
狂犬病病毒固定株感染对微管结合蛋白EB3表达的影响
2015-03-08 09:35李莹莹陈云云任严新WaqasAhmad关振宏张茂林
中国预防兽医学报 2015年5期
宋 颖,李莹莹,陈云云,任严新,Waqas Ahmad,段 铭,关振宏,张茂林*
(1.吉林大学人兽共患病研究所 人兽共患病研究教育部重点实验室,吉林 长春 130062;2.吉林大学 动物医学学院,吉林 长春 130062)
狂犬病是由狂犬病病毒(Rabies virus,RABV)引起的一种高致死性的人兽共患传染病,发病动物伴有明显的神经症状,如:恐水、恐声、惊厥等[1]。经脑内及外周接种而感染RABV 的成年鼠,脑组织切片中能够观察到神经元突起的减少或轴突、树突呈串珠样改变和肿胀[2-3]。研究表明RABV 感染体外培养的原代神经元细胞能致使其树突棘数量减少,这一变化是由活性Ras 相关C3 肉毒素底物1(Rac1)含量的降低而引起[4]。由于微丝是维持树突棘结构的细胞骨架,而微管是细胞内物质运输及保持细胞形态的重要细胞骨架,其变化严重影响微丝的结构和功能。
EB 家族(Microtubule-associated protein RP/EB family)存在于微管正极端[5],其家族成员EB1 和EB3 能够抑制微管解聚,同时EB3 只在较成熟的脑内高表达,表明其具有组织特异性这一特点。在细胞骨架分子的信号网络中,研究RABV 株感染条件下EB3 的变化,有助于揭示细胞骨架动态变化及神经功能异常的机制[6]。为此,本研究应用RABV 固定株CVS 感染体外培养的神经元,研究EB3 转录、蛋白水平及其在细胞内分布的变化,为狂犬病神经功能异常的致病机制提供骨架方面的数据。
1 材料和方法
1.1 病毒株及实验动物 RABV 鼠脑研磨液由本实验室保存;SPF 级Wister 大鼠购自亿思实验动物中心,生产许可证号码:SCXK(吉)2011-0004。
1.2 主要试剂 Neurobasal medium 和B-27 Serum-Free Supplement(50×)购自Gibco 公司;抗EB3 兔单克隆抗体(MAb)和抗β-actin 兔MAb 购自CST 公司;FITC 标记的抗RABV N 蛋白MAb 购自Abcam 公司;TRIzol 试剂和Cy3 标记的山羊抗兔MAb 购自Invitrogen 公司:HRP 标记的山羊抗兔MAb 和Hochest 33258 细胞核荧光染色液购自Sigma 公司;PrimeScript RT reagent Kit with gDNA Eraser 购自TaKaRa 公司;FastStart Universal SYBR Green Master(Rox)购自Roche 公司。
1.3 引物设计及合成 根据GenBank 中登录的大鼠EB3 mRNA 序列(NM_00100765)设计一对引物,5'-GTGGACAAAATCATTCCCGTA-3'/5'-TTTCCATCA TAGTTTGCGTCA-3',其扩增产物为107 bp。
1.4 原代神经元细胞的培养 对细胞培养板及玻璃爬片经多聚赖氨酸预处理。取胎鼠脑,将收集海马及皮层并消化分离细胞,采用含2 % B27、1 % 谷氨酰胺、1 % 双抗的Neurobasal medium 重悬细胞并进行铺板。3 d 之后加5 μmol/L 的阿糖胞苷,6 d 之后更换50 %新鲜培养基[7]。10 d 时,按10 TCID50浓度接种病毒鼠脑研磨上清液。分别按时间点收集样品。以未接毒细胞为对照组。
1.5 病毒感染细胞荧光定量PCR检测 总RNA 提取步骤参照TRIzol Reagent 说明书,并测定其浓度。采用反转试剂盒制备cDNA。对样品进行荧光定量PCR 检测,反应条件及体系参照说明书。
1.6 EB3表达变化的western blot检测 对原代神经元细胞进行病毒接种,培养后收集细胞并提取蛋白。测定其浓度并调平。SDS-PAGE 电泳后转膜,经含5 % 脱脂奶粉的封闭液室温封闭2 h,分别以兔抗EB3 MAb(1∶1 000)和兔抗β-actin MAb(1∶1 000)为一抗,以HRP 标记的山羊抗兔MAb(1∶1 000)为二抗,显色底物反应3 min,经化学发光呈像系统进行图像采集。
1.7 EB3细胞内分布的间接免疫荧光试验(IFA)取24 孔板中的病毒接种的细胞爬片经4 %多聚甲醛固定及0.3 % Triton-X 处理。经5 % 灭活羊血清的PBS 封闭。以抗EB3 兔MAb 和FITC 标记的抗RABV N 蛋白MAb(1∶100)为一抗,Cy3 标记的山羊抗兔MAb(1∶100)为二抗与Hochest 33258 细胞核荧光染色液(10 μg/mL)室温同时作用2 h。利用抗荧光猝灭剂封片,通过激光共聚焦显微镜采集图像。
1.8 数据分析 荧光定量PCR 检测结果经2-△△Ct法进行处理。Western blot 结果采用Image J 采集灰度值,进行蛋白含量变化倍数的计算。目的蛋白的变化倍数=(实验组目的蛋白灰度值×蛋白上样量/实验组内参灰度值)/(对照组目的蛋白灰度值×蛋白上样量/对照组内参灰度值)。所得结果再经SPSS 软件统计分析,各组间差异采用单因素方差分析,所得结果以表示,p<0.05 表示差异具有统计学意义。
2 结果
2.1 病毒感染对原代神经元细胞EB3基因转录的影响 对实验组及对照组分别收取RABV 感染0 h、6 h、12 h 及48 h 样品,进行荧光定量PCR 检测(X±SD,n=5)。结果表明,感染病毒6 h 后,与对照组相比,EB3 的转录水平下降明显。在细胞感染12 h 之后,EB3 的转录水平进一步下降。在感染24 h之后,原代神经元细胞内EB3 基因的表达水平有所回升,至48 h 时进一步回升但仍然低于对照组,表明RABV 可以在转录水平上影响EB3 的表达(图1)。
2.2 病毒感染对神经元EB3表达的调节作用 对实验组及对照组分别收取0 h、6 h、12 h 及48 h 样品,进行western blot 检测,采用Image J 对EB3 的蛋白含量进行分析结果显示,病毒可导致β-actin 含量有一定程度的减少(图2)。本实验采用EB3 的变化倍数进行比较。结果显示,一方面,对照组中48 h 与96 h 相比,随着培养时间的延长,EB3 比例明显增高。另一方面,实验组中病毒感染1 h 之后,EB3 含量减少,随着感染时间的延长,EB3 的比例逐渐下降。感染48 h 之后,与对照组相比,EB3 比例明显减少,含量几乎消失。结果表明,随着感染时间的延长,EB3 含量不断降低,病毒感染能够抑制EB3 的表达(图3)。
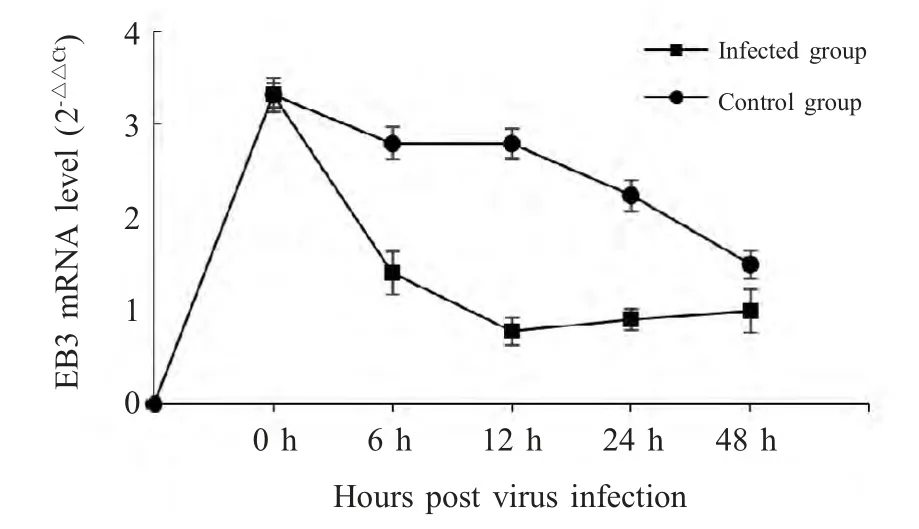
图1 病毒感染对神经元内EB3 mRNA 水平的影响Fig.1 The influence of virus infection to mRNA level of EB3 in neuron

图2 病毒感染神经元细胞的EB3 表达水平Fig.2 Protein level of EB3 in virus infected neuron

图3 病毒感染对神经元EB3 表达的影响Fig.3 The influence of virus infection to protein level of EB3 in neuron
2.3 病毒感染对原代神经元细胞内EB3的分布调节作用 对实验组及对照组分别收取48 h 与96 h细胞爬片。结果显示,48 h 对照组神经元细胞内,EB3 在神经突上呈索状均匀分布,在胞体内均匀分布于胞浆中,并于细胞膜附近集中分布,形成较亮的着色圈;在96 h 对照组中,神经元突起明显增多,EB3 仍然均匀分布于胞体及突起上,胞膜附近仍然有EB3 的聚集。而在感染病毒48 h 后,神经元胞体内有少量病毒光斑,EB3 荧光点在突起处较连续;神经元感染96 h 后,在胞体及突起上均能看到绿色光斑,红色光斑杂乱地分布于突起上及胞体细胞膜内侧,且变得不连续(图4)。

图4 病毒感染对神经元内EB3 分布的影响Fig.4 Distribution changes of EB3 in neuron after virus infection
3 讨论
EB3 是微管正极端标记蛋白EB 家族的成员,属于结构相对保守的二聚蛋白,参与调节微管的动态变化,促进肌细胞的延长[8],参与细胞间的黏附及轴突的生长等[9]。EB3 的N 端能够与微管相互作用。C 端能够与p140cap 和APCL 相互结合[5-6]。实验证明,EB3 在神经元内作为微管结合蛋白,能够使p140cap 结合于微管末端,从而通过调节Rac1 而影响树突棘的可塑性[6]。所以,EB3 蛋白不仅可作为微管正极端蛋白以稳定微管,而且能够在微管缺失的树突棘内通过调节p140cap 调节actin 的聚合状态,从而影响树突棘的形态,为微管和微丝之间相互联系的纽带。
本实验从微管正极端标记蛋白家族成员EB3 出发,探素RABV 感染对神经元微管的影响。本研究表明,当RABV 感染神经元后,通过荧光定量PCR检测,可以检测感染6 h 之后,EB3 基因的转录水平不断下降。在48 h 后,转录量有所回升,这可能是由于随着RABV 感染时间的延长,神经细胞因成熟或反馈性修复引起EB3 基因mRNA 水平回升。将感染病毒后EB3 的转录水平和蛋白水平进行比较,在感染48 h 之后,mRNA 水平有所上升,而蛋白水平明显下降,两者变化趋势不一致。这种转录调控和翻译调控对EB3 表达调节方向的不一致[9],可能是由于RABV 感染对神经细胞内EB3 的调节作用以翻译调控为主有关,进而改变其细胞内定位,影响神经元突起的形态。Jaworski 和Di 等分别研究证明EB3 在原代神经元细胞中能够影响p140cap 在细胞骨架近膜部分的定位及p140cap 在肿瘤细胞中能够对附近Rac1 活性进行调节[6,10]。再结合本实验结果,推测RABV 对神经元内Rac1 的活性也具有潜在的影响。因此,本实验对病毒感染后Rac1 活性也进行了检测(相关数据另文发表),发现病毒感染也能够抑制神经元内Rac1 的活性,从而影响到微丝的重排。
本实验室前期研究表明RABV 街毒株能够破坏神经元微丝,从而影响神经元的形态。所以本研究综合其他关于EB3 对神经元形态的影响及RABV 对神经元微丝的影响两方面研究成果,从微管角度出发,证明了病毒能够通过抑制微丝的上游微管相关蛋白,影响神经元微管的稳定及微丝的重排,最终改变神经元的形态,而其对神经元功能的影响还需要进一步的研究。本实验丰富了RABV 对神经元细胞骨架影响的实验数据。同时,为进一步对RABV街毒株引起的神经元退行性病变机制研究提供相关的实验依据。
[1]扈荣良,张守峰,刘晔.我国狂犬病预防和控制建议[J].中国人兽共患病学报,2012,28(5):487-491.
[2]Feng Guo-ping,Mellor R H,Bernstein M,et al.Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP[J].Neuron,2000,28(1):41-51.
[3]Scott C A,Rossiter J P,Andrew R D,et al.Structural abnormalities in neurons are sufficient to explain the clinical disease and fatal outcome of experimental rabies in yellow fluorescent protein-expressing transgenic mice[J].J Virol,2008,82(1):513-521.
[4]宋艳.狂犬病病毒街毒株感染致突触F-actin 重构及其机制的研究[D].长春:吉林大学,2013.
[5]Komarova Y,De Groot C O,Grigoriev I,et al.Mammalian end binding proteins control persistent microtubule growth[J].J Cell Biol,2009,184(5):691-706.
[6]Jaworski J,Kapitein L C,Gouveia S M,et al.Dynamic microtubules regulate dendritic spine morphology and synaptic plasticity[J].Neuron,2009,61(1):85-100.
[7]Beaudoin III G M J,Lee S H,Singh D,et al.Culturing pyramidal neurons from the early postnatal mouse hippocampus and cortex[J].Nat Protoc,2012,7(9):1741-1754.
[8]Zhang Tan,Zaal K J M,Sheridan J,et al.Microtubule plus-end binding protein EB1 is necessary for muscle cell differentiation,elongation and fusion[J].J Cell Sci,2009,122(9):1401-1409.
[9]Garcia-Sanz J A,Mikulits W,Livingstone A,et al.Translational control:a general mechanism for gene regulation during T cell activation[J].FASEB J,1998,12(3):299-306.
[10]Di Stefano P,Damiano L,Cabodi S,et al.p140cap protein suppresses tumour cell properties,regulating Csk and Src kinase activity[J].EMBO J,2007,26(12):2843-2855.
猜你喜欢
中国骨质疏松杂志(2022年9期)2022-10-18
中老年保健(2022年1期)2022-08-17
——水芹主要害虫识别与为害症状
长江蔬菜(2022年13期)2022-07-29
昆明医科大学学报(2021年8期)2021-08-13
土壤与作物(2021年2期)2021-06-01
小读者之友(2020年4期)2020-05-15
中国癌症防治杂志(2019年5期)2019-01-04
中成药(2018年9期)2018-10-09
猪业科学(2018年8期)2018-09-28
中国实验动物学报(2017年2期)2017-05-18
