
淹水胁迫对杨桐幼苗生理生化性质的影响
2015-03-06 09:16吴江吴家胜
东北林业大学学报 2015年4期
吴江 吴家胜
(黄岩区林业特产局,黄岩,318020) (亚热带森林培育国家重点实验室培育基地(浙江农林大学))
责任编辑:潘 华。
我国拥有18 000 km的海岸线、众多的河流和湖泊,形成了数千公顷的湿地、季节性积水地[1]。淹水胁迫是植物一种主要的非生物胁迫,每年都给农业生产带来巨大的经济损失,而关于植物的耐水机理研究成为植物逆境研究的热点[2]。
杨桐(Cleyera japonica),是山茶科红淡比属的一种常绿灌木或小乔木[3]。杨桐是日本国民传统的供神祭祖的吉祥物,人们把杨桐枝条捆扎成束插放在庭堂上方供神,消耗量很大[4]。目前,对杨桐抗性方面的研究还未见报道,本文采用不同的水涝程度对杨桐幼苗进行水分胁迫,探讨杨桐抗涝机理和适应机制,为杨桐的采枝园的扩大栽培及其水分管理提供科学的理论依据。
1 材料与方法
试验材料为1年生杨桐幼苗。2月选取长势一致杨桐苗进行盆栽,每盆(3.78 L)1株,常规管理。选用双套盆法对试验材料进行淹水处理,共计4个水分梯度:土壤含水量为田间持水量75%作为对照处理(W4);轻度水渍,控制水面在土壤表面以下10 cm(W1);水渍胁迫,水面与土面持平(W2);水涝胁迫,水面高出土面4 cm(W3)。每个处理3个重复,每个重复6盆,于处理第5天取样,选取2~3轮的功能叶片,剪碎混均匀后作为混合样。
SOD采用氮蓝四唑光还原比色法测定,POD采用愈创木酚法测定[5]。可溶性糖采用蒽酮法测定,可溶性蛋白质采用考马斯亮蓝法测定,细胞膜透性采用电导仪法测定[6]。叶片相对含水量采用烘干法测定[7]。
主成分分析采用DPS7.05软件系统进行,方差分析采用Duncan法进行多重差异性比较。
2 结果与分析
2.1 水涝胁迫对杨桐叶片保护酶活性和质膜透性的影响
从表1可以发现,杨桐幼苗叶片SOD活性、POD活性、质膜透性随着胁迫程度的加剧显现上升趋势,淹水胁迫对SOD、POD的影响达到显著水平,W3处理下比对照显著上升。质膜透性W3处理下SOD活性、POD活性、质膜透性比对照分布上升了75.0%、30.0%和25.1%。
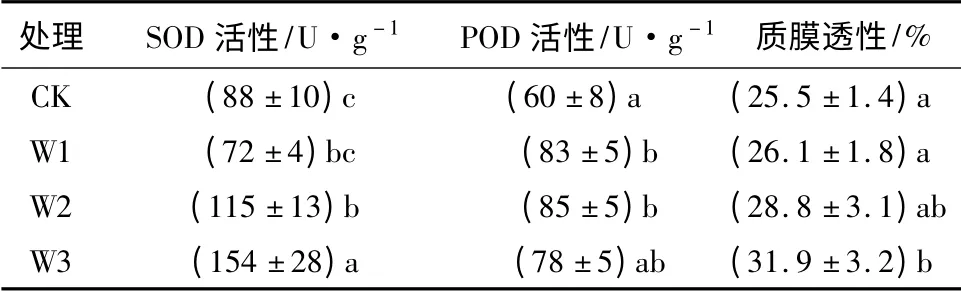
表1 水涝胁迫对杨桐叶片SOD、POD、质膜透性的影响
2.2 淹水胁迫对杨桐幼苗叶片叶绿素质量分数的影响
从表2可以发现,杨桐幼苗叶片叶绿素质量分数随着胁迫程度的加剧显现下降趋势,淹水胁迫对叶绿素a和叶绿素b质量分数的影响达到显著水平,淹水胁迫下w(叶绿素a)∶w(叶绿素b)有也有不同程度的下降,但未达到显著性差异。W3处理下叶绿素a和叶绿素b分别比对照下降39.8%和29.3%。
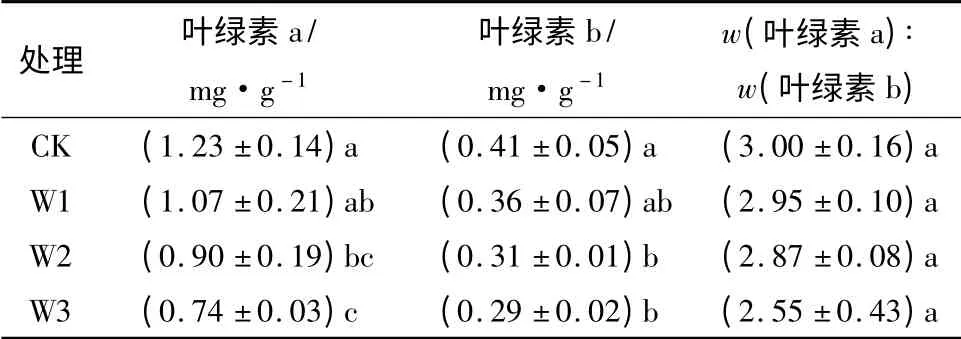
表2 不同处理对杨桐幼苗叶片叶绿素质量分数的影响
2.3 淹水胁迫对杨桐幼苗叶片渗透调节和相对含水量的影响
从表3可以发现,杨桐幼苗叶片可溶性蛋白和可溶性糖质量分数随着胁迫程度的加剧而有所增加,各个处理和对照相比均显著增加,淹水胁迫下叶片相对含水量有不同程度的下降,但未达到显著性差异。W3处理下可溶性糖、可溶性蛋白比对照上升了140.3%和101.6%。

表3 不同处理对杨桐幼苗叶片可溶性糖、可溶性蛋白和相对含水量的影响
2.4 主成分分析
对杨桐幼苗叶片的9个指标进行主成分分析,统计表明:前3个主成分已经基本携带了9个抗涝指标的大部分信息,前3个成分共累积贡献率为88.58%(第1主成分64.20%,第2主成分13.60%,第3主成分10.78%)。第1主成分中,各项系数差异不大,能基本反映各指标的综合情况。第2主成分与w(叶绿素a)∶w(叶绿素b)、POD密切相关,第3主成分与POD密切相关。
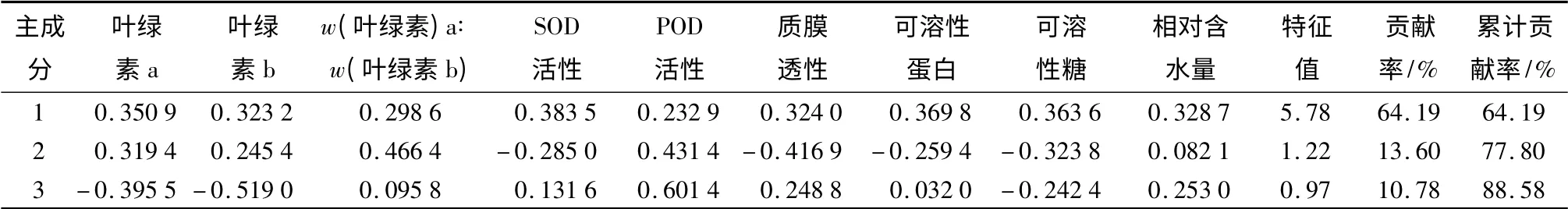
表4 杨桐抗涝性状指标主成分特征向量和贡献率
3 结论与讨论
植物在淹水胁迫下,活性氧的积累会对细胞造成伤害,抗氧化防御系统的存在,是细胞免于伤害的主要原因之一[8-9]。SOD是超氧阴离子自由基的主要清除剂,能将其转化成H2O2和O2,POD能催化H2O2生成氧气[10]。细胞膜透性能反映膜的稳定性,较高的保护酶活性可以维持细胞的形态和功能[11]。本试验研究发现,杨桐幼苗叶片SOD活性、POD活性、质膜透性随着胁迫程度的加剧显现上升趋势,淹水胁迫对SOD、POD的影响达到显著水平,W3处理下比对照显著上升,这与前人对芝麻[12]、喜树[13]等研究一致。
可溶性糖、可溶性蛋白等是植物重要的渗透调节物质,渗透调节是植物对水淹胁迫的一种适应和保护机制[14-15],同时可溶性蛋白是植物体代谢过程中蛋白损伤的重要指标[16]。植物体内累积大量渗透调节物质降低植物体内渗透势,以利植物在水涝逆境下维持植物体正常生长所需水分[17]。本试验中,杨桐幼苗叶片可溶性蛋白和可溶性糖质量分数随着胁迫程度的加剧而有所增加,各个处理和对照相比均显著增加,淹水胁迫下叶片相对含水量有不同程度的下降,但未达到显著性差异。这与徐锡增[18]、曾德静[19]的研究一致,说明杨桐通过可溶性糖和可溶性蛋白来调节细胞渗透势。
叶绿素质量分数是植物光合性能、衰老状况的直观表现,其质量分数的高低是一个有效的涝害鉴定指标[20]。本文研究发现,杨桐幼苗叶片叶绿素质量分数随着胁迫程度的加剧显现下降趋势,淹水胁迫对叶绿素a和叶绿素b质量分数的影响达到显著水平,淹水胁迫下w(叶绿素a)∶w(叶绿素b)也有不同程度的下降,但未达到显著性差异。
植物抗涝机理较为复杂,植物通过多条途径抵御水涝对植物的伤害,不能采用单一指标对其进行抗涝评价。本试验对9个与抗涝相关指标进行主成分分析,发现3个主成分贡献率为88.58%,为杨桐幼苗抗涝删选提供理论指导。
[1]曹福亮,方升佐.水分(淹水)胁迫与林木适应性[M].北京:中国林业出版社,2003.
[2]王良桂,杨秀莲.淹水对2个桂花品种生理特性的影响[J].安徽农业大学学报,2009,36(3):382-386.
[3]钱玉红,孙丽华.浙江杨桐考察[J].浙江林业科技,1994,14(1):42-46.
[4]吴家胜,应叶青,程晓建,等.杨桐不同立体经营模式比较研究[J].浙江林学院学报.2007,24(5):555-558.
[5]李合生.植物生理生化试验原理和技术[M].北京:高等教育出版社,2000:134-200.
[6]邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000:258-260.
[7]汤章城.现代植物生理学试验指南[M].北京:科学出版社,1999:302-306.
[8]王晶英,赵雨森,王臻,等.干旱胁迫对银中杨生理生化特性的影响[J].水土保持学报,2006,20(1):197-200.
[9]余玲,王彦荣,Garnet T T,等.紫花苜蓿不同品种对干旱胁迫的生理响应[J].草业学报,2006,6(3):75-85.
[10]谭淑端,朱明勇,党海山,等.三峡库区狗牙根对深淹胁迫的生理响应[J].生态学报,2009,29(7):3685-3691.
[11]任鹏,赵宝平,刘瑞芳,等.水分胁迫对燕麦穗颖渗透调节和抗氧化能力的影响[J].西北植物学报,2014,34(10):2049-2055.
[12]刘华山,韩锦峰,孟凡庭,等.土壤渍水下芝麻叶片中几个与抗性能力有关的生理指标的变化[J].植物生理学通讯,2001,37(2):106-108.
[13]蔡金峰,曹福亮,汪贵斌.淹水胁迫对喜树幼苗LDH及保护酶的影响[J].福建林学院学报,2008,28(1):65-68.
[14]张晓平,方炎明,陈永江.淹涝胁迫对鹅掌楸属植物叶片部分生理指标的影响[J].植物资源与环境学报,2006,15(1):41-44.
[15]廖文燕,高捍东.金钱松幼苗在淹水胁迫过程中的生理响应[J].林业科技开发,2011,25(3):27-31.
[16]曹永慧,周本智,陈双林.弱光下水分胁迫对不同产地披针叶茴香幼苗生理特性的影响[J].生态学报,2014,34(4):814-822.
[17]王瑞,梁坤伦,周志宇,等.不同淹水梯度对紫穗槐营养生长和生理响应[J].草业学报,2012,21(1):149-155.
[18]徐锡增,唐罗忠,程淑婉.涝渍胁迫下杨树内源激素及其它生理反应[J].南京林业大学学报,1999,23(1):1-5.
[19]曾德静,王铖,刘军,等.水涝胁迫下海州常山形态和生理指标的变化[J].浙江农林大学学报,2013,30(2):172-178.
[20]李雪,金研铭,郭太君,等.连续水淹胁迫对雪柳叶片部分生理指标的影响[J].北方园艺,2013(12):54-57.
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
世界科学(2020年1期)2020-02-11
中国生物医学工程学报(2019年5期)2019-07-16
中国果业信息(2019年1期)2019-01-05
中成药(2018年8期)2018-08-29
生物学教学(2017年9期)2017-08-20
Coco薇(2017年5期)2017-06-05
现代电生理学杂志(2015年4期)2015-07-18
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
