
杉木纯林土壤性质与林下植被的通径分析1)
2015-03-08 05:45卢妮妮高志雄张鹏徐雪蕾王新杰
东北林业大学学报 2015年7期
卢妮妮 高志雄 张鹏 徐雪蕾 王新杰
(北京林业大学,北京,100083)
责任编辑:王广建。
杉木作为一种重要的用材树种,其材质优良和生长迅速的特点深受林农的青睐[1]。第七次全国森林资源清查结果显示,我国杉木人工林面积达8.54×106hm2,占人工林总面积的21.35%[2]。杉木人工林树种单一,生态系统结构简单,抵抗力较低,这将大大影响杉木林的生长,降低木材产量[3-5]。林下植被作为森林生态系统的组成部分之一,在维护地力及促进养分循环方面发挥着重要的作用[6]。土壤理化性质和林分结构是决定林木生长的重要因子[7]。近年来,已有不少关于杉木林土壤理化性质的研究,并取得了一定的成果。有学者通过对土壤有机碳[8]、有机质[9]的研究,提出营造混交林[10-11]和发育林下植被[12-13]来改良土壤的方法。目前对林下植被与土壤的物理性质进行了简单的相关分析[14],相关系数虽然可以在一定程度上反映变量间的关系,但因其自身的片面性[15-16],限制了对因子之间关系的深入理解。通径分析不仅能准确表述变量间的真实关系,而且能把性状间的相关性分解成直接作用和间接作用。
以福建三明市将乐国有林场的杉木人工纯林为研究对象,选取土壤大量元素和微量元素指标,分析了不同林龄阶段杉木人工纯林土壤养分特征,并对土壤养分和林下植被进行通径分析为杉木林地力恢复及高产稳产提供理论依据。
1 研究区概况
研究区位于福建省西北部的三明市将乐国有林场(E 117°05'~117°40',N 26°26'~27°04')。该地区属低山丘陵区,平均海拔258 m;属中亚热带季风气候,年平均气温19.8 ℃;年平均降水量1 684 mm,全年降水量的80%发生在3—9月;境内夏季时间长,冬季较温暖,无霜期273 d;土壤类型主要为红壤,花岗岩母质,平均土层厚度60 cm;森林资源丰富,森林覆盖率达84.5%。主要造林树种为杉木(Cunninghamia lanceolata (Lamb)Hook)、马尾松(Pinus massoniana L.)、毛竹(Phyllostachys pubescens);常见灌木包括刚竹(Phyllostachys viridis)、盐肤木(Rhus chinensis)、乌饭树(Vaccinium bracteatum)、悬钩子(Rubus palmatus)、苦竹(Pleioblastus amarus)、粗叶榕(Ficus hirta Vahl)、黄瑞木(Adinandra millettii)、楤木(Aralia chinensis)等;主要的草本植物有芒萁(Dicranopteris dichotoma (Thunb.)Bernh)、山葡萄(Vitis amurensis Rupr.)、乌毛蕨(Stenoloma chusanum Ching)、龙牙草(Agrimonia pilosa Ledeb)、福建莲座蕨(Angiopteris fokiensis Hieron.)、狗脊蕨(Woodwardia orientalis Sw.)、五节芒(Miscanthus floridulus (Lab.)Warb.Ex Schum et Laut)、鱼腥草(Houttuynia cordata Thunb)、堇菜(Viola rockiana W.Beck)等。
2 试验数据与方法
2.1 样地选择
充分考虑海拔、坡向、坡度等立地因子,参考吴蔚东等[17]选择样地的原则,在福建省三明市将乐国有林场,选取立地条件相似、不同林龄阶段的杉木纯林(初始栽植密度为2 500 株/hm2)作为研究对象。标准地基本情况见表1。
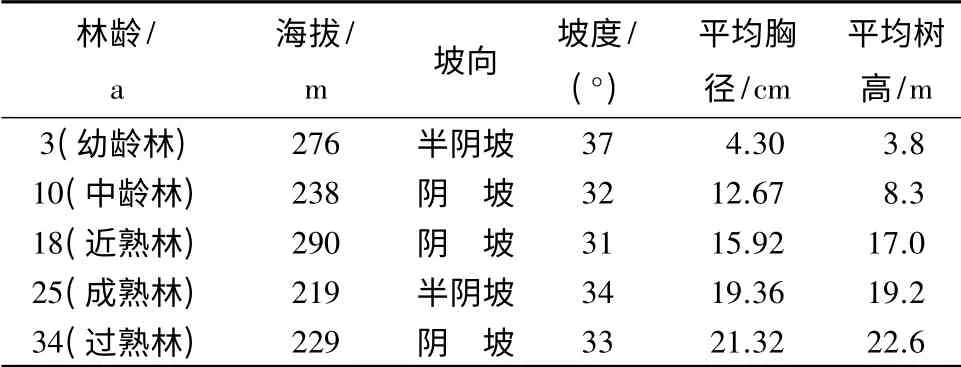
表1 标准地基本情况
2.2 研究方法
2.2.1 试验地设置
采用空间代替时间的方法,分别在3、10、18、25、34年生杉木林内设置20 m×20 m 的标准地,标准地设置3 个重复,对标准地林木进行每木检尺。灌木样方设置在标准地四角及中心,面积为5 m×5 m,调查灌木种类、高度、地径、盖度、生长状况和分布状况等。在灌木样方四角及中心设置1 m×1 m的草本样方,调查草本的种类、高度、盖度、株数、生长和分布状况等。标准地布设见图1。
2.2.2 土壤理化性质测定
在标准地合适位置挖掘土壤剖面,用环刀法取土样,取土深度为50 cm,每10 cm 为一层,分层测定土壤密度、含水率、毛管孔隙度和总孔隙度。同时在灌木样方中心用土钻取土,分5 层取样,每层取1 kg 样品装于自封袋,以测定土壤化学性质。具体测定方法参考杉木林生态系统定位研究方法[18]。

图1 标准地布设图
通过Excel 2010 和SPSS 18.0 对数据进行统计和分析,通径分析步骤参考杜家菊[19]的分析方法。
2.2.3 灌木和草本生物量测定
在标准地内详细记录每个样方中的灌木和草本的种类及生长状况,灌木分为叶(花、果)、枝干、根3部分分别实测鲜质量,草本分为地上部分和地下部分挖全株实测鲜质量,并分别对各部分取样。将各部分样品在鼓风干燥箱中于105 ℃恒温下烘干至恒质量,测量干物质质量,计算含水率,以此来计算生物量。每个林龄生物量样本不少于20 株。
3 结果与分析
3.1 不同林龄杉木纯林土壤物理性质
不同林龄的杉木人工纯林土壤物理性质测定结果如表2所示。由表2可知,不同林龄杉木纯林土壤物理性质随土壤深度变化具有相似规律。土壤密度随土层深度的加深而增大,且各土层土壤密度具有显著差异(P<0.05),>40~50 cm 土层的土壤密度比>0~10 cm 土层增大了16%。随着杉木林龄的增长,土壤密度逐渐增大,过熟林较幼龄林增大36%。土壤含水率随土层深度的加深而减少,幼龄林的土壤含水率最大,土壤含水率随林木林龄增长呈逐渐减小的趋势,但方差分析显示各林龄阶段土壤含水量整体无差异(P>0.05)。土壤孔隙度与土壤团聚直径和土壤中有机质质量分数有关,毛管孔隙借毛管力的作用保持水分,它们能影响土壤中的水、肥、气、热状况和林木生长[20]。图2显示,土壤总孔隙度整体上随着土层深度的加深而减小,>40~50 cm 土层土壤总孔隙度较>0~10 cm 土层减少了6.19%,且最上层和最下层土壤总孔隙度差异显著(P<0.05)。随着杉木林分的发育,土壤总孔隙度逐渐减小,过熟林时最小,但各林龄阶段总孔隙度无显著差异(P>0.05);毛管孔隙度在林木成熟过程中无明显变化。
3.2 不同林龄杉木纯林土壤化学性质
3.2.1 杉木纯林土壤中的大量元素
不同林龄杉木纯林土壤化学性质测定结果见表3。由表3可知,随着林分林龄的增大,不同林龄杉木纯林土壤大量元素(钾、氮、磷、有机质)质量分数呈现相似变化规律。全钾、全氮、全磷质量分数呈逐渐增加趋势,且各林龄阶段质量分数差异显著(P<0.05);速效钾、速效磷和有机质质量分数均表现为先减小再增大的趋势,在近熟林时达到最小值,方差分析显示近熟林速效钾、速效磷和有机质质量分数与幼龄林、过熟林具有显著差异(P<0.05),与中龄林和成熟林差异不显著(P>0.05)。垂直方向上,随着土层深度的加深,土壤中速效钾、碱解氮、速效磷、全氮和有机质质量分数逐渐减少,其中各土层速效钾、碱解氮和有机质质量分数差异显著(P<0.05),速效磷质量分数无显著差异(P>0.05)。各土层全钾质量分数和全磷质量分数无明显变化规律,且各土层质量分数无显著差异(P>0.05)。

表2 不同林龄杉木纯林土壤物理性质

表3 不同林龄杉木纯林土壤化学性质
3.2.2 杉木纯林土壤中的微量元素
从表3可知,各林龄阶段杉木纯林土壤微量元素质量分数变化具有相似特征。随着林分林龄的增大,铝和锌元素质量分数呈先减少后增加的趋势,极小值点出现在成熟林阶段。铁元素的变化则相反,表现为先增加后减少,极大值点出现在中龄林阶段。镁、钠、钙元素的质量分数随林分林龄的增加始终在增加,过熟林阶段的钠元素质量分数为幼龄林阶段的2 倍,反映了杉木纯林在发育过程中将导致土壤碱化,在经营时应注意建设林地的灌溉排水设施。各林龄阶段锰元素质量分数无明显变化。方差分析显示,各林龄阶段杉木纯林土壤铝、铁、锌和钠元素质量分数变化差异显著(P<0.05),镁、钙和锰元素的质量分数变化无显著差异(P>0.05)。
垂直方向上,铝、铁和镁元素质量分数随着土层深度的加深逐渐增加,其中各土层铝和铁元素质量分数差异显著(P<0.05),镁元素质量分数无显著差异(P>0.05)。钙和锰元素质量分数均随着土层深度加深先减少后增加,极小值点均出现在>30~40 cm 土层,且该层钙和锰元素质量分数与其他土层质量分数差异显著(P<0.05)。钠元素质量分数呈逐渐减小趋势,各土层质量分数差异显著(P<0.05),>40~50 cm 土层比>0~10 cm 土层减少了31.5%,表明土壤碱化主要发生在表层土壤。随着土层深度的加深,锌元素质量分数无显著变化(P>0.05)。
3.3 不同林龄杉木纯林林下灌草多样性及生物量
不同林龄杉木纯林下灌木和草本的Shannon-Wiener 多样性指数如表4。灌木的Shannon-Wiener指数随杉木林分林龄的增加呈先减小后增大的趋势,成熟林阶段灌木的多样性指数最小。方差分析显示,杉木发育的各个阶段灌木多样性差异不显著(P>0.05)。随着杉木的生长,林下草本多样性指数先后出现一个增加和减少的变化过程,在近熟林时达到最大值,比幼龄林时增加了34.67%,且幼龄林与其他成长阶段林下草本多样性指数差异显著(P<0.05)。
林下灌木和草本的生物量一定程度上反映了乔木层的树冠状况和土壤的肥沃程度[21],对不同林龄的杉木纯林中的灌木和草本的生物量进行测定,结果如表4。由表4可知,灌木生物量随着林分林龄的增加始终在增大,在过熟林中达到最大值,各个发育阶段除幼龄林和中龄林差异不显著外(P>0.05),均具有显著的差异(P<0.05)。随着林分的发育,草本的生物量先增大后减小,在成熟林中达到最大值。
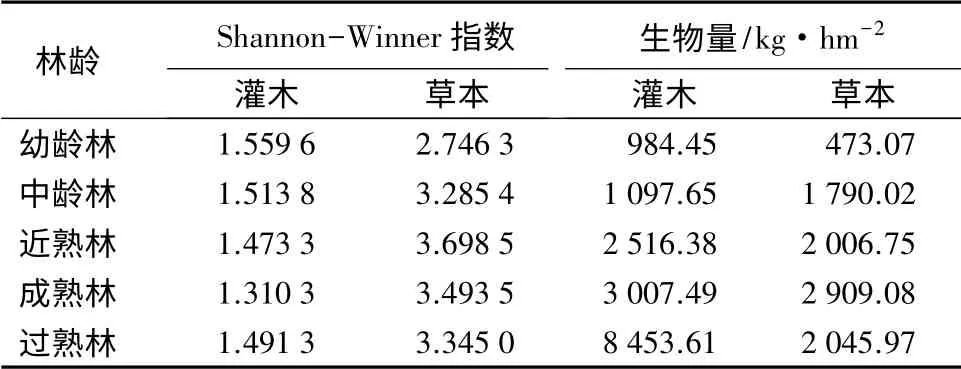
表4 不同林龄杉木纯林下灌草的多样性和生物量
3.4 土壤理化性质与林下植被多样性和生物量的通径分析
对不同发育阶段的杉木纯林的土壤理化指标与林下植被的多样性指数和生物量进行通径分析,结果如表5。由表5可知,全磷对灌木多样性有较大的直接通径系数,表观表现为全磷与灌木多样性显著相关(P<0.05)。对草本多样性有较大直接通径系数的是土壤化学元素中的速效钾、全钾、铁和锌,其中速效钾和锌与草本多样性显著负相关(P<0.05)。总孔隙度、土壤密度和锌对灌木生物量的直接通径系数较大,同时通过其他指标对灌木生物量的间接通径系数也较大,表观表现为总孔隙度和土壤密度与灌木生物量极显著相关(P<0.01),锌与灌木生物量显著相关(P<0.05),说明总孔隙度、土壤密度和锌是影响灌木生物量大小的主要因素。毛管孔隙度只对草本生物量有较大的直接通径系数,且两者之间呈极显著相关关系(P<0.01);速效钾对草本生物量有较大的直接通径系数。整体上,土壤物理性质对灌木生物量和草本生物量有较大的影响,土壤化学指标中速效钾、全钾、全磷、铁和锌对灌木多样性、草本多样性、灌木生物量和草本生物量均达到显著相关(P<0.05)。

表5 土壤理化性质对林下植被的通径系数
4 结论与讨论
不同林龄杉木纯林土壤物理性质随土壤深度变化具有相似规律。土壤密度随土层深度的加深而增大,且各土层土壤密度具有显著差异(P<0.05)。随着杉木林龄的增长,土壤密度逐渐增大,过熟林较幼龄林增大36%。土壤含水率和总孔隙度均随土层深度的加深而减少,幼龄林时期的土壤含水率和总孔隙度最大。随着杉木林分的发育,土壤含水率和土壤总孔隙度逐渐减小,过熟林时最小。毛管孔隙度在林木成熟过程中无明显变化。
随着林龄的增大,全钾、全氮、全磷质量分数呈逐渐增加趋势,且各林龄阶段质量分数差异显著(P<0.05);速效钾、速效磷和有机质质量分数均表现为先减小再增大的趋势,这与盛炜彤[22]的研究结果类似,表明在杉木发育过程中,林地的土壤肥力在逐渐下降后略有回升;垂直方向上,随着土层深度的加深,土壤中速效钾、碱解氮、速效磷、全氮和有机质质量分数逐渐减少。微量元素中的铝和锌元素质量分数随林分林龄的增加呈先减少后增加的趋势,铁元素则表现为先增加后减少,镁、钠、钙元素的质量分数始终在增加,锰元素质量分数无明显变化。垂直方向上,铝、铁和镁元素质量分数随着土层深度的加深逐渐增加,钙和锰元素质量分数则先减少后增加,钠元素质量分数呈逐渐减小趋势。
随着林龄的增加,灌木的Shannon-Wiener 指数先减小后增大,生物量逐渐增大;草本的Shannon-Wiener 指数和生物量均先增大后减小。
土壤物理性质对灌木生物量和草本生物量较大的影响,土壤化学指标中速效钾、全钾、全磷、铁和锌对灌木多样性、草本多样性、灌木生物量和草本生物量均达到显著相关(P<0.05)。本试验主要研究了不同发育阶段杉木纯林林地土壤理化性质及其对林下植被的影响,认为人工促进林下植被更新可对部分土壤养分的循环起到一定的作用,为杉木林地力恢复、可持续经营提供了理论依据。但土壤元素对林下植被的影响由气候环境、群落结构、土壤酶、真菌等因素共同决定,因此,结合土壤微生物群落环境研究土壤元素与林下植被的关系将成为下一步研究的重点。
[1] 余新妥.中国杉木研究[J].福建林学院学报,1988,8(3):203-220.
[2] 国家林业局.全国森林资源统计:第七次全国森林资源清查[M].北京:中国林业出版社,2010.
[3] 杨玉盛,何宗明,陈光水,等.杉木多代连栽后土壤肥力变化[J].土壤与环境,2001,10(1):33-38.
[4] 杨玉盛,陈光水,黄宝龙.杉木多世代连栽的土壤水分和养分变化[J].南京林业大学学报,2000,24(2):25-28.
[5] 侯银.林分结构对杨树人工林土壤微生物多样性的影响[D].南京:南京林业大学,2013.
[6] 李博,杨持,林鹏.生态学[M].北京:高等教育出版社,2000.
[7] 刘方,罗汝英,蒋建屏.土壤养分状况与杉木生长[J].南京林业大学学报,1991,15(2):41-46.
[8] 王丹,王兵,戴伟,等.不同发育阶段杉木林土壤有机碳变化特征及影响因素[J].林业科学研究,2009,22(5):667-671.
[9] Dixon R K,Brown S,Houghton RA,et al.Carbon pools and flux of global forest ecosystems[J].Science,1994,263:185-190.
[10] 邓仕坚,张家武,陈楚莹,等.不同树种混交林及其纯林对土壤理化性质影响的研究[J].应用生态学报,1994,5(2):126-132.
[11] 何贵平,陈益泰,胡炳堂,等.杉木与马褂木、檫树混交林及其纯林生物量和土壤肥力研究[J].林业科学研究,2001,14(5):540-547.
[12] 姚茂和,盛炜彤.林下植被对杉木林地力影响的研究[J].林业科学研究,1991,4(3):246-252.
[13] 盛炜彤,杨承栋.关于杉木林下植被对改良土壤性质效用的研究[J].生态学报,1997,17(4):377-385.
[14] 郭琦,王新杰,衣晓丹.不同林龄杉木纯林林下生物量与土壤理化性质的相关性[J].东北林业大学学报,2014,42(3):85-88,98.
[15] 刘广深,徐冬梅,许中坚,等.用通径分析法研究土壤水解酶活性与土壤性质的关系[J].土壤学报,2003,40(5):756-762.
[16] 明道绪.通径分析的原理与方法[J].农业科学导报,1986,1(4):39-43.
[17] 吴蔚东,张桃林,高超,等.红壤地区杉木人工林土壤肥力质量性状的演变[J].土壤学报,2001,38(3):285-293.
[18] 田大伦.杉木林生态系统定位研究方法[M].北京:科学出版社,2004.
[19] 杜家菊,陈志伟.使用SPSS 线性回归实现通径分析的方法[J].生物学通报,2010,45(2):4-6.
[20] 田洪艳,周道玮,李质馨,等.土壤胀缩运动对草原土壤的干扰作用[J].草地学报,2003,11(3):261-268.
[21] 陈民生,赵京岗,刘杰,等.人工林林下植被研究进展[J].山东农业大学学报:自然科学版,2008,39(2):321-325.
[22] 盛炜彤,杨承栋,范少辉.杉木人工林的土壤性质变化[J].林业科学研究,2003,16(4):377-385.
猜你喜欢
防护林科技(2022年5期)2022-08-22
黑龙江气象(2021年2期)2021-11-05
数理化解题研究(2021年13期)2021-08-19
机械工程与自动化(2020年6期)2020-12-28
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
森林工程(2018年4期)2018-08-04
方圆(2018年23期)2018-01-07
广西林业科学(2016年4期)2016-03-16
亚热带资源与环境学报(2015年1期)2015-01-22
