
RNA干扰沉默Fra-1基因对膀胱癌T24细胞侵袭和迁移的影响
2015-03-04 05:31谢海峰张宏波乔庆东马小芳孟翔宇河南省新乡市中心医院泌尿外一科河南新乡45000郑州大学基础医学院解剖学教研室河南郑州45000郑州大学基础医学院基础肿瘤学教研室河南郑州45000
中国现代医学杂志 2015年23期
谢海峰,张宏波,乔庆东,马小芳,孟翔宇(.河南省新乡市中心医院泌尿外一科,河南新乡45000;.郑州大学基础医学院解剖学教研室,河南郑州45000;.郑州大学基础医学院基础肿瘤学教研室,河南郑州45000)
RNA干扰沉默Fra-1基因对膀胱癌T24细胞侵袭和迁移的影响
谢海峰1,张宏波2,乔庆东1,马小芳3,孟翔宇2
(1.河南省新乡市中心医院泌尿外一科,河南新乡453000;2.郑州大学基础医学院解剖学教研室,河南郑州450001;3.郑州大学基础医学院基础肿瘤学教研室,河南郑州450001)
摘要:目的研究RNA干扰沉默Fra-1基因表达对人膀胱癌T24细胞侵袭及迁移能力的影响。方法设计合成Fra-1特异性siRNA并转染人膀胱癌T24细胞。实验设空白对照组(转染时只加脂质体)、阴性对照组(无关序列siRNA转染)和干扰组(Fra-1特异siRNA转染)。采用RT-PCR检测Fra-1、MMP-2及MMP-9 mRNA水平。Western blot检测Fra-1、MMP-2、MMP-9、STAT3及磷酸化STAT3(p-STAT3)蛋白表达水平,Transwell小室法和划痕实验检测细胞侵袭和迁移能力。结果干扰组T24细胞Fra-1、MMP-2及MMP-9 mRNA水平较其他两组明显降低(P<0.01),并且其MMP-2、MMP-9及p-STAT3蛋白表达受到明显抑制(P< 0.01)。干扰组T24细胞侵袭力和迁移力较其他两组明显下降(P<0.01)。结论RNA干扰可有效抑制T24细胞中Fra-1基因的表达,降低细胞侵袭及迁移能力,其机制可能与STAT3/MMPs信号通路有关。
关键词:膀胱肿瘤;RNA干扰;转染;Fra-1;STAT3;MMPs
Fra-1基因定位于染色体11q13,编码长度为1.7 kb的成熟mRNA,其表达产物Fra-1蛋白由271个氨基酸组成,作为核转录因子AP-1家族中Fos亚家族的成员之一,其可通过亮氨酸拉链结构域与Jun蛋白形成异源二聚体,调控靶基因转录,介导细胞的生长、发育、分化及凋亡[1]。Fra-1在多种肿瘤组织中高表达,参与肿瘤发生、发展及侵袭转移[1-3]。本研究中,笔者采用RNA干扰技术抑制膀胱移行细胞癌T24细胞中Fra-1基因的表达,观察对T24细胞侵袭及迁移能力的影响并探讨其分子机制,为以Fra-1为靶点的膀胱癌基因治疗提供理论和实验依据。
1 材料与方法
1.1细胞和试剂
人膀胱移行细胞癌T24细胞株购自中国科学院上海生命科学研究院细胞资源中心,RT-PCR试剂盒为大连TaKaRa生物公司产品,兔抗人Fra-1、MMP-2、MMP-9及β-actin多克隆抗体购自Santa Cruz公司,鼠抗人STAT3,p-STAT3及Matrigel基质胶购自BD公司,HRP标记的羊抗兔及羊抗鼠二抗购自北京中杉金桥公司,Transwell小室购自Costar公司;Lipofectin2000购自Invitrogen公司。
1.2Fra-1 siRNA设计及制备
根据人Fra-1基因(NM_005438.3)设计序列:正向引物5'-GUUCCGAGCGGACGGGUCCUU-3',反向引物3'-GGACCCGUCCGCUCGGAACUU-5'。siRNA序列均由广州锐博公司合成并纯化。为了避免可能偏靶,采用严格BALST进行比对,GenBank中证实Fra-1 siRNA仅与Fra-1基因存在相应的匹配位点,无关序列dsRNA与任何已知哺乳动物基因无匹配。
1.3细胞培养及siRNA转染
采用无抗生素、含体积分数10%FBS的RPMI 1640培养液,在37℃、体积分数为5%二氧化碳CO2条件下培养T24细胞。调整细胞浓度,接种至6孔板,每孔约1×106个细胞。实验分成3组,空白对照组(转染时只加脂质体)、阴性对照组(无关序列siRNA转染)和干扰组(Fra-1特异siRNA转染)。待细胞生长至70%~80%时,采用脂质体Lipofe-ctamine 2000转染,转染步骤严格按照说明书进行。
1.4转染效率观察
细胞转染48 h后,使用480 nm波长蓝光激发,荧光显微镜下观察细胞,判断转染效率。
1.5RT-PCR检测Fra-1、MMP-2及MMP-9 mRNA水平
细胞转染48 h后收集细胞,PBS洗涤2、3次,按Trizol说明书操作提取细胞总RNA,取2μg总RNA用MMLV逆转录酶将其逆转录为cDNA。以2μl的cDNA为模板进行PCR扩增,β-actin为内参照。引物由上海英骏生物技术有限公司合成(见附表)。PCR扩增条件:94℃预变性5 min,然后94℃变性30 s,57℃退火30 s,72℃延伸10 s,总共30个循环,最后72℃延长5 min。PCR产物经2%琼脂糖凝胶电泳。紫外线透射观察仪观察并照相。

附表RT-PCR引物序列
1.6Western blot检测Fra-1、MMP-2,MMP-9、STAT3及p-STAT3蛋白表达
转染48 h后收集细胞,使用预冷的裂解液对细胞进行裂解,按说明书操作提取蛋白,Bradford法进行蛋白质定量。SDS-PAGE(8%分离胶,5%浓缩胶)电泳后,蛋白质湿转至PVDF膜上。5%脱脂奶粉封闭,分别加入稀释的一抗37℃孵育2 h,TBST洗膜后,加入相应的辣根酶标记的二抗37℃孵育1 h,TBST洗膜。化学发光增强液与膜充分接触,暗室曝光显影,冲洗胶片,凝胶成像系统扫描成像。
1.7Transwell侵袭实验
用50 mg/L Matrigel(1∶8)稀释液包被Transwell小室底部膜的上室面,4℃风干,37℃水化30 min,取转染48 h细胞,用100μl含0.1%BSA的无血清培养基调整细胞密度至1×106个细胞/ml并将悬液加入Transwell小室,下室加入600μl完全培养基作为趋化剂,培养24 h。取出上室,用棉签擦去上室中未侵袭细胞,PBS浸洗1遍。10%甲醇固定30 s,PBS连续洗3遍。用0.2%结晶紫染色10 min,用棉签擦去多余结晶紫,PBS再洗3遍。光镜下观察滤膜下表面的细胞,随机选取5个视野的细胞计数,取平均值。
1.8划痕实验
收集对数生长期细胞,以1×106个/孔密度接种于6孔培养板。24 h后进行细胞转染,步骤同1.3。转染6 h后,用无菌Tip头(规格200μl)在单层细胞上做“1”字形划痕,宽度和长度在1 mm×20 mm左右,每组设3个复孔。用无血清培养基轻轻吹打划痕附近,吸去培养基,反复3次,尽量将刮掉的细胞冲洗干净。加入不含双抗无FBS的RPMI 1640培养液继续培养,于24和48 h观察并拍照,显微镜下计数单位面积划痕区内迁移的细胞数。
1.9统计学方法
采用SPSS17.0软件进行数据分析,实验数据用均数±标准差(±s)表示,组间比较用单因素方差分析,以P<0.05为差异有统计学意义。
2 结果
2.1细胞转染效率
T24细胞转染48h后,荧光显微镜下观察,阴性对照组和干扰组90%以上细胞可见绿色荧光(见图1)。
2.2siRNA对T24细胞Fra-1、MMP-2及MMP-9 mRNA水平的影响
由图2显示,Fra-1 siRNA转染可有效抑制T24细胞Fra-1、MMP-2及MMP-9 mRNA的表达,其抑制率分别为51.5%、47.3%和35.6%(P<0.01)。阴性对照组与空白对照组比较,差异无统计学意义(P> 0.05)。
2.3siRNA对T24细胞Fra-1、MMP-2、MMP-9、STAT3及p-STAT3蛋白表达的影响
由图3显示,与空白对照组比较,干扰组细胞Fra-1、MMP-2、MMP-9及p-STAT3蛋白表达明显下调(P<0.01),其下调量分别为65.4%、53.7%、47.0%和81.0%,而总STAT3蛋白表达无明显变化(P>0.05)。空白对照组与阴性对照组比较差异无统计学意义(P>0.05)。
2.4siRNA对T24细胞体外侵袭和迁移能力的影响
Transwell侵袭实验结果见图4,空白对照组、阴性对照组及干扰组中穿透Matrigel膜的细胞数分别为(505.7±51.6)、(496.5±49.7)和(307.5±47.1)。干扰组细胞侵袭能力较空白对照组和阴性对照组分别减少39.2%(P<0.01)和38.1%(P<0.01),而空白对照组与阴性对照组间比较,差异无统计学意义(P>0.05)。
划痕实验结果见图5,划痕处理24 h后,空白对照组、阴性对照组及干扰组中迁移细胞数分别为(32.7±0.3)、(31.5±0.5)和18.3±0.6)。划痕处理48 h后,以上3组中迁移细胞数分别为(53.6±1.8),(51.3±2.1)和(29.5±1.5)。在两个时间点,干扰组分别与其他两组比较,差异有统计学意义(P<0.01),而空白对照组与阴性对照组比较,差异无统计学意义(P>0.05)。
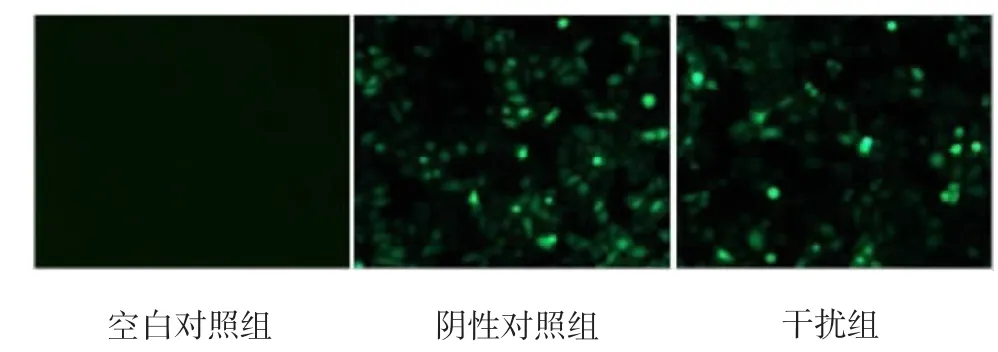
图1 T24细胞转染48 h后荧光显微镜观察结果
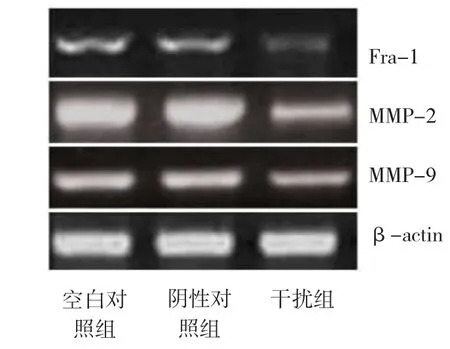
图2 RT-PCR检测T24细胞Fra-1、MMP-2 和MMP-9 mRNA水平
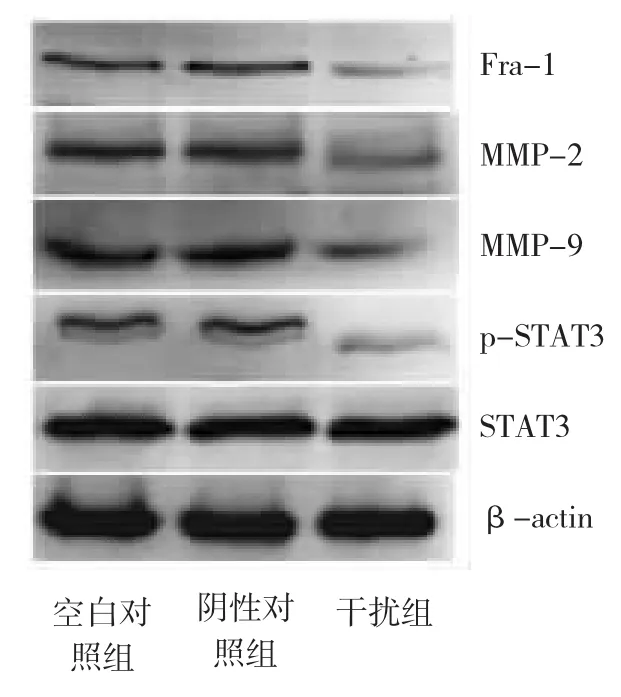
图3 Western blot检测T24细胞Fra-1、MMP-2、MMP-9、STAT3及p-STAT3蛋白表达
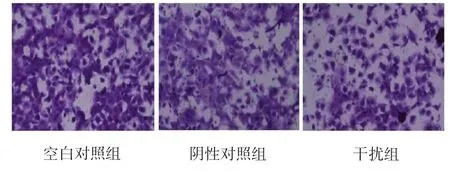
图4 Transwell小室法检测T24细胞的侵袭能力

图5 划痕实验检测T24细胞的迁移能力
3 讨论
膀胱癌是我国最常见的泌尿系肿瘤,占全部恶性肿瘤的3.2%,具有恶性程度高、浸润发展、易转移及复发率高等特点,其中侵袭和转移是临床上导致膀胱癌治疗失败的主要原因。本实验以T24细胞为研究对象,该细胞株来源于转移性人膀胱移行细胞癌,适合膀胱癌的侵袭转移研究。
Fra-1在肿瘤组织的高表达与肿瘤的侵袭能力密切相关。DESMET等[3]发现RNA干扰Fra-1后乳腺癌细胞的体内侵袭能力显著降低,认为Fra-1与肿瘤转移及复发密切相关,可作为乳腺癌预后的指标。SAYAN等[4]亦发现80%侵袭性膀胱癌中Fra-1呈高表达,Fra-1可通过直接上调受体酪氨酸激酶AXL影响膀胱癌细胞的侵袭能力。本实验中,笔者亦证实膀胱癌T24细胞表达Fra-1,siRNA转染后Fra-1 mRNA水平及蛋白表达量均明显下调,并且细胞的迁移及侵袭能力显著减弱。
MMP-2和MMP-9是基质金属蛋白酶类(matrix metallopmteinase,MMP)家族中的重要成员,能够降解Ⅳ型、Ⅴ、Ⅵ及Ⅹ型胶原等多种细胞外基质成分[5]。临床研究表明[6],膀胱癌的恶性程度、侵袭转移能力及预后与MMP-2及MMP-9异常高表达密切相关。ADISESHAIAH等[7]发现,Fra-1可通过诱导MMP-1、MMP-9活性及EGFR信号途径活化增强肺癌上皮细胞的侵袭能力。KIMURA等[8]亦发现骨肉瘤143B细胞中MMP-1的基因表达依赖于Fra-1激活。SCHR魻ER等[9]发现宫颈癌细胞中,STAT3可通过间接诱导单核细胞趋化蛋白-1而上调MMP-9的表达。本实验中,通过RNA干扰下调T24细胞Fra-1基因表达后发现,细胞MMP-2及MMP-9 mRNA水平和蛋白表达量均明显下调,说明RNA干扰下调Fra-1表达可影响侵袭相关基因MMP-2及MMP-9表达,从而抑制膀胱癌细胞的侵袭能力。
细胞因子或生长因子与相应的受体结合后可募集JAKs催化受体的酪氨酸残基磷酸化并活化STAT3,使之转运到细胞核内与特定DNA序列结合,调控基因转录[10]。已有研究表明[11-12],STAT3信号通路可在增殖、分化、侵袭、转移、血管形成及抗凋亡等多方面参与肿瘤的发生、发展。ZUGOWSKI等[13]发现AP-1可激活Stat3,上调其下游基因MMP-1表达,增强结肠癌细胞的侵袭能力。此外,LUO等[14]还发现Fra-1可活化乳腺癌细胞Stat3信号途径,上调MMP9、VEGF及TGF-β表达,从而增强肿瘤细胞的侵袭和转移能力。本实验发现干扰组中p-STAT3蛋白表达下调81%,说明RNA干扰可明显抑制T24细胞中STAT3磷酸化,从而阻断STAT3活化,有效地调控其下游的侵袭相关基因MMP-2及MMP-9的表达。
本实验研究初步证实RNA干扰沉默Fra-1基因可有效抑制T24细胞中Fra-1基因表达,降低T24细胞的侵袭及迁移能力,其分子机制可能与STAT3/ MMPs信号通路有关,提示Fra-1可作为膀胱肿瘤基因治疗的理想靶点。
参考文献:
[1] YOUNG MR, COLBURN NH. Fra-1 a target for cancer prevention or intervention[J]. Gene, 2006, 379: 1-11.
[2] USUI A, HOSHINO I, AKUTSU Y, et al. The molecular role of Fra-1 and its prognostic significance in human esophageal squamous cell carcinoma[J]. Cancer, 2012, 118(13): 3387-3396.
[3] DESMET CJ, GALLENNE T, PRIEUR A, et al. Identification of a pharmacologically tractable Fra-1/ADORA2B axis promoting breast cancer metastasis [J]. Proc Natl Acad Sci USA, 2013, 110 (13): 5139-5144.
[4] SAYAN AE, STANFORD R, VICKERY R, et al. Fra-1 controls motility of bladder cancer cells via transcriptional upregulation of the receptor tyrosine kinase AXL [J]. Oncogene, 2012, 31 (12): 1493-1503.
[5]SHUMANMOSSLA,JENSEN-TAUBMANS, STETLER-STEVE-NSON WG. Matrix metalloproteinases: changing roles in tumor progression and metastasis[J]. Am J Pathol, 2012, 181(6): 1895-1899.
[6] OFFERSEN BV, KNAP MM, HORSMAN MR, et al. Matrix metalloproteinase-9 measured in urine from bladder cancer patients is an independent prognostic marker of poor survival [J]. Acta Oncol, 2010, 49(8): 1283-1287.
[7] ADISESHAIAH P, VAZ M, MACHIREDDY N, et al. A Fra-1-dependent, matrix metalloproteinase driven EGFR activation promotes human lung epithelial cell motility and invasion[J]. J Cell Physiol, 2008, 216(2): 405-412.
[8] KIMURA R, ISHIKAWA C, ROKKAKU T, et al. Phosphorylated c-Jun and Fra-1 induce matrix metalloproteinase-1 and thereby regulate invasion activity of 143B osteosarcoma cells[J]. Biochim Biophys Acta, 2011, 1813(8): 1543-1553.
[9] SCHR魻ER N, PAHNE J, WALCH B, et al. Molecular pathobiology of human cervical high-grade lesions: paracrine STAT3 activation in tumor-instructed myeloid cells drives local MMP-9 expression[J]. Cancer Res, 2011, 71(1): 87-97.
[10] JOHNSTON PA, GRANDIS JR. STAT3 signaling: anticancer strategies and challenges[J]. Mol Interv, 2011, 11(1): 18-26.
[11] WANG XH, LIU BR, QU B, et al. Silencing STAT3 may inhibit cell growth through regulating signaling pathway, telomerase, cell cycle, apoptosis and angiogenesis in hepatocellular carcinoma: potential uses for gene therapy[J]. Neoplasma, 2011, 58(2): 158-171.
[12] ERNST M, PUTOCZKI TL. Stat3: linking inflammation to (gastrointestinal) tumourigenesis [J]. Clin Exp Pharmacol Physiol, 2012, 39(8): 711-718.
[13] ZUGOWSKI C, LIEDER F, M譈LLER A, et al. STAT3 controls matrix metalloproteinase-1 expression in colon carcinoma cells by both direct and AP-1-mediated interaction with the MMP-1 promoter[J]. Biol Chem, 2011, 392 (5): 449-459.
[14] LUO YP, ZHOU H, KRUEGER J, et al. The role of proto-oncogene Fra-1 in remodeling the tumor microenvironment in support of breast tumor cell invasion and progression[J]. Oncogene, 2010, 29(5): 662-673.
(申海菊编辑)
Effects of Fra-1 gene silencing by small interfering RNA on invasion and metastasis of bladder cancer T24 cells
Hai-feng XIE1, Hong-bo ZHANG2, Qing-dong QIAO1, Xiao-fang MA3, Xiang-yu MENG2(1. Department of Urology, Xinxiang Central Hospital, Xinxiang, Henan 453000, P.R. China; 2. Department of Anatomy, 3. Department of Basic Oncology, School of Basic
Medicine, Zhengzhou University, Zhengzhou, Henan 450001, P.R. China)
Abstract:【Objective】To investigate the effects of Fra-1 gene silencing by small interfering RNA (siRNA) on invasion and migration of bladder cancer T24 cells in vitro.【Methods】The Fra-1 siRNA was constructed and then transfected into T24 cells in vitro. The blank control group, negative control group and siRNA transfection group were designed in this study. The expression level of Fra-1, MMP-2 and MMP-9 mRNAs were detected by RT-PCR. The expressions of Fra-1, MMP-2, MMP-9, total STAT3 and p-STAT3 proteins were detected by Western blot. The invasion and migration capabilities were evaluated by using Transwell chamber migration assay and scratch wound healing assay, respectively.【Results】The levels of Fra-1 mRNA, MMP-2 and MMP-9 mRNAs in the siRNA transfection group were significantly decreased compared with that in other groups (P< 0.01), so were the expressions of MMP-2, MMP-9 and p-STAT3 proteins (P< 0.01). In the siRNA transfection group, the invasion and migration capabilities of T24 cells were also significantly decreased compared with those in other groups (P< 0.01).【Conclusions】Small interfering RNA can efficiently inhibit the expression of Fra-1, which results in decreased capacities of invasion and migration of T24 cells. The mechanism may be involved in STAT3/MMPs signaling pathway.
Key words:bladder tumor; small interfering RNA; transfection; Fra-1; STAT3; matrix metalloproteinase
[通信作者]张宏波,E-mail:zhanghb@zzu.edu.cn
收稿日期:2014-11-28
文章编号:1005-8982(2015)23-0023-05
中图分类号:R737.14
文献标识码:A
猜你喜欢
新农业(2022年21期)2022-11-18
中日友好医院学报(2022年4期)2022-10-15
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年1期)2022-02-28
皮肤病与性病(2021年3期)2021-07-30
实用肿瘤学杂志(2020年6期)2020-12-09
看世界·学术下半月(2020年7期)2020-09-10
东北大学学报(自然科学版)(2020年5期)2020-05-22
Drug Combination Therapy(2020年1期)2020-02-14
满族文学(2018年6期)2018-12-27
