
盐酸埃克替尼对人肺腺癌细胞系PC-9作用的比较蛋白质组学研究
2015-03-08 12:50刘晓丽张玉萍
中华肺部疾病杂志(电子版) 2015年6期
刘晓丽 李 维 孟 夏 张玉萍
明宗娟 史红阳 石 婕 钟玉洁
王 薇 杨拴盈
·论著·
盐酸埃克替尼对人肺腺癌细胞系PC-9作用的比较蛋白质组学研究
刘晓丽李维孟夏张玉萍
明宗娟史红阳石婕钟玉洁
王薇杨拴盈
【摘要】目的应用比较蛋白质组学方法分析盐酸埃克替尼作用前后人肺腺癌细胞系PC-9细胞相关蛋白的变化,从蛋白质水平探讨埃克替尼的作用机制。方法应用人类EGFR基因突变检测试剂盒和实时定量PCR仪检测PC-9细胞EGFR突变;应用MTT法检测埃克替尼对PC-9细胞增殖的影响;应用双向凝胶电泳(2-DE)技术分别对未经干预的PC-9细胞总蛋白和经埃克替尼作用的PC-9细胞总蛋白进行分离,获得蛋白质表达谱;应用基质辅助激光解吸电离飞行时间质谱(MALDI-TOF-MS)结合生物信息学进行蛋白质鉴定。结果PC-9细胞的EGFR基因存在第19外显子缺失突变;在10-4-10-9M浓度范围内,埃克替尼明显抑制PC-9细胞的生长,且呈浓度依赖;埃克替尼诱导前后表达水平≥1.5倍的蛋白点共有56个。选取差异≥1.7倍的29个蛋白点进行MALDI-TOF-MS分析,最终鉴定出16种蛋白质。根据功能,这些蛋白可分为代谢酶类、细胞骨架类、分子伴侣、信号传导分子、转录及翻译相关蛋白。结论应用比较蛋白质组学方法初步鉴定了16种蛋白质,这些蛋白质与埃克替尼作用机制密切相关,部分可能成为新的分子治疗靶点。
【关键词】PC-9细胞;埃克替尼;比较蛋白质组学;生物信息学
肺癌是目前发病率及病死率最高的恶性肿瘤[1-2]。非小细胞肺癌(non-small cell lung cancer, NSCLC)约占肺癌发病总数的80%以上。其5年生存率仅15%。对于早期NSCLC,目前首选手术治疗,晚期NSCLC的治疗是以化疗为主的综合治疗。然而传统的化疗和放疗虽然可暂时缓解症状,使肿瘤缩小,但并不能有效提高生存期,且常因放化疗的毒副反应,使患者无法耐受治疗。表皮生长因子受体酪氨酸激酶抑制剂(epidermal growth factor receptor tyrosine inhibitor, EGFR-TKI)是目前抗癌药研究中最受瞩目的领域[3]。埃克替尼是一种小分子EGFR-TKI,由浙江贝达药业有限公司自主研发。对于EGFR基因突变的患者,盐酸埃克替尼具有明显的抗肿瘤作用[4]。目前针对该药作用机制的研究多集中在PI3K/AKT、Ras/Raf/MEK/MAPK、JAK/STAT3信号通路,其它作用机制尚不明确。比较蛋白质组学是蛋白质组学的重要组成部分,它不仅在探讨发病机制、早期诊断等方面发挥巨大的作用,而且可以通过研究药物处理前后蛋白的差异变化,深入探讨药物作用机制,并发现新的、潜在的药物作用靶点。本研究通过比较蛋白质组学方法观察埃克替尼作用前后相关蛋白变化,以期从蛋白质水平探讨埃克替尼作用机制,并为开发新靶向治疗药物的提供理论依据。
材料与方法
一、实验材料
1. 细胞: 人肺腺癌细胞系PC-9细胞购自上海复祥生物科技有限公司,用含10%胎牛血清的DMEM高糖培养基进行传代培养。
2. 主要仪器: 细胞培养箱(美国Thermo公司);超净工作台(天津泰斯特仪器有限公司);倒置显微镜(日本Nikon公司);超声细胞破碎仪(浙江新芝生物科技有限公司);PowerWave XS 酶标仪(Bio-tek公司);ABl7500实时定量PCR仪(美国应用生物系统公司):Ettan IPGphor Isoelectric Focusing System(GE Amersham公司);光密度扫描仪GS-710(美国Bio-Rad公司);凝胶图像分析软件Image Master(GE Amersham公司);4800 MALDI-TOF质谱仪(AB SCIEX);UMax Powerlook 2100XL扫描仪(GE Amersham公司);Hofer SE 600(GE Amersham)。
3. 试剂: 盐酸埃克替尼由浙江省贝达药业有限公司馈赠;DMEM高糖培养基、胎牛血清、胰蛋白酶溶液均购自美国Hyclone公司;MTT、DMSO均购自美国Sigma公司;Amoy Dx试剂盒购自厦门艾德生物医药科技有限公司;DNA提取试剂盒购自日本Takara公司;PH3-10非线性胶条、PH3-10L IPG Buffer均购自美国Pharmacia公司;考马斯亮蓝G-250/R-350、Urea、CHAPS、DTT、IAA、Tris均购自美国GE Amersham公司;Bradford蛋白定量试剂盒、SDS、APS、TEMED、甘氨酸、甘油均购自美国Bio-Rad公司;Cocktail酶抑制剂购自美国Roche公司;氯化钠、氯化钾、十二水合磷酸氢二钠、磷酸氢二钾、乙醇、乙酸、戊二醛、甲醛、柠檬酸均为国产分析纯试剂。
二、实验方法
1. 细胞EGFR基因突变检测: 将处于对数生长期的细胞,胰酶消化,离心重悬后提取DNA,所有步骤按培养贴壁细胞DNA提取操作说明;利用人类EGFR基因突变检测试剂盒(Amoy Dx)和实时定量PCR仪检测突变基因。
2. 细胞增殖抑制率测定: 取生长状态稳定、对数生长期的细胞,接种至96孔板中,使每孔细胞数为5 000个左右。过夜细胞贴壁后,更换培养液,加入盐酸埃克替尼,使各标记孔内药物浓度分别为100 μmol/L、10 μmol/L、1 μmol/L、0.1 μmol/L、0.01 μmol/L、0.001 μmol/L,每个浓度组设6个复孔,对照组加入等量培养液。继续孵育72 h,每孔加5 mg/ml的MTT溶液20 μl,孵育4 h弃上清,每孔加入二甲基亚砜(dimethyl sulfoxide, DMSO) 150 μl,振荡10 min,使结晶物充分溶解;将培养板放入酶联免疫检测仪,测量并记录各孔的光吸收值。癌细胞存活率由下式计算:细胞存活率=(实验组光吸收值/对照组光吸收值)×100%,通过SPSS12.0统计软件,将实验组与对照组作t检验。确定埃克替尼作用PC-9细胞72 h的半抑制浓度(IC50);以抑制率为纵坐标、药物浓度为横坐标绘制曲线图。
3. 细胞总蛋白的提取: PC-9细胞生长至对数生长期后,更换培养基,使实验组细胞暴露于IC50的埃克替尼中,继续孵育72 h。吸出两组细胞的培养液,用预冷的PBS缓冲液清洗三遍,将PBS吸除干净,加入250 μl细胞裂解液,每10 min充分混匀一次,于冰上裂解细胞1 h,用刮刀收集细胞。冰浴超声裂解(80 w,5次,每次10 s,间隔15 s,冰上完成),超声后离心(12 000×g、45 min),4 ℃,取上清;Bradford法蛋白定量,分装,-80 ℃冰箱保存。
4. 2-DE: 将样品100 μg与重泡涨液充分混合,低速离心后,加入持胶槽,将IPG干胶条(PH3-10L,13 cm)去保护膜,胶面朝下从一端放入持胶槽中,使重泡涨液浸润整个胶条,用矿物油覆盖,操作过程中注意不要产生气泡;水化和等电聚焦在20 ℃自动进行,设置参数如下:30 v 12 hrs,500 v 1 hrs,1 000 v 1 hrs,8 000 v 8hrs,500v 4hrs;将胶条进行两步平衡后,移至12.5%分离胶的上端,进行十二烷基磺酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfatepolyacrylamide gelelectrophoresis, SDS-PAGE)。电泳完成后,按蛋白质银染试剂盒的操作手册进行银染。
5. 凝胶图像分析: 两组样品常规各跑3次双向凝胶电泳,GS-710光密度扫描仪对染色后的胶图进行扫描,通过Image-Master软件对两组图像进行背景消减、蛋白点检测、匹配、量化及数据分析。应用SPSS12.0软件进行统计学分析,采用t检验。选取差异1.7倍以上的蛋白质点进行质谱分析。
6. 质谱分析: 将银染蛋白点进行胶内酶切,脱色,还原,烷基化,酶解,萃取,冻干。将制备好的样品用串联飞行时间质谱仪(4 800 Plus MALDI TOF/TOFTM Analyzer)进行测试分析,激光源波长为355 nm,离子源加速电压为2 kV,采用正离子模式和自动获取数据的模式采集数据,MS扫描范围为800~4 000 Da,选择信噪比大于50的母离子进行MS/MS分析,每个样品点上选择8个母离子,二级质谱(MS/MS)累计叠加2 500次,碰撞能量2 kV,CID关闭;质谱测试原始文件用Mascot2.2软件检索相应数据库,最后得到鉴定的蛋白质结果。
7. 生物信息学分析: 利用NCBI数据库对差异表达明显且与肿瘤密切相关蛋白进行mRNA、蛋白质序列进行检索,在此基础上利用ProtParam、SOPMA、TMpred、PSORT、SWISS-MODEL、SMART、STRING、KEGG等工具对它们的理化性质、二级结构、跨膜结构域、亚细胞定位、三维结构、功能结构域、蛋白相互作用、信号通路等进行初步分析。
结果
一、EGFR基因表达情况
采用ADx-ARMS法对人肺腺癌细胞系PC-9细胞的EGFR基因18、19、20、21外显子进行突变检测。结果显示PC-9细胞的EGFR基因存在第19外现在缺失突变。
二、细胞增殖抑制率
药物干预72 h后通过倒置显微镜可观察到贴壁细胞生长速度减慢,形态改变,出现细小颗粒,并可见细胞碎片。不同浓度埃克替尼作用于PC-9细胞72 h后,MTT法检测细胞OD值并计算抑制率。结果发现在10-4-10-9M浓度范围内,埃克替尼明显抑制PC-9细胞的生长,且呈浓度依赖性,72 h IC50约为80 nmol/L,见图1。
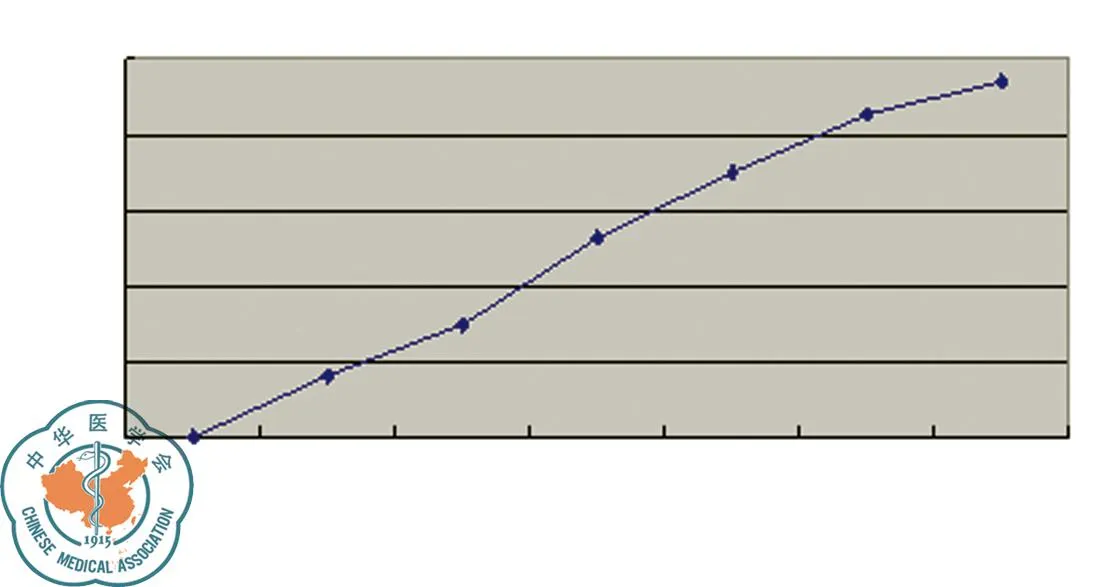
图1 不同浓度埃克替尼对PC-9细胞生长的影响
三、2-DE凝胶图谱
每组蛋白常规跑3次电泳,用GS-710光密度扫描仪对胶图进行扫描,采用Image-master软件进行分析,结果显示对照组3块胶的平均蛋白斑点数为1 410±52,平均匹配率为63.45%;实验组的平均蛋白斑点数为1 480±52,平均匹配率为66.61%。对照组和实验组间蛋白差异≥1.5倍的点共有56个。图2为对照组和实验组间蛋白差异点。

图2 对照组和实验组间蛋白差异点凝胶图谱
四、质谱分析结果
选取差异≥1.7倍的29个蛋白点进行MALDI-TOF-MS分析,最终成功获得16种蛋白的PMF,其中埃克替尼干预后下调的蛋白有13种,分别为细胞角蛋白9(cytokeratin9, CK9)、吡咯琳-5-羧酸还原酶(pyrroline-5-carboxylate reductase, PYCR)、ran特异性GTP酶活化蛋白(ran-specific GTPase-activating protein, RANBP1)、肿瘤蛋白18(stathmin1/oncoprotein18, STMN1)、NEM1-NEM2 蛋白、rasGTP酶激活蛋白结合蛋白1(ras GTPase-activating protein-binding protein1, G3BP1)、泛素结合酶E2D3(ubiquitin-conjugating enzyme E2D3, UBE2D3)、微管蛋白α-1A链亚型2(tubulin alpha-1A chain isoform2, TUBA1A isoform2)、 视黄醇脱氢酶(retinol dehydrogenase, RDH)、热休克蛋白40(DnaJ homolog, subfamily B, member1, DNAJB1)、不均一核糖核蛋白A/B(heterogeneous nuclear ribonucleoprotein A/B, hnRNPA/B)、电压依赖性阴离子选择性通道蛋白1(voltage-dependent anion-selective channel protein1, VDAC1)、真核翻译起始因子3亚组1(eukaryotic translation initiation factor 3 subunit1, EIF31);加药后上调的蛋白点有3种:层黏连蛋白结合蛋白(laminin-binding protein, RPSA)、葡萄糖-6-磷酸脱氢酶(glucose-6-phosphate dehydrogenase, G6PD)、果糖-1,6二磷酸酶(fructose-1,6-bisphosphatase, FBPase),见表1,图3。
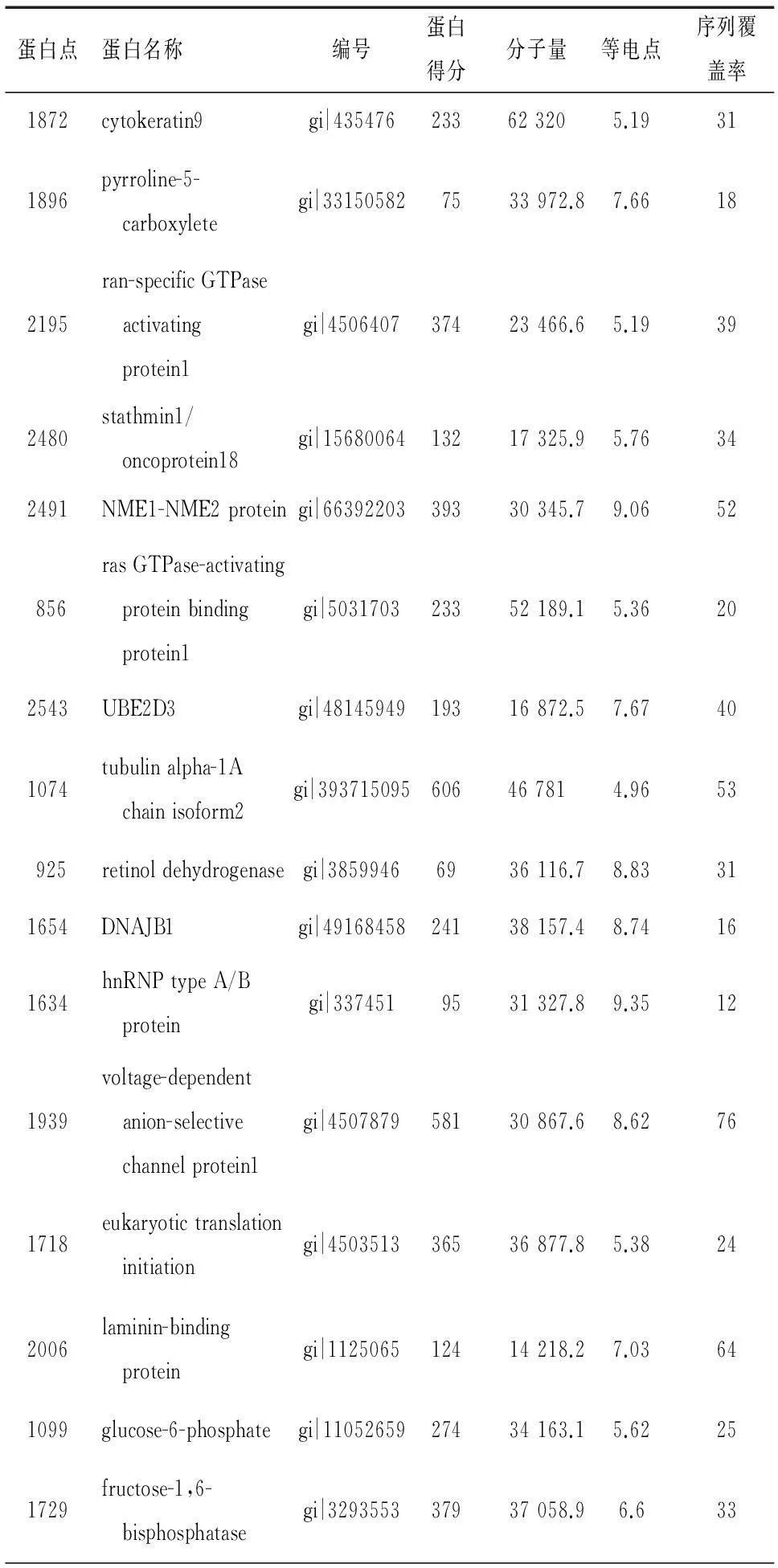
表1 差异表达蛋白质谱分析结果
图3 1872点的一级肽质量指纹图谱
五、生物信息学结果
查阅相关文献,对差异表达明显且与肿瘤密切相关的NME1-NME2、STMN1、G3BP1蛋白进行生物信息学分析。NME1-NME2蛋白由267个氨基酸构成,相对分子质量约为3 0137.0,理论等电点9.06,是一种可溶性、亲水性蛋白质;其二级结构主要有α-螺旋(37.83%)和无规则卷曲(26.97%);TMpred结构域分析软件显示NME1-NME2蛋白有2个可能的跨膜结构域;SMART数据库搜索结果提示该蛋白有2个NDK结构域;亚细胞定位结果显示NME1-NME2可能存在于细胞质中;图4为NME1-NME2蛋白质的三维结构图;STRING库预测所得与NME1-NME2相互作用蛋白得分高的主要有KCNN4、CCND1、PAPD4、ACLY、MYC、CNOT8、HSP72、NME1、EEF1A1,另外,NME1-NME2可能与STAT3通路有一定的关联(图5)。
图4 NME1-NME2蛋白的三维结构图
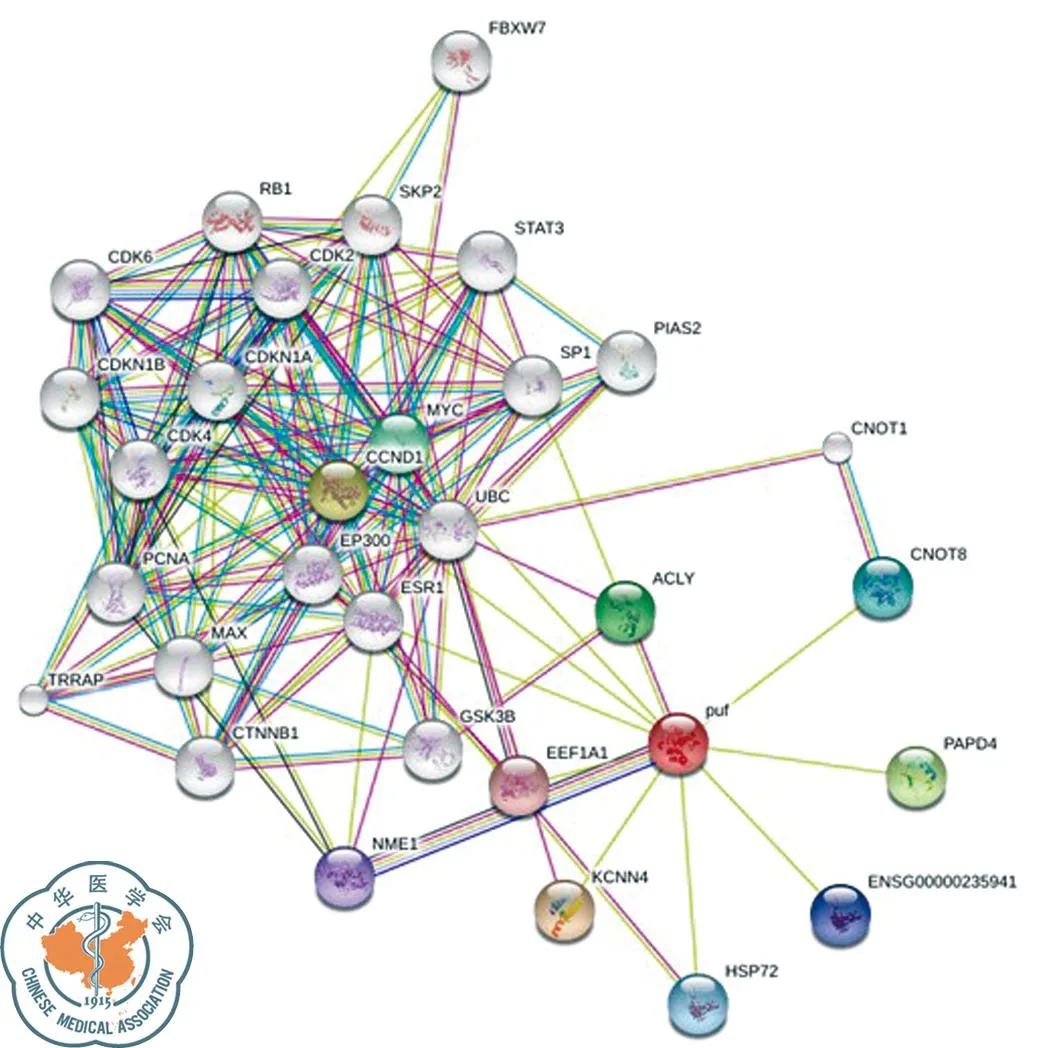
图5 STRING数据库:NME1-NME2相互作用蛋白预测结果
STMN1基因有四个mRNA转录变本,转录变本1、2、3翻译合成同一种蛋白质stathmin亚型a,转录变本4翻译合成stathmin亚型b;stathmin亚型a由149个氨基酸组成,相对分子质量为17 302.5,理论等电点5.75;stathmin亚型b由174个氨基酸组成,相对分子质量为19 823.5,理论等电点为6.52;两种蛋白均为可溶性、亲水性蛋白;stathmin亚型a二级结构主要有α-螺旋(78.52%)和无规则卷曲(18.79%);stathmin亚型b二级结构主要由a螺旋(71.84%)和无规则卷曲(21.84%)构成;TMpred软件显示stathmin亚型a多肽链可能并不存在跨膜结构域,stathmin亚型b有1个可能的跨膜结构域; stathmin亚型a和亚型b均含有一个stathmin结构域;亚细胞定位显示STMN1存在于细胞质和细胞骨架中;图6为STMN1蛋白的三维结构图;与STMN1蛋白相互作用密切的有AURKB、MAPK3、MAPK13、CDK1、TP53、MAPK1、CAMK2B、CAMK2G、NACA、ADCYAP1,可见其参与MAKP信号通路、细胞周期调控,并与TP53密切相关(图7)。

图6 STMN1蛋白的三维结构图
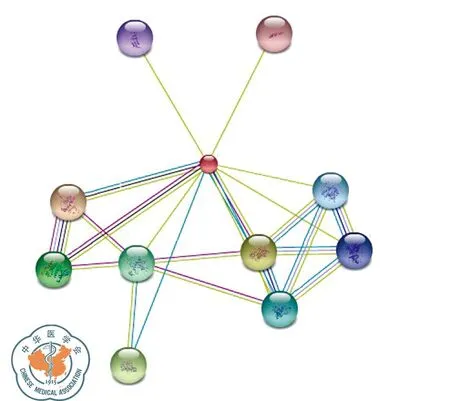
图7 STRING数据库:STMN1相互作用蛋白预测结果
G3BP1:该蛋白质由466个氨基酸构成,相对分子质量为52 164.2,理论等电点为5.36;二级结构主要有a-螺旋(23.82%)和无规则卷曲(55.79%);没有跨膜结构域,是亲水性、可溶性蛋白;G3BP1蛋白含有NTF2和RRM结构域;G3BP1分布广范,主要存在于细胞质中;图8为G3BP1蛋白质的三维结构;与G3BP1蛋白相互作用密切的主要有CAPRIN1、USP10、EIF4G1、RASA1、CSK、HDAC6、OGFOD1、PRMT1、STYXL1、ALPP,是ras信号转导通路中的一个重要元素(图9)。

图8 G3BP1蛋白的三维结构图
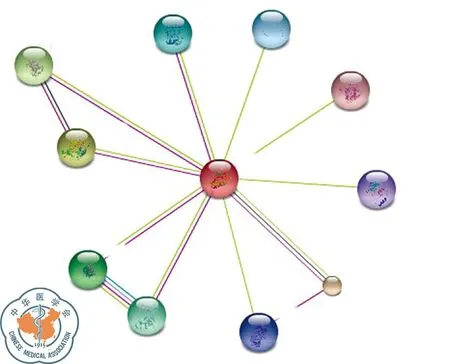
图9 STRING数据库:G3BP1相互作用蛋白预测结果
讨论
肺癌的发生、发展是一个多基因参与,多分子调控的复杂过程,且早期缺乏特异性症状,发病机制不明,容易发生远处转移,预后极差。EGFR-TKI能特异性、竞争性地结合在EGFR激酶功能区的ATP结合位点,抑制其激酶活性从而阻断癌细胞增殖、转移等相关信号传导通路,已成为肺癌治疗研究中的热点。国外上市的EGFR-TKI包括吉非替尼和厄洛替尼,其主要适应症为二线或三线用于其它抗癌药物治疗失败的晚期NSCLC患者和用于EGFR基因突变的NSCLC患者的一线治疗。盐酸埃克替尼是我国自主设计、合成、筛选出的具有自主知识产权的EGFR-TKI。大量临床前及临床数据表明埃克替尼能够明显抑制肿瘤细胞的生长,且毒副作用较易瑞沙、特罗凯明显减少。
EGFR基因敏感突变是EGFR-TKI分子靶向治疗发挥疗效的关键预测因素。常见敏感突变主要有G719X、Exon19 deletions、L858R、L861Q、S768I突变,耐药突变包括单独的T790M突变和外显子20插入突变等。人肺腺癌细胞系PC-9经ADx-ARMS法检测提示存在exon19 deletion突变,其余exon 18(G719X)、exon 20(S768I、Insertion、T790M)、exon 21(L858R、L861Q)突变阴性,是对EGFR-TKI敏感的细胞系。本研究提示低浓度的盐酸埃克替尼就能明显抑制PC-9细胞的生长,其72 h IC50为80 nmol/L。既往研究表明吉非替尼72 h IC50接近50 nmol/L[5-6]。由此可见,埃克替尼IC50较吉非替尼IC50相近,是一种对NSCLC有治疗潜力的药物。
比较蛋白质组学技术已广泛应用于肿瘤发病机制、早期诊断及治疗的各个方面。目前对盐酸埃克替尼的研究多集中在PI3K/AKT、Ras/Raf/MEK/MAPK、JAK/STAT3信号通路方面,其它作用机制尚不明确。PC-9细胞系存在EGFR酪氨酸激酶区19外显子突变(E746-A750缺失),是目前对EGFR突变与肺癌的分子机制研究采用的主要细胞系之一。我们应用比较蛋白质组学方法,通过比较埃克替尼作用前后PC-9细胞蛋白差异表达的变化,发现了16种蛋白。根据功能,这些蛋白可分为代谢酶类、细胞骨架类、分子伴侣、信号传导分子、转录及翻译相关蛋白。
细胞结构的完整性和机械稳定性依赖于细胞骨架蛋白家族的维系。该蛋白家族包括微丝、中间丝及微管蛋白。角蛋白是中间丝家族的主要成员,表达于正常上皮细胞、部分非上皮细胞及上皮源性恶性肿瘤中。CK可分为Ⅰ型CK(酸性细胞角蛋白,CK9-20)和Ⅱ型(碱性细胞角蛋白,CK1-8),其结构由头、杆、尾三部分组成,杆部位于中间,其始端、末端均为高保守序列。所有上皮细胞至少表达一种Ⅰ型和一种Ⅱ型角蛋白单体(CK19例外),以独特的成对方式存在于真核细胞,并具有组织特异性。在肿瘤发生和转移时,肿瘤细胞多保留起源细胞的CK类型,对上皮组织的定性具有独特意义。细胞角蛋白在骨髓、血液及淋巴结中不表达,因此,被认为是检测循环癌细胞的敏感标记物[7],其中细胞角蛋白19片段已广泛应用于NSCLC的诊断及预后判断。既往研究表明CK7、CK8、CK20、CK18、CK19等与肿瘤发生、发展密切相关,而并未发现CK9与人类恶性肿瘤的特殊关联[8]。在本研究中,我们发现CK9在埃克替尼处理后的人肺腺癌细胞系PC-9中表达呈下调状态。另外,Fu等[9]应用差异蛋白质组学方法进行肝癌患者血清筛查时发现CK9在肝内转移的肝癌组血清CK9的表达水平也是上调的,提示CK9可能参与了埃克替尼抑制肿瘤细胞生长的过程。
自从1920年Warburg[10]发现肿瘤细胞即使在正常的氧气供应情况下,糖酵解依然增加并伴随大量乳酸生成,线粒体呼吸大幅减少这一现象后,人们普遍意识到代谢途经改变是细胞恶性转化必不可少的因素之一。研究证实脯氨酸代谢与血脯氨酸过多症、精神分裂症、皮肤松弛症及肿瘤有密切联系。在后者中,肿瘤细胞倾向于依赖脯氨酸的生物合成,其主要合成途经有两条:一条途径为以谷氨酸为底物合成脯氨酸,另一条途径为以鸟氨酸为底物合成脯氨酸。谷氨酸在吡咯琳合成酶作用下生成吡咯琳-5-羧酸,然后在PYCR作用下还原为脯氨酸。鸟氨酸在鸟氨酸转氨酶(ornithine aminotransferase, OAT)、PYCR作用下还原为脯氨酸。人类PYCR有三种同工酶:PYCR1位于染色体17q25.3;PYCR2定位于1q42.12;PYCRL基因定位于8q24.3。目前已证实PYCR在原发性及转移性肺肿瘤中明显升高[11],PYCR1参与乳腺肿瘤的发生,而对PYCR2、PYCRL的研究较少[12]。De Ingeniis等[13]通过13C同位素标记对黑素瘤细胞进行观察,首次提出PYCR1、PYCR2主要存在于线粒体中,介导谷氨酸转化为脯氨酸,而PYCRL存在于胞质中,参与鸟氨酸转化为脯氨酸的过程。
G蛋白作为一种信号蛋白,实质是一种GTP水解酶,可根据其为单体还是多聚体分为异三聚体G蛋白和小G蛋白[14]。异三聚体G蛋白由α、β、γ亚基组成,β、γ亚基以二聚体形式锚链于细胞质膜上,其可结合或解离α亚基。当α亚基结合GTP后,其构象发生改变,从二聚体上解离下来,与下游效应分子结合,从而控制相关信号通路。真核生物的小G蛋白是单体蛋白分子,又可以分为5个家族:Ras、Rab,Rho,Arf和Ran。Ran是小G蛋白的一个亚族,具有GTP水解酶活性,分子量约为(20~30)×103,与其它小G蛋白一样,在信号传导通路中起“分子开关”的作用。Ran蛋白活性主要受Ran鸟苷酸交换因子(Ran guanine exchang factor, RanGEF)RCC1、Ran结合蛋白(Ran binding proteins, RanBPs)、RanGTPase激活蛋白(RanGTPase activator protein, RanGAP)调控。RANBP1位于细胞质中,它可以抑制RCC1活性,间接促进RanGTP水解为RanGDP的过程,在细胞的核质运输、纺锤体组装、细胞周期、控制有丝分裂和有丝分裂后的核重组中有一定的调节作用[15]。
抑微管装配蛋白(Stathmin 1, STMN1),又被称为p17, p18, p19, 19K, 肿瘤蛋白18(oncoprotein18, Op18)及LAP18,是一种分子量为19kDa,存在于细胞质中的微管不稳定蛋白。它主要调节微管聚合、解聚的动态过程,促使微管在细胞周期的间期及有丝分裂后期解聚。在细胞周期进展的不同阶段Stathmin通过自身磷酸化水平调节细胞微管系统的动力学平衡,并以此改变细胞的增殖、分化、活性等生物学行为[16]。研究发现,Stathmin为多种细胞信号传导通路的中继站,在人类肺癌、急性白血病、前列腺癌、乳腺癌等恶性肿瘤均可检测到Stathmin的高表达[17-21]。另外,其mRNA的高表达水平亦与紫杉醇、长春瑞滨耐药性密切相关[22]。由此可见,stathmin有望成为治疗恶性肿瘤的新药物作用靶点[23]。
NM23基因(non-metastatic 23,也称为NME基因)是人类发现的第一个抑制肿瘤转移的基因,其表达与多种肿瘤转移潜能病理学指标、淋巴结浸润及不良预后密切相关[24]。目前已发现NME基因家族中有10个成员,根据基因组结构及磷酸转移酶活性不同可分为两组:Ⅰ组和Ⅱ组,Ⅰ组主要包括NME1,NME2,NME3,NME4基因(亦即NM23-H1~H4),这些基因具有较高的同源性,分别编码具有二磷酸核苷激酶(nucleoside diphosphate kinase, NDK)活性蛋白NDPK-A~D;Ⅱ组成员为NME5、NME6、NME7、NME8、NME9基因,编码不具有或具有较低NDPK活性的蛋白质。其中NME10的分组目前仍颇受争议[25]。在NME10个家族成员中,NME1、NME2亚型与肿瘤发展、转移相关性最为密切。另有相关研究表明,NME1、NME2与肺癌转移呈负相关[26]。
G3BP是一个在真核生物进化过程中高度保守的RasGAP(ras GTPase-activating protein) 结合蛋白[27]。它是由3.3kb的mRNA编码,分子量为68×103的蛋白质,主要定位于细胞质中。宁钧宇等[28]通过原核表达G3BP蛋白诱导免疫制备抗G3BP单克隆抗体,应用ELISA、蛋白免疫印迹、免疫组化等方法进行抗体鉴定和肿瘤检测,发现在结肠癌、肺癌、胃癌、乳腺癌组织中,G3BP均明显表达。另外还发现,G3BP2的过表达在原位癌形成的早期就已经出现[29];G3BP可通过与p53 C-端结合,致使p53不能形成多聚体,阻碍p53活性位点的转录后修饰,影响其活性,导致p53不能发挥正常功能[27];G3BP与抑癌基因PTEN的表达量相反,G3BP高表达引起的PTEN表达下调可能是肿瘤发生的机制之一[30]。已有研究表明G3BP N-端单克隆抗体具有良好的抗肿瘤效应。由此可见,G3BP与肿瘤的发生、发展、侵润及转移等关系密切,G3BP有望成为肿瘤治疗的新靶点。
泛素(Ubiquitin)是一种高度保守的小蛋白,存在于大多数真核细胞中,其主要功能为标记需要分解掉的蛋白质。参与泛素化过程的酶类主要有三种:泛素激活酶(ubiquitin-activating enzymes, E1s)、泛素结合酶(ubiquitin-conjugating enzymes, E2s)、泛素蛋白连接酶(ubiquitin-protein ligases, E3s)。泛素通过三种酶相互作用与目标蛋白结合,引起靶蛋白的降解。在恶性肿瘤中,泛素-蛋白酶体途经发生异常是细胞获得无限增殖性及永生性的重要原因,一些癌基因产物、肿瘤抑制因子p53、细胞周期蛋白等均通过此途经降解[31]。本研究显示UBE2D3在埃克替尼作用后PC-9细胞中下调,说明埃克替尼可能通过UBE2D3促进肺腺癌细胞凋亡、抑制其增殖。
微管是真核生物细胞骨架的重要组成成分,由α、β微管蛋白异源二聚体及微管辅助蛋白组成,参与维持细胞形态、有丝分裂、染色体分离、细胞内物质转运等过程,其不断延伸和缩短的特有动力学,在细胞有丝分裂和染色体分离中起关键的作用[32]。当微管蛋白与紫杉醇、长春瑞滨、多西他赛结合后,微管动力学受到影响,使有丝分裂的纺锤体形成障碍,并启动细胞固有死亡程序,从而抑制肿瘤的生长。TUBA1A亚型2是由TUBA1A基因的转录变体3翻译合成的。目前关于NSCLC与微管蛋白的研究较多的为βⅢ-tubulin,a-tubulin研究较少。Chen等[33]应用免疫组织化学法对158例原发NSCLC和30例癌旁组织中的微管蛋白α进行检测,发现微管蛋白α的表达率高达65.2%,而癌旁组织未见表达。另外,在肿瘤进展期,其表达量亦逐渐增加,在Ⅲ-Ⅳ期肺癌患者和低分化肿瘤中明显升高。在体外研究中发现a-tubulin表达参与紫杉醇耐药机制。Han等[34]对紫杉醇耐药的H460/T800细胞系进行观察,发现降低a-tubulin的表达能够明显增强细胞系对紫杉醇等抗纺锤体药物的敏感性。由此可见a-tubulin是影响化疗耐药的因素之一。
维甲酸(retinoic acid, RA)或全反式维甲酸(all-trans retinoic acid, atRA)是人体内重要的具有“激素样”生理功能的物质,它是维持胚胎发育中器官及正常结构形成的重要物质,并在抑制细胞生长、增殖,诱导细胞分化、凋亡及调节免疫等方面具有广泛的生物学效应。维甲酸受体目前认为主要有两种:RAR和RXR,其作用机制主要通过与核内受体结合影响转录来实现。动物实验研究中发现在致癌因素作用下,维甲类能够阻止动物模型皮肤、口腔、肺、膀胱肿瘤的发生[35-38]。另外,大量实验及临床研究亦发现维甲酸可阻止或逆转人类细胞、组织的恶性转化,如血液系统恶性肿瘤及肝脏、皮肤、肺脏、乳腺等原位癌[39-40]。维生素A也称为视黄醇,经过两步生物化学反应生成维甲酸,其中视黄醇脱氢酶为限速酶,显示出较强的视黄醇氧化和视黄醛还原活性。本实验中,视黄醇脱氢酶明显下调。综上所述,维甲酸及视黄醇脱氢酶可能成为极具潜力的靶标。
热休克蛋白(heat shock proteins, HSPs),是一类分布广泛的在进化过程中高度保守的蛋白质,它在细胞新陈代谢、免疫过程、细胞周期调节、转录激活及信号传导等方面发挥重要作用。根据分子量大小,HSPs被分为6个主要家族:HSP100、HSP90、HSP70、HSP60、HSP40和小分子HSPs(如HSP27)[41]。已经证实HSP27、HSP70、HSP90均参与了肿瘤的发生、发展。HSP40/DNAJ家族共有41种蛋白质,分布在细胞内不同的位置,可分为DNAJA、DNAJB、DNAJC亚族,大部分成员含有保守的J结构域,能够与抗凋亡分子伴侣HSP70结合,调节ATP酶活性,从而参与蛋白的翻译、折叠、异位等过程。目前对其研究仍较少,生理功能仍不明确,在既往研究中表明HSP40可通过泛素途经调节角蛋白数量[42];少数成员与肿瘤进展及转移相关,如DNAJB4(HLJ1)、DNAJA3(Tid1)、DNAJB6(MRJ)、DNAJC12(JDP1)、DNAJA1(HDJ2)[43]。本研究中DNAJB1在埃克替尼作用的肺腺癌细胞系PC-9中下调2.04倍。我们推测DNAJB1可能同其它热休克蛋白一样,参与了肿瘤的发生、发展。
应用生物信息学方法对NME1-NME2、STMN1、G3BP1三种蛋白进行分析,初步了解了它们的跨膜结构域、理化性质、亚细胞定位等生物学特征。NME1-NME2和stathmin亚型α均存在跨膜结构域,而G3BP1和stathmin亚型β无跨膜结构域,说明NME1-NME2、stathmin亚基α具有锚链于膜上的能力。结构域分析显示NME1-NME2具有两个核苷二磷酸激酶(NDK)序列,它催化二磷酸核苷向三磷酸核苷的转化,在信号传导等方面发挥重要作用;stathmin亚型α、β均含有一个stathmin样结构域,主要参与微管的解聚活性,可调控细胞周期;G3BP1具有核转运因子2结构域(nuclear transport factor 2, NTF2)和RNA识别基序的结构域(RRM),在真核生物基因表达调控中发挥重要作用。亚细胞定位分析提示NME1-NME2主要存在于细胞质中,推测可能参与了胞质内信号过程;STMN1主要存在于细胞质和细胞骨架中,与其调节微管系统的动力学功能相吻合;G3BP1分布广泛,主要存在于细胞质中,与Ras-GTPase激活蛋白结合活化Ras通路;STRING库预测所得与NME1-NME2相互作用蛋白得分高的主要有KCNN4、CCND1、PAPD4、ACLY、MYC、CNOT8、HSP72、NME1、EEF1A1,另外,NME1-NME2可能与STAT3通路有一定的关联;与STMN1蛋白相互作用密切的有AURKB、MAPK3、MAPK13、CDK1、TP53、MAPK1、CAMK2B、CAMK2G、NACA、ADCYAP1,可见其参与MAKP信号通路、细胞周期调控,并与TP53密切相关,相关研究表明p53可能是通过调节STMN1的表达,将细胞周期阻滞于G2/M期[44];与G3BP1蛋白相互作用密切的主要有CAPRIN1、USP10、EIF4G1、RASA1、CSK、HDAC6、OGFOD1、PRMT1、STYXL1、ALPP,是ras信号转导通路中的一个重要元素,参与肿瘤进展和转移过程[45]。
综上所述,本研究首次应用蛋白质组学方法研究了EGFR-TKI类药物的作用机制,目前尚未见相关报道。应用2-DE/MALDI-MS方法对埃克替尼作用前后的PC-9细胞进行了比较蛋白质组学研究,初步鉴定了16种蛋白质,这些蛋白质与埃克替尼作用机制密切相关,部分有可能成为新的分子治疗靶点。
参考文献
1钱桂生. 肺癌不同病理类型发病率的变化情况及原因[J/CD]. 中华肺部疾病杂志:电子版,2011,4(1):1-6.
2Torre A, Bray F, Siegel RL, et al. Global cancer statistics, 2012[J]. CA Cancer J Clin, 2015, 65(2): 87-108.
3Ciardiello F, Tortora G. A novel approach in the treatment of cancer:
targeting the epidermal growth factor receptor[J]. Clin Cancer Res, 2001, 7 (10): 2958-2970.
4Guan YS, He Q, Li M. Icotinib: activity and clinical application in Chinese patients with lung cancer[J]. Expert Opin Pharmacother, 2014, 15(5): 717-728.
5郑静娴, 张为民, 谢波, 等. 多西他赛与吉非替尼不同时序应用对人肺腺癌细胞的作用[J]. 临床肿瘤学杂志, 2012, 17(5): 385-391.
6Xuan X, An C, Zhou C. The mechanism of gefitinib resistance induced by hepatocyte growth factor in sensitive non-small cell lung cancer cells in vitro[J]. Zhongguo Fei Ai Za Zhi, 2013, 16(1): 1-6.
7Moll R, Franke WW, Schiller DL, et al. The catalog of human cytokeratins: patterns of expression in normal epithelia, tumors and cultured cells[J]. Cell, 1982, 31(1): 11-24.
8侯秀丽. 细胞角蛋白与肿瘤的关系[J]. 国际肿瘤学杂志,2009,36 (11):4.
9Fu BS, Liu W, Zhang JW, et al. [Serum proteomic analysis on metastasis-associated proteins of hepatocellular carcinoma][J]. Nan Fang Yi Ke Da Xue Xue Bao, 2009, 29 (9): 1775-1778.
10Warburg O. On respiratory impairment in cancer cells[J]. Science, 1956, 124(3215): 269-270.
11Greengard O, Herzfeld A. The undifferentiated enzymic composition of human fetal lung and pulmonary tumors[J]. Cancer Res, 1977, 37(3): 884-891.
12Possemato R, Marks KM, Shaul YD, et al. Functional genomics reveal that the serine synthesis pathway is essential in breast cancer[J]. Nature, 2011, 476(7360): 346-350.
13De Ingeniis J, Ratnikov B, Richardson AD, et al. Functional specialization
in proline biosynthesis of melanoma[J]. PLoS One, 2012, 7(9): e45190.
14Hepler JR, Gilman AG. G proteins[J]. Trends Biochem Sci, 1992,
17(10): 383-387.
15曾晓, 孙露霜, 杨立琳, 等. Ran调节因子RanBP1的生物学作用[J]. 东北农业大学学报,2010,41(3):150-153.
16Mistry SJ, Atweh GF. Role of stathmin in the regulation of the mitotic spindle: potential applications in cancer therapy[J]. Mt Sinai J Med, 2002, 69(5): 299-304.
17Curmi PA, Nogues C, Lachkar S, et al. Overexpression of stathmin in breast carcinomas points out to highly proliferative tumours[J]. Br J Cancer, 2000, 82(1): 142-150.
18Ocak S, Friedman DB, Chen H, et al. Discovery of new membrane-associated proteins overexpressed in small cell lung cancer[J]. J Thorac Oncol, 2014, 9(3): 324-336.
19Machado-Neto JA, de Melo Campos P, Favaro P, et al. Stathmin1 is involved in the highly proliferative phenotype of high-risk myelodysplastic syndromes and acute leukemia cells[J]. Leuk Res, 2014, 38(2): 251-257.
20Sabherwal Y, Mahajan N, Helseth DL, et al. PDEF downregulates stathmin expression in prostate cancer[J]. Int J Oncol, 2012, 40(6): 1889-1899.
21Brattsand G. Correlation of oncoprotein 18/stathmin expression in human breast cancer with established prognostic factors[J]. Br J Cancer, 2000, 83(3): 311-318.
22Seve P, Dumontet C. Chemoresistance in non-small cell lung cancer[J]. Curr Med Chem Anticancer Agents, 2005, 5(1): 73-88.
23Rana S, Maples PB, Senzer N, et al. Stathmin1: a novel therapeutic
target for anticancer activity[J]. Expert Rev Anticancer Ther, 2008, 8(9): 1461-1470.
24Li Y, Tong Y, Wong YH. Regulatory functions of Nm23-H2 in tumorigenesis: insights from biochemical to clinical perspectives[J]. Naunyn Schmiedebergs Arch Pharmacol, 2015, 388(2): 243-256.
25Desvignes T, Pontarotti P, Fauvel C, et al. Nme protein family evolutionary history, a vertebrate perspective[J]. BMC Evol Biol, 2009, 9: 256.
26Salerno M, Ouatas T, Palmieri D, et al. Inhibition of signal transduction
by the nm23 metastasis suppressor: possible mechanisms[J]. Clin Exp Metastasis, 2003, 20(1): 3-10.
27张浩, 邵荣光. G3bp:一个潜在的肿瘤治疗靶点[J]. 药学学报,2010,45(8):945-951.
28宁钧宇, 由江峰, 裴斐, 等. G3bp单克隆抗体制备及g3bp在肿瘤组织中表达的检测[J].中国病理学杂志,2005,34(4):5.
29French J, Stirling R, Walsh M, et al. The expression of Ras-GTPase activating protein SH3 domain-binding proteins, G3BPs, in human breast cancers[J]. Histochem J, 2002, 34(5): 223-231.
30Huang Y, Wernyj RP, Norton DD, et al. Modulation of specific protein expression levels by PTEN: identification of AKAP121, DHFR, G3BP, Rap1, and RCC1 as potential targets of PTEN[J]. Oncogene, 2005, 24(23): 3819-3829.
31Popovic D, Vucic D, Dikic I. Ubiquitination in disease pathogenesis
and treatment[J]. Nat Med, 2014, 20(11): 1242-1253.
32Margolis RL, Wilson L. Opposite end assembly and disassembly of microtubules at steady state in vitro[J]. Cell, 1978, 13(1): 1-8.
33Chen QY, Jiang ZY, Wu LJ, et al. [Expression of alpha-tubulin and MDR1 and their correlation with the biological features of non-small cell lung carcinoma][J]. Zhonghua Zhong Liu Za Zhi, 2010, 32(4): 278-282.
34Han EK, Gehrke L, Tahir SK, et al. Modulation of drug resistance by alpha-tubulin in paclitaxel-resistant human lung cancer cell lines[J]. Eur J Cancer, 2000, 36(12): 1565-1571.
35Winslow S, Leandersson K, Larsson C. Regulation of PMP22 mRNA
by G3BP1 affects cell proliferation in breast cancer cells[J]. Mol Cancer, 2014, 12(1): 156.
36Wang Y, Wen W, Yi Y, et al. Preventive effects of bexarotene and budesonide in a genetically engineered mouse model of small cell lung cancer[J]. Cancer Prev Res (Phila), 2009, 2(12): 1059-1064.
37Shah RK, Valdez TA, Wang Z, et al. Pulsed-dye laser and retinoic acid delay progression of oral squamous cell carcinoma: a murine model[J]. Laryngoscope, 2001, 111(7): 1203-1208.
38Moon RC, Kelloff GJ, Detrisac CJ, et al. Chemoprevention of OH-BBN-induced bladder cancer in mice by oltipraz, alone and in combination with 4-HPR and DFMO[J]. Anticancer Res, 1994, 14(1A): 5-11.
39Okuno M, Kojima S, Matsushima-Nishiwaki R, et al. Retinoids in cancer chemoprevention[J]. Curr Cancer Drug Targets, 2004, 4(3): 285-298.
40Edelman MJ, Smith R, Hausner P, et al. Phase Ⅱ trial of the novel retinoid, bexarotene, and gemcitabine plus carboplatin in advanced non-small-cell lung cancer[J]. J Clin Oncol, 2005, 23(24): 5774-5778.
41Kampinga HH. Chaperones in preventing protein denaturation in living
cells and protecting against cellular stress[J]. Handb Exp Pharmacol, 2006, (172): 1-42.
42Yamazaki S, Uchiumi A, Katagata Y. Hsp40 regulates the amount of keratin proteins via ubiquitin-proteasome pathway in cultured human cells[J]. Int J Mol Med, 2012, 29(2): 165-168.
43Mitra A, Shevde LA, Samant RS. Multi-faceted role of HSP40 in cancer[J]. Clin Exp Metastasis, 2009, 26(6): 559-567.
44Lu Y, Liu C, Cheng H, et al. Stathmin, interacting with NF-κ B, promotes tumor growth and predicts poor prognosis of pancreatic cancer[J]. Curr Mol Med, 2014, 14(3): 328-339.
45Giehl K. Oncogenic Ras in tumour progression and metastasis[J]. Biol Chem, 2005, 386 (3):193-205.
(本文编辑:黄红稷)
刘晓丽,李维,孟夏,等. 盐酸埃克替尼对人肺腺癌细胞系PC-9作用的比较蛋白质组学研究[J/CD]. 中华肺部疾病杂志: 电子版, 2015, 8(6): 674-683.
Comparative proteomics on mechanisms of lung adenocarcinoma cell line PC-9 induced by icotinib
LiuXiaoli,LiWei,MengXia,ZhangYuping,MingZhongjuan,Shihongyan,ShiJie,ZhongYnjie,WangWei,YangShanying.DepartmentofRespiratoryMedicine,theSecondAffiliatedHospital,MedicalSchoolofXi′anJiaotongUniversity,Xi′an710004,China
Correspondingauthor:YangShanying,Email:yangshuanying66@163.com
【Abstract】ObjectiveTo analyze icotinib-associated proteins of PC-9 cell with comparative proteomics and bioinformatics technique to discuss molecule mechanism of icotinib. MethodsAmoy Dx and real-time PCR were used to detect the EGFR mutation in PC-9 cells gene. MTT was used to investigate theproliferationinhibition oficotinibon PC-9 cells. Two dimensional gel electrophoresis (2-DE) was used to separate totally soluble proteins extracted from PC-9 cells either exposed to or not exposed to icotinib, and differentially expressed proteins were analyzed and identified by matrix-assisted laser desorption/ionization time of flight mass spectrometry (MALDI-TOF-MS). Proteins were identified by blasting NCBI or SWISS PORT database with Mascot software. ResultsThere was a deletion mutation located in exon 19.In the 10-4-10-9M concentration range,icotiib significantly concentration-dependent inhibited the growth of the PC-9 cells. 56 differentially expressed protein spots(Ratio≥1.5)were found. We finally selected 29 obviously differential protein spots (Ratio≥1.7) to analyze by MALDI-TOF-TOF-MS and got 16 proteins successfully. They can be classified into 5 categories based on their functions: enzymes related to cell metabolism, cytoskeleton proteins, signal transduction molecules, chaperone, proteins related to transcription and translation. Conclusions16 icotinib-related proteins are identified by comparative proteomics. These proteins may be associated with anti-tumor mechanism of icotinib and some of them may become potential drug targets.
【Key words】PC-9 cell;Icotinib;Lung cancer;Comparative proteomics;Bioinformatics
(收稿日期:2015-11-06)
中图法分类号:R563,R743
文献标识码:A
通讯作者:杨拴盈,Email: yangshuanying66@163.com
基金项目:国家自然科学基金资助(81172234)
作者单位: 710004 陕西,西安交通大学第二附属医院呼吸内科
DOI:10.3877/cma.j.issn.1674-6902.2015.06.002
猜你喜欢
中国骨质疏松杂志(2022年9期)2022-10-18
——水芹主要害虫识别与为害症状
长江蔬菜(2022年13期)2022-07-29
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
土壤与作物(2021年2期)2021-06-01
实用肿瘤学杂志(2020年4期)2020-12-08
家庭影院技术(2019年7期)2019-08-27
家庭影院技术(2018年11期)2019-01-21
中国癌症防治杂志(2019年5期)2019-01-04
医学综述(2011年12期)2011-12-09
