采样点设置
根据青山水库水域形状(岸线、库湾)、主要支流入口、大坝位置、周边情况(交通、居民、农田等)、以及水深等情况设置9个采样点。样点分布如图1。

图1 样点分布图Fig.1 Distribution of sampling sites in Qingshan reservoir QSH代表采样点
1.2 实验方法
野外采样每个样点根据透明度等距离分表层、中层、下层, 定量样品用5 L采水器在不同水层分别采集水样, 倒入25号(Φ=64 μm)浮游生物网过滤, 收集滤过液注入60 mL标本瓶, 加3 mL福尔马林溶液摇匀后保存。野外采集、观察、监测方法依据《淡水浮游生物研究方法》(郭智明, 1984)和《湖泊生态调查观测与分析》(冯建社等, 1997)执行, 物种鉴定主要依据《淡水微型生物图谱》(郭智明, 1984)。
水温、浊度、pH值、溶解氧、电导率、氨氮、叶绿素含量现场用YSI 6600多功能水质检测仪直接测得; 透明度现场用透明度盘直接测量; 总磷、总氮及化学需氧量用采样瓶取水样100 mL带回实验室测得(总氮用岛津 TOC分析仪测得, 测量精度为0.05 mg/L;总磷和化学需氧量用国标法测得)。
1.3 数据处理与分析
根据中国环境监测总站制定的《湖泊(水库)富营养化评价方法及分级技术规定》, 运用综合营养状态指数法对青山水库水质状况进行评价。计算公式为:

公式中: TLI(Σ)—综合营养状态指数;
Wj—第j种参数的营养状态指数的相关权重;
TLI(j)—代表第j种参数的营养状态指数。
为避免使用单一多样性指数造成的评价分析偏差, 同时运用 Simpson多样性指数(D)、Shannon-Wiener多样性指数(H)、Margalef丰富度指数(Dmg)、Pielou均匀度指数(J)对轮虫群落特征进行分析。计算公式分别为(Spatharis et al, 2011):
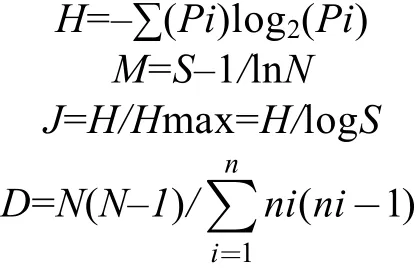
式中: S为生物的种类数; N为群落的个体总数; ni为第i种的个体数; Pi为第i种占总样品量的个体比例。
优势种是根据物种的出现频率及个体数量来确定, 用优势度来表示。优势度计算公式(Mchaughton,1967):
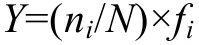
公式中: ni为第i种的个体数, N为所有种类总个体数,fi为第i种的出现频率。Y>0.02的种类定为优势种。
利用SPSS 19.0软件对数据进行统计分析以及利用Sigmaplot 10.0软件进行作图。
2 结果与分析
2.1 青山水库水环境参数变化
对照国家标准局颁布的《地表水环境质量标准》(GB3838-2002)分析青山水库水质检测指标的变化动态(图 2), 溶解氧年均值达到了 I—II类标准(5.10 mg/L),总磷属于I—II类(0.074 mg/L), COD(24.21 mg/L)、氨氮(14.27 mg/L)与总氮(3.79 mg/L)均超过 V 类标准,青山水库水质受氨氮、总氮、COD的影响较大。根据上述理化指标综合分析的结果, 青山水库水质处于 V类标准, 只能适用于农业用水及一般景观要求水域。
2.2 青山水库轮虫群落结构分析
2.2.1 种类组成、优势种及污染指示种 经分析鉴定, 共发现轮虫30种, 属于单巢目9个科10属。轮虫污染指示种共 17种, 其中 o-寡污性为 3种, 占17.6%; o-β 中污性为 6 种, 占 35.3%; β-中污性为 5 种,占 29.4%; β-α 中污性为 2 种, 占 11.8%; α-中污性为 1种, 占5.9%。从指示不同污染等级的轮虫种类占总数的比例看, 各级中污性种类种类占大多数。群落组成中, 常见种有针簇多肢轮虫(Polyarthra trigla)、长肢多肢轮虫(P. dolichoptera)、前节晶囊轮虫(Asplanchna priodonala)、卜氏晶囊轮虫(A. brightwelli)裂足臂尾轮虫(Brachionus diversicornis)和壶状臂尾轮虫(B.urceus)。春季优势种为长肢多肢轮虫和针簇多肢轮虫;夏季、秋季和冬季优势种都是针簇多肢轮虫。全年水平上, 青山水库优势物种是针簇多肢轮虫, 为β-中污类型指示种, 由此可见, 青山水库水质有一定程度的富营养化, 处于中度污染状态(表1)。

图2 理化因子的时间变化规律Fig.2 Time variation of physical and chemical factors
2.2.2 轮虫现存量分析 青山水库轮虫密度的季节变化: 夏季>春季>秋季>冬季。夏季密度最大, 各采样点的平均密度为 326 ind./L, 密度的最大峰值出现在8号采样点, 为1218 ind./L。该采样点共发现25种, 其中纵长异尾轮虫、卜氏晶囊轮虫和褶皱臂尾轮虫均以较高的密度出现, 分别为292、292、194 ind./L。密度最小峰值出现在5号采样点, 为36 ind./L, 该采样点仅发现针簇多枝轮虫单一种。其余采样点轮虫的密度均在200 ind./L左右波动。春季的平均密度为242 ind./L, 密度最大值出现在 4号采样点, 密度为450 ind./L, 最小值出现在2号, 密度为50 ind./L; 秋季和冬季的密度均较低, 平均密度只有 65 ind./L和52 ind./L, 这可能与秋冬两季水温降低、日照时间缩短、滤食性鱼类越冬前因营养储备而大量摄食、以及一些种类的轮虫繁殖期结束准备进入或处于休眠状态有关。空在间分布上, 8号采样点全年平均密度最大, 年平均密度为360 ind./L, 1号和2号采样点最低,分别是56 ind./L和58 ind./L(图3)。
轮虫生物量变化趋势基本与密度一致, 各季节平均生物量分别为: 0.0178、0.0294、0.01和0.0063 mg/L。
2.2.3 轮虫群落生物多样性指数 轮虫群落结构的季节变化, Simpon多样性指数和均匀度指数变化趋势基本吻合, 从春季至冬季依次递减, 且变化比较平缓, 春季分别为 0.91和 0.96, 冬季分别为 0.43和0.49; Margalef多样性指数和Shannon-Wiener指数变化趋势基本一致, 相较于前两个指数, 变化起伏较大,大小顺序为夏季>春季>秋季>冬季, 最高夏季为 1.88和1.70, 最低冬季仅为0.49和0.65, 最高值和最低值相差较大; 在空间水平上, 四种生物指数的最低值均出现在5号站点, 而最高值则均出现了两次峰值, 分别在3号点和8号点均出现了一次峰值(图4)。多样性指数的空间变化, D与J的变化趋势、M与H变化趋势是基本吻合的。
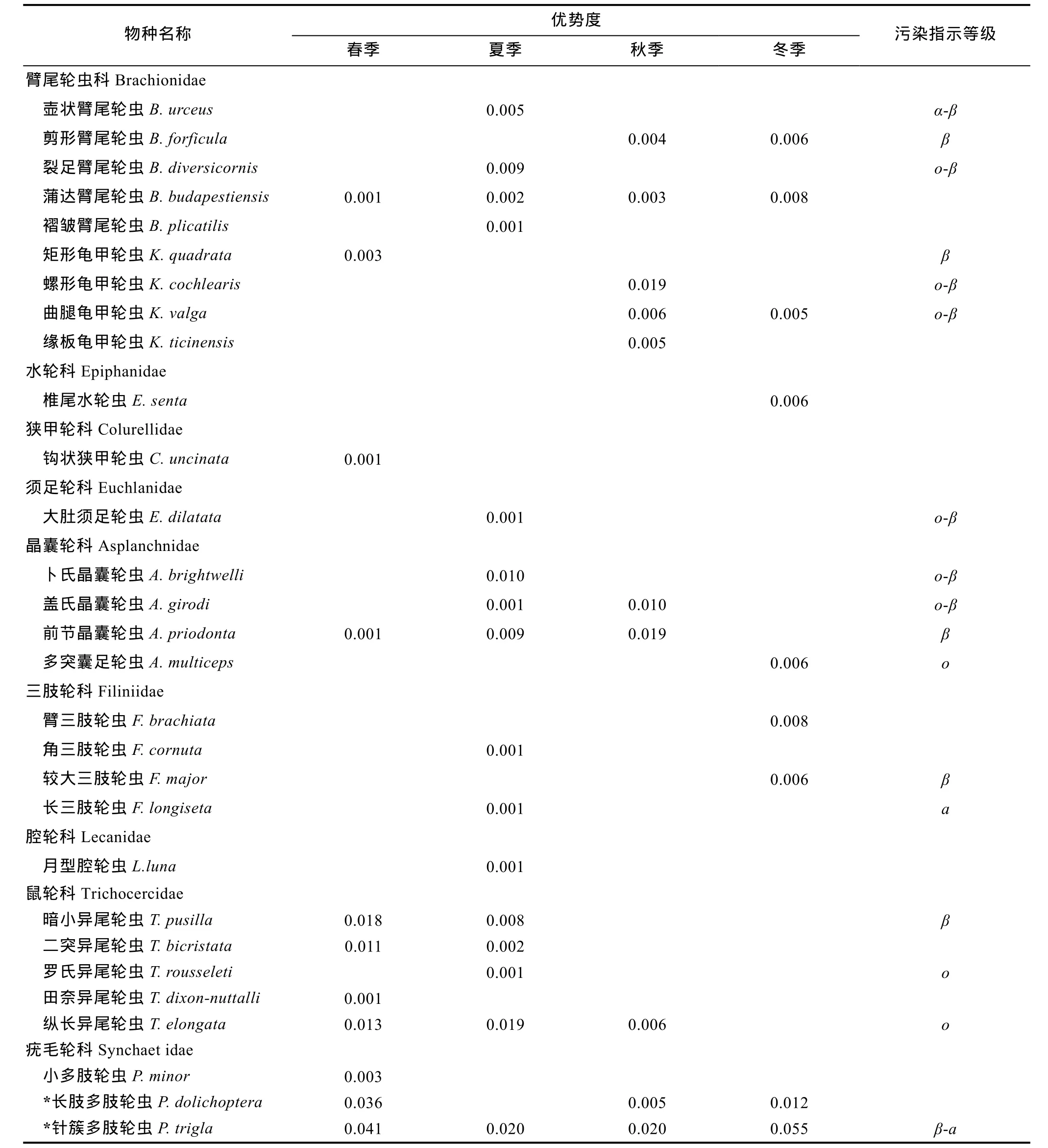
表1 青山水库轮虫优势度及污染指示种Tab.1 The pollution indicator species of rotifer in Qingshan reservoir

图4 轮虫生物多样性指数的时空变化Fig.4 The temporal and spatial variations in biotic indices of rotifers
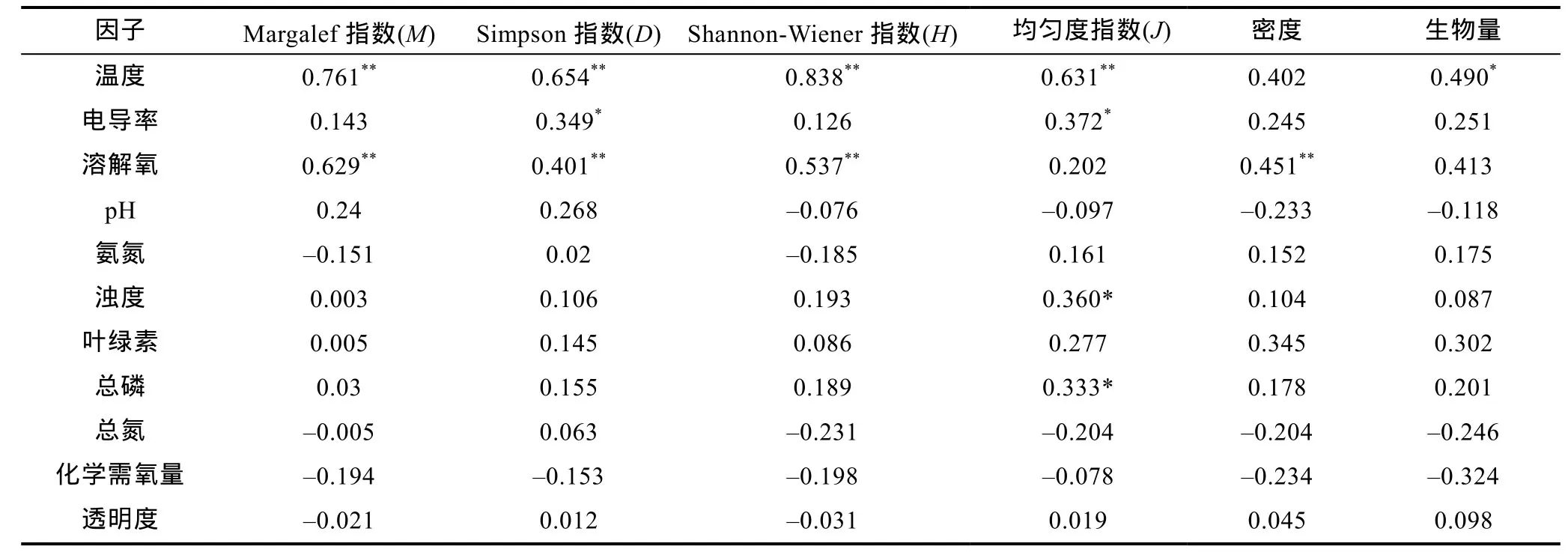
表2 青山水库轮虫群落结构与环境因子相关性分析Tab.2 Correlation of rotifers community structure and environmental factors in Qingshan reservoir
2.2.4 轮虫群落结构与环境因子相关性 依据各个季节中 9个站点的数据对青山水库各样点理化指标与轮虫密度、生物量以及多样性指数进行SPSS相关性分析(表2), 表中数据为Person相关性系数。
分析结果表明, 温度和溶解氧是影响青山水库轮虫群落结构演替的主要因素。轮虫密度与溶解氧呈极显著正相关(R=0.451, P<0.01), 生物量与温度呈显著正相关(R=0.490, P<0.05)。四种生物多样性指数都与温度呈极显著正相关, 并且除了均匀度外的其它三个指数都与溶解氧呈极显著正相关。通过SPSS多元回归分析, 轮虫群落结构不只受单一理化因子的影响, 多种理化因子的交叉、共同作用产生的影响较大。Margalef多样性指数受温度、氨氮和总氮的共同影响最大(R=0.837, P<0.01), Simpson多样性指数受温度、氨氮和pH的共同影响最大(R=0.701, P<0.01),Shannon-Wiener指数受温度和氨氮的共同影响最大(R=0.875, P<0.01), 均匀度指数受温度、氨氮和溶解氧的共同作用最大(R=0.864, P<0.01)。
2.3 聚类和主成分分析
依据各个站点轮虫群落的密度、生物量和生物多样性指数对青山水库库区进行系统聚类分析(图 5),得到结果: 青山水库库区采样点可分为四大类, 其中,上游1号和2号采样点为第一类, 4号、5号、6号、7号和9号采样点为第二类, 3号和8号采样点各自自成一类。这与库区的地形结构、人为活动影响的实际情况比较符合。1号和2号位于上游起始位置, 水位较浅且水体泥沙量较大, 枯水期库底暴露; 4号、5号和6号都是属于库湾区域, 均有山涧溪流汇入, 水质特征较为一致; 3号和8号分别为上游入库、下游出库与主库区的交汇水域, 水的动力作用对水域水质的影响与其它采样点有较大差异。
主成分分析统计分析(表 3), 因子 1、2、3和 4特征根值均大于1, 4个因子累计贡献率达到82.68 %,基本包含了全部指标具有的数据信息, 可以较准确地反映各站点的水质状况。按4个主成分的方式重新计算, 得到因子载荷矩阵, 则第一主成分因素为水温、溶解氧、pH, 第二主成分因素为电导率、叶绿素和总磷, 第三主成分因素为浊度和透明度, 第四主成分因素为氨氮和总氮、COD。青山水库主成分分析表明影响水体水质的主要因素是氨氮和总氮、COD。

图5 青山水库各采样点轮虫群落聚类树状图Fig.5 Cluster dendrogram of rotifers community at all sampling points in Qingshan reservoir
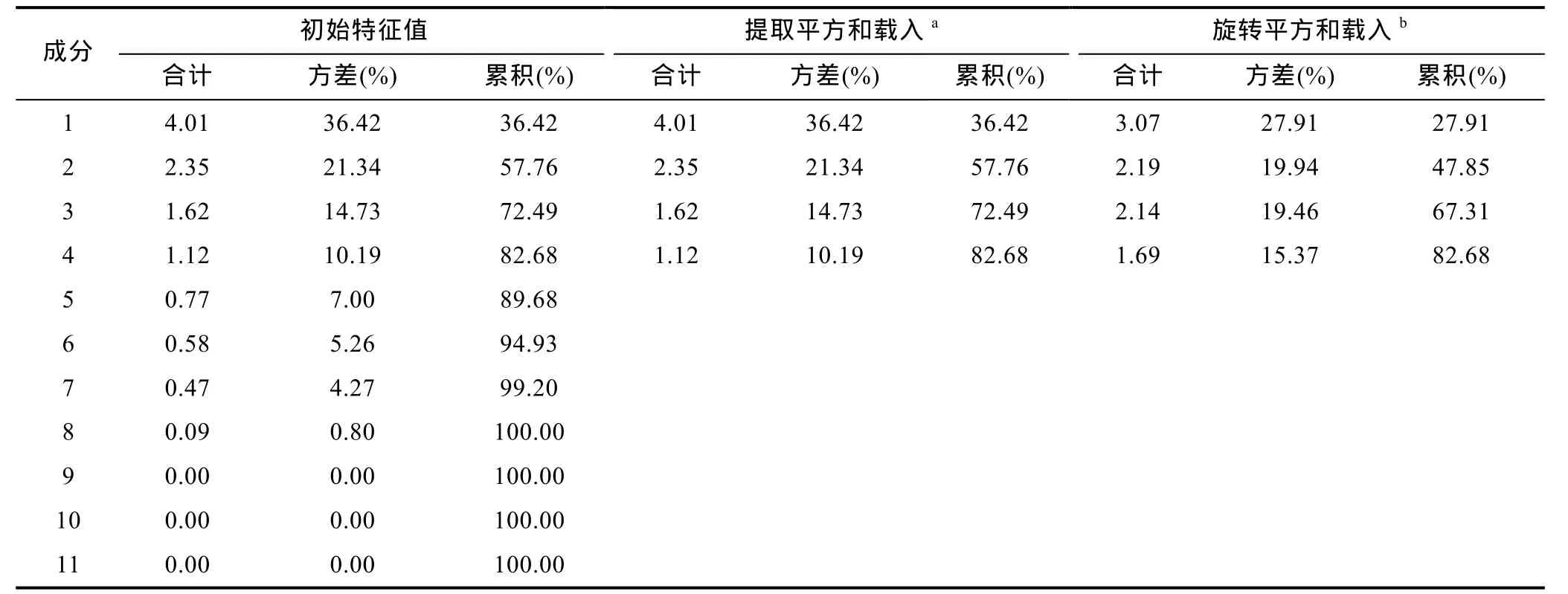
表3 主成分统计信息表Tab.3 Statistics of the principal component analysis
2.4 青山水库水质评价
2.4.1 富营养化指数综合评价 为避免单一因子评价的不足, 依据中国环境监测总站《湖库富营养化评价方法及分级技术规定》采用综合营养状态指数法对青山水库水体营养状态进行分析。在本次评价过程中, 综合考虑了叶绿素(Chal)、总磷(TP)、总氮(TN)、化学需氧量(COD)、透明度(SD)五项指标。 采用0—100的一系列连续数字对湖泊营养状态进行分级:在同一营养状态下, TLI(∑)指数值越高, 其营养程度越严重。评价结果表明, 青山水库所有采样点均属于中度富营养状态(表4)。

2.4.2 生物多样性指数评价 轮虫生物多样性指数常用于水质评价(熊金林等, 2003; Yoshida et al,2003), 本文利用轮虫群落的 Simpson多样性指数(D)、Shannon-Wiener多样性指数(H)、Margalef丰富度指数(Dmg)对青山水库水质进行评价, 评价结果表明, 三种生物多样性指数对水质评价结果基本一致,青山水库水质处于多污型-α中污型之间(表5)。
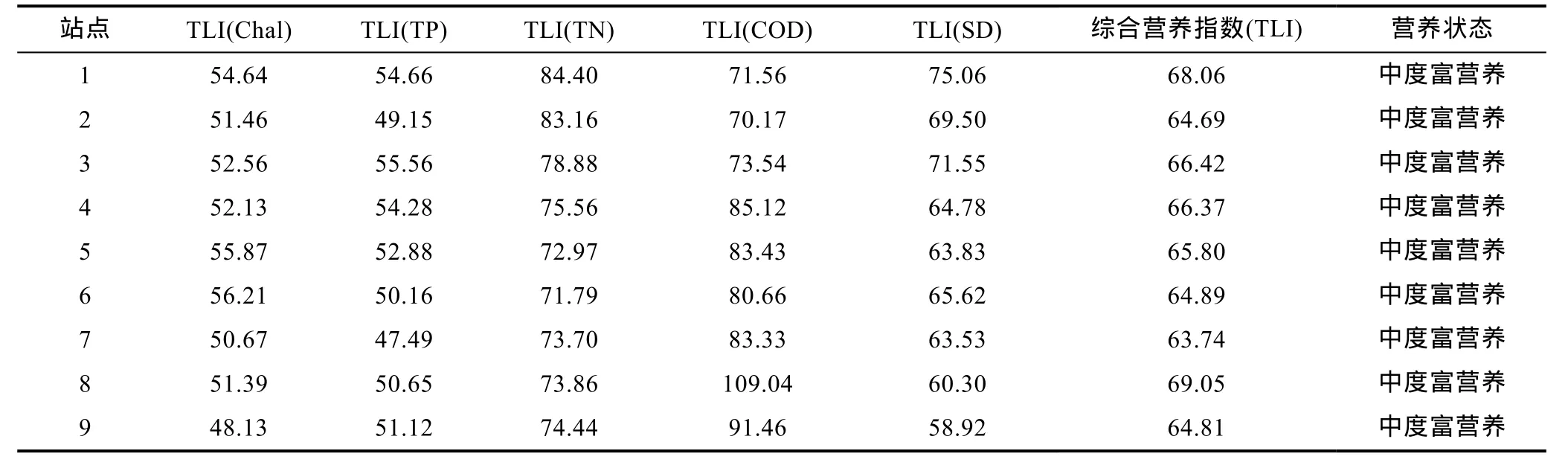
表4 青山水库各个站点的综合营养指数Tab.4 The overall nutrition index of each site in Qingshan reservoir

表5 轮虫群落生物多样性指数对水质的评价Tab.5 The assessment of water quality based on rotifers community biodiversity
根据评价过程中出现的一些问题, 对三种多样性指数的适用性提出一些观点: 首先, 多样性指数比较适用于一些环境较稳定的水体, 如相对封闭的湖泊、水库或者水流较缓、受外界影响小的河流等。郭智明提出对于流速大、流程长、底质变化多的河流利用多样性指数进行生物评价是不适宜的(郭智明,1984)。其次, 许多水体都不同程度受到外界的扰动,很难达到稳定的状态, 轮虫群落结构也在不断地变化。利用多样性指数较为客观地评价一个水体的水质状况, 需要尽可能地进行周期性地连续采样, 在完成一个调查周期后, 将指数值取平均, 这样的结果会比较客观。本文的研究就以年为周期分四个季度进行采样。最后, 由于每种指数的计算原理和方法均不相同,评价的侧重点也会有所不同。比较可行地是运用多种指数并且结合理化指标进行综合评价。冯建社等认为利用不同生物指数对水体进行评价有时会存在一定差异, 因此实际评价中不能仅根据一种指数结果就轻易下结论, 应综合考虑并结合理化监测结果, 才能得到符合实际的结论(冯建社等, 1997)。
3 讨论
3.1 影响轮虫群落结构的环境因子
水温、溶解氧、叶绿素、pH、营养盐等是影响轮虫生长发育、群落组成和数量变化等极为重要的环境因子, 是影响轮虫分布的重要因素。虽然诸多理化因子均与轮虫的发生及其季节变化有一定的相关性,但多数学者都认为决定轮虫种类季节演替的首要因子是水温(李共国等, 2003; Arora et al, 2003; 温新利等, 2004)。金琼贝等(1991)认为在 20—30°C, 随温度升高, 轮虫种类数会不断增加; 武汉东湖的长期研究显示, 轮虫总数的高峰一般出现在20°C以上的水温;温瑞塘河轮虫密度主要受到水温变化的影响, 两者呈显著正相关(肖佰财等, 2012)。随温度上升, 轮虫卵的发育时间缩短, 在丰富的食物供给条件下, 使种群周转加快, 导致种群密度迅速增加, 因此轮虫总数的高峰一般出现在20°C以上(章宗涉等, 1991)。夏、秋季水体中的藻类、动物尸体等在细菌的作用下, 很快形成碎屑(林婉莲等, 1984)。在25°C时, 壶状臂尾轮虫的产卵量显著高于其它各温度下的产卵量, 种群的净生殖率和内禀增长率最大(席贻龙等, 2000)。一些以碎屑为主要食物的轮虫因有丰富的食物而大量繁殖; 同时, 另一些以藻类、碎屑为主要食物的种类也得到相应的增长。本研究中显示, 青山水库轮虫密度和生物量高峰出现在夏、秋季, 与以往研究相一致。除水温外, 溶解氧也是一个影响轮虫群落演替的重要因素, Herzig(1987)提出, 在许多水体分层的富营养化湖泊中, 溶解氧的不足可以限制轮虫的发生。本研究中, 通过对环境因子与轮虫群落结构相关性分析得知, 轮虫密度、生物量以及生物多样性指数均与水温和溶解氧呈显著正相关, 水温和溶解氧是影响青山水库轮虫发生的主要因素, 这与以往研究相一致。
3.2 水质评价
轮虫发育时间快, 生命周期短, 能较为迅速地反映环境的变化, 被认为是很好的指示生物, 一般可根据湖泊中的轮虫种类演替来推测湖泊(水库)营养型的变化。国内外学者用轮虫评价水体污染状况的文章较多(宗志祥等, 1993; 张琦等, 1997; 林育真等, 1998;饶小珍等, 2000), Duggan等(2001)证实了不同营养状态的水体会出现不同的轮虫, 认为针簇多肢轮虫和独角聚花轮虫等在低营养水体更容易生长, 萼花臂尾轮虫和长三肢轮虫等更喜于在富营养水平水体中生存。还有研究认为小型浮游轮虫在贫营养水体中更有竞争力, 而大型浮游轮虫更喜于出现在富营养水体中(Gannon et al, 1978; Yoshida et al, 2003)。一般认为, 富营养湖泊典型指示种是: 臂尾轮属、裂痕龟纹轮虫、沟痕泡轮虫、圆筒异尾轮虫、长三肢轮虫、暗小异尾轮虫、螺形龟甲轮虫、矩形龟甲轮虫、真翅多肢轮(Gannon et al, 1978)。青山水库臂尾轮属5种, 其中春季1种, 冬季出现了4种, 秋季和冬季分别出现2种; 暗小异尾轮虫也是青山水库的常见种, 可能对低温比较敏感, 仅在春季和夏季出现, 在9个采样点的出现频率为50%。这些富营养水体的指示种在青山水库出现频率高,证明青山水库存在一定程度的富营养化趋势。
物种多样性是衡量群落稳定性的一个重要尺度,也是反映水体营养状况的重要参数(Zhan et al,1991)。通常多样性指数是反映丰富度和均匀度的综合指标, 而物种丰富度可以用Shannon-Wiener指数、Simpson指数和Margalef指数等多种指数来表示。以往学者的研究表明, 应使用2种或2种以上多样性指数评价水质, 以确保评价结果的可信性(Hill, 1973;孙军等, 2004)。故本研究利用Shannon-Wiener指数、Simpson指数和Margalef指数同时对青山水库水质状况进行评价。况琪军等人在利用多样性指数评价水体营养状态时指出: 若H值为0—1则水体重污染, 1—3为中污染, 大于3为轻污染或无污染; Margalef丰富度指数Dmg值0—3为α—中污染, 3—4为β—中污染,4—5为轻度污染, 大于5为清洁水(况琪军等, 2005)。本研究中, 基于H值和Dmg值分析结果, 青山水库水质处于中度—重度污染之间, 与轮虫群落优势种评价结果基本一致, 这也证实了水体环境可以影响轮虫种群或群落的结构特征。根据轮虫群落结构聚类分析表明,青山水库分为四大区域。主成分分析结果表明水质的下降可能与营养盐的增加有关, 特别是氮的输入。
4 结论
综合理化指标、综合营养状态指数、轮虫多样性指数以及轮虫优势种、指示种等多种评价方法, 可以认为在 2012—2013年调查期间, 青山水库水质属于中度—重度污染之间, 营养程度为中度富营养状态,青山水库水环境污染存在加重的趋势。因此, 迫切需要相关部门合作, 采取有效的措施来改善和保护青山水库周边区域的生物资源和水生态环境。综合全部的调查分析结果, 采用理化指标检测分析与轮虫群落结构特征评价水质状况, 得到的结果基本一致。因此, 本研究认为轮虫群落结构的时空动态变化是可以作为青山水库一个较为理想的水质监测指标。本文只对青山水库轮虫群落结构及水质评价作了初步的研究与探讨, 随着对青山水库水源地保育工作的不断开展、研究工作的不断深入, 针对这一方面的研究也会更加系统完善。
冯建社, 肖 勇, 钱志光, 1997. 用浮游动物及底栖动物的种群结构和数量分布评价河水水质的研究. 内蒙古环境保护, 9(2): 35—37
刘建康, 1999. 高级水生生物学. 北京: 科学出版社, 205
孙 军, 刘东艳, 2004. 多样性指数在海洋浮游植物研究中的应用. 海洋学报, 26(1): 62—75
李共国, 虞左明, 2003. 千岛湖轮虫群落结构及水质生态学评价. 湖泊科学, 15(2): 169—176
肖佰财, 孙陆宇, 冯德祥等, 2012. 温瑞塘河后生浮游动物群落结构及其与环境因子的关系. 水生态学杂志, 33(4):14—20
况琪军, 马沛明, 胡征宇等, 2005. 湖泊富营养化的藻类生物学评价与治理研究进展. 安全与环境学报, 5(2): 87—91
张 琦, 陈菊芳, 杞 桑等, 1997. 广东肇庆星湖浮游轮虫的调查及其与水质关系的研究. 生态科学, 16(2): 27—31
林育真, 李玉仙, 郭沛涌等, 1998. 东平湖轮虫群落与水质评价. 山东师范大学学报(自然科学版), 13(1): 63—67
林婉莲, 刘鑫洲, 刘健康, 1984. 四种浮游生物的碎屑形成过程. 水生生物学集刊, 8(2): 133—140
金相灿, 屠清瑛, 章宗涉, 1990. 湖泊富营养化调查规范(第二版). 北京: 中国环境科学出版社
金琼贝, 盛连喜, 张 然, 1991. 温度对浮游动物群落的影响. 东北师范大学学报(自然科学版), (4): 103—111
周淑蝉, 黄祥飞, 唐 涛等, 2006. 香溪河库湾轮虫现状及水质评价初探. 水生生物学报, 30(1): 52— 57
宗志祥, 许崇任, 任久长等, 1993. 洋河水库轮虫群落用于水质评价的研究. 环境科学学报, 13(1): 101—106
饶小珍, 许友勤, 陈寅山, 2000. 福州内河的轮虫与水质污染评价. 福建师范大学学报(自然科学版), 16(1): 71—75
郭智明, 1984. 滦河十四个断面大型底栖无脊椎动物的调查和水质评价. 环境科学, 5(4): 39—45
席贻龙, 黄祥飞, 2000. 温度对壶状臂尾轮虫实验种群动态的影响. 海洋与湖沼, 31(1): 23—28
章宗涉, 黄祥飞, 1991. 淡水浮游生物研究方法. 北京: 科学出版社, 38—47
温新利, 席贻龙, 张 雷等, 2004. 青弋江芜湖段轮虫群落结构和物种多样性的初步研究. 生物多样性, 12(4): 387—395
谢 平, 诸葛燕, 戴 莽等, 1996. 水体富营养化对浮游生物群落多样性的影响. 水生生物学报, 20(增刊): 30—37
熊金林, 梅兴国, 胡传林, 2003. 湖北四湖泊营养类型与轮虫群落的关系. 动物学杂志, 38(6): 8—14
Aare M, 1983. Rotifers as indicators of lake types in Estonia.Hydrobiology, 104(1): 357—361
Arora J, Mehra N K, 2003. Seasonal dynamics of rotifers in relation to physical and chemical conditions of the river Yamurna (Delhi), India. Hydrobiologia, 491(1—3): 101—109
Duggan I C, Green J D, Shiel R J, 2001. Distribution of rotifers in North Island, New Zealand and their potential use as bioindicators of lake trophic state. Hydroblologia, 446—447(1): l55— l64
Gannon J E, Stemberger R S, 1978. Zooplankton (especially Crustaceans and rotifers) as indicators of water quality.Transactions of the American Microscopical Society, 97(1):16—35
Herzig A, 1987. The analysis of planktonic rotifer populations: A plea for long-term investigations. Hydrobiologia, 147(1):l63—180
Hill M O, 1973. Diversity and evenness: A unifying notation and its consequences. Ecology, 54(2): 427—432
Lim L C, Wong C C, 1997. Use of the rotifer, Brachionus calyciflorus Pallas, in freshwater ornamental fish larviculture.Hydrobiologia, 358(1—3): 269—273
Mchaughton S J, 1967. Relationship among functional prosperities of California grassland. Nature, 216: 168—169
Sládeček V, 1983. Rotifers as indicators of water quality.Hydrobiologia, 100(1): 169—201
Spatharis S, Roelke D L, Dimitrakopoulos P G et al, 2011.Analyzing the (mis) behavior of Shannon index in eutrophication studies using field and simulated phytoplankton assemblages. Ecological Indicators, 11(2): 697—703
Yoshida T, Urabe J, Elser J J, 2003. Assessment of ‘top-down’and ‘bottom-up’ forces as determinants of rotifer distribution among lakes in Ontario, Canada. Ecological Research, 18(6):639—650
Zhan Y T, Yang C S, Fan Z N, 1991. Study on interrelation between distribution of phytoplankton and water pollution in Fuqi River. China Environmental Sciences, 11(1): 29—33
猜你喜欢
时代邮刊·上半月(2022年7期)2022-07-22 00:31:25
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
云南画报(2021年11期)2022-01-18 03:15:32
食品安全导刊(2021年20期)2021-08-30 06:40:50
科学养鱼(2020年10期)2020-11-23 09:04:10
宝藏(2020年4期)2020-11-05 06:48:36
课外语文·中(2020年2期)2020-03-24 22:18:09
课外语文(2020年5期)2020-03-14 08:16:40
知识窗(2019年4期)2019-04-26 03:16:02
水生生物学报(2015年1期)2015-02-28 16:01:05
