
广西特有属
——异裂菊属开花生物学及繁育系统特征
2015-02-28 02:12史艳财唐健民陈宗游柴胜丰
西北植物学报 2015年4期
史艳财,邹 蓉,唐健民,陈宗游,柴胜丰,韦 霄
(广西壮族自治区中国科学院广西植物研究所,广西桂林 541006)
广西特有属
——异裂菊属开花生物学及繁育系统特征
史艳财,邹 蓉,唐健民,陈宗游,柴胜丰,韦 霄*
(广西壮族自治区中国科学院广西植物研究所,广西桂林 541006)
以异裂菊属5个种的实生苗为试验材料,通过田间观察,采用TTC法、联苯胺-过氧化氢法、杂交指数、花粉-胚珠比等方法,对异裂菊属的开花物候、花部特征、繁育系统及访花昆虫行为进行了研究。结果表明:(1)绢叶异裂菊的花期最早,为6~7月份;小花异裂菊和柳州异裂菊花期为6~8月份;凹脉异裂菊和异裂菊的花期为8~12月份,整个花期持续约5个多月,单花花期约为3 d,单个花序花期约为5 d。(2)异裂菊属的花粉在开花后1~18 h具有活力,柱头的可授性持续约48 h;杂交指数OCI为4,花粉/胚珠比(P/O)为1 450~2 250,繁育系统属于异交、部分自交亲和,需要传粉者。(3)传粉昆虫主要是食蚜蝇科灰带管食蚜蝇和胡蜂科黑尾胡蜂,昆虫访花高峰期在上午10:00~12:00,每次可连续访问多朵花,每朵花访问时间约为2~10 s。
异裂菊属;花部特征;杂交指数;花粉/胚珠比;繁育系统
在中国近30 000种高等植物中,至少有3 000多种处于受威胁或濒临灭绝的境地,如何保护濒危植物已成为当前植物学界急需解决的重大课题。繁育系统对植物的进化路线和表征变异有较大影响,是种群有性生殖的纽带,已成为当今进化生物学研究中最为活跃的领域之一[1]。繁育系统主要包括花部综合特征、花各性器官的寿命、花开放式样、自交亲和程度和交配系统[2],其中交配系统是核心[1]。开花生物学特性、植株的开花数目和排列方式会影响繁育系统,特别是植株上的开花数目会增加对传粉者的吸引。在众多的致濒因素中,繁育系统的生殖力、存活力、适应力低下往往是植物濒危的根本原因[1,3]。小花木兰因花期及花粉可授期短、昆虫访花频率低、同花期植物的竞争和外界不良环境等原因造成传粉率低下,成为其濒临灭绝的重要原因[4]。而传粉强度和传粉效率是影响刺五加[5]、南川升麻[6]、鹅掌楸[7]等濒危植物结实、导致其濒危的重要限制因子。因此,关于濒危植物繁育生物学特性的研究对掌握其演化维持机制具有重要意义。
异裂菊属(HeteroplexisChang),菊科,产自中国广西,为广西稀有的特有属,1937年作为一个单种属发表,现该属共有5个种:异裂菊(HeteroplexisvernonioidesChang)、小花异裂菊(HeteroplexismicrocephalaY.L.Chen)、绢叶异裂菊(HeteroplexissericophyllaY.L.Chen)、凹脉异裂菊(HeteroplexisimpressinerviaJ.Y.Liang)和柳州异裂菊(HeteroplexisincanaJ.Y.Liang)[8]。异裂菊属分布区域狭窄,例如,小花异裂菊仅分布于广西阳朔、融安等县的石灰岩石山灌丛或者向阳处[9],种群个体数量稀少且正在逐年减少。异裂菊、小花异裂菊和绢叶异裂菊已被列为中国重点保护植物[10]。
目前关于异裂菊属的研究较少,仅有关于小花异裂菊核型[11]、萜类成分及其活性[12]、迁地保护适应性[13]等少数几个方面。从繁殖生态学入手,掌握稀有植物花部特征和传粉适应性等基本的繁育特征是有效开展其保育工作的关键和首要步骤。目前涉及异裂菊属植物繁育系统、开花生物学特性方面的研究还未见报道。本研究的主要目的在于:(1)探明异裂菊属花部综合特征表现;(2)分析异裂菊属繁育系统。通过上述研究,综合分析异裂菊属在石灰岩石山环境下的开花和繁育特性,为种群保育提供生殖生物学方面的科学指导。
1 材料和方法
1.1 实验材料
试验在广西壮族自治区桂林市广西植物研究所种质资源圃进行。试验地气候特点为:中亚热带季风气候;四季分明且雨热基本同季;年平均气温为18.9 ℃;7月最热,月平均气温为28 ℃;1月最冷,月平均气温7.8 ℃;年平均无霜期309 d;年平均降雨量1 949.5 mm;年平均蒸发量1 490~1 905 mm;年平均相对湿度为73%~79%;年平均日照时数为1 670 h;土壤为酸性红壤。
以从广西龙州县、阳朔县、宁明县等地引种并在广西植物研究所栽培3a的异裂菊属的小花异裂菊、凹脉异裂菊、柳州异裂菊、绢叶异裂菊和异裂菊5个种的实生苗为试验材料,每个种约有30株。
1.2 方 法
1.2.1 花部特征和开花动态观察 采集盛开花30朵,统计其花瓣数、花冠直径、花瓣长度和宽度、花丝长度、柱头高度、花萼长和宽以及子房长和宽等指标,并计算其平均值。观察和记录单花的开花特性。随机选取5~10株未开放的花序进行标记,观察花序的开放顺序和开花天数[14]。
1.2.2 花粉活力和柱头可授性检测 于盛花期对异裂菊属5个种的花朵进行标记,按预定时间取花粉,每次取5朵,使用TTC(0.5%)法检测花粉活性。以染色率表示花粉的活力:
花粉活力=红色花粉数/观察花粉总数×100%
柱头的可授性检验采用过氧化氢-联苯胺法(1%联苯胺∶3%过氧化氢∶水=4∶11∶22,体积比)。若柱头具有可授性,则周围出现蓝色并有大量气泡产生[15]。
1.2.3 杂交指数估算 按照Dafni[16]的标准进行花序直径、花朵大小和开花行为的测量及繁育系统的评判:(1)花朵或花序直径:<1 mm记为0;1~2 mm记为1;2~6 mm记为2;> 6 mm记为3;(2)花药开裂时间与柱头可授期之间的时间间隔:同时或雌蕊先熟记为0;雄蕊先熟记为1;(3)柱头与花药的空间位置:同一高度记为0;空间分离记为1。
三者之和为OCI值。OCI=0时,繁育系统为闭花受精;OCI=1时,繁育系统为专性自交:OCI=2时,繁育系统为兼性自交;OCI=3时,繁育系统为自交亲和,有时需要传粉者;OCI=4时,繁育系统为部分自交亲和,异交,需要传粉者。
1.2.4 花粉/胚珠比(P/O) 随机选取花蕾尚未开裂的花朵30个,用1 mol·L-1HCl在60 ℃条件下水解1 h去除药壁,花粉粒悬浮液定容至10 mL,用微量移液器吸取2 μL悬浮液于显微镜下观察,统计花粉量,重复10次。取子房在体视显微镜下用解剖刀剖开,统计胚珠数,重复20次。用平均每朵花的花粉量除以平均每朵花的胚珠数得到花粉/胚珠比P/O。按Cruden[17]的标准:P/O为2.7~5.4时,繁育系统为闭花受精;P/O为18.1~39.0时,繁育系统为专性自交;P/O为31.9~396.0时,繁育系统为兼性自交;P/O为244.7~2 588.0时,繁育系统为兼性异交;P/O为2 108.0~195 525.0时,繁育系统为专性异交[18]。
1.2.5 访花昆虫 在异裂菊属植物的盛花期,选晴天观察整个居群的昆虫访花情况,观察记录单花访花昆虫的种类和数量、昆虫访问单花时间长度和访花频率。不定时捕捉访花昆虫作凭证标本[19]。
2 结果与分析
2.1 开花物候、花部特征和开花动态
2.1.1 开花物候 连续2年观察发现,异裂菊属5个种之间开花物候之间存在较大差异。绢叶异裂菊的花期最早,5月中旬为现蕾期,6月下旬为开花旺期,7月中为开花末期,7月至10月为结实期。小花异裂菊的花期较绢叶异裂菊的花期迟20~30 d左右,两者的花期都为2个月左右。柳州异裂菊6月初现蕾,8月上旬为开花旺期,8月底为开花末期。花期结束时间较绢叶和小花异裂菊迟约1个月。凹脉异裂菊和异裂菊的花期基本一致,6月底为现蕾期,8月初为始花期,10月为开花旺期,12月为开花末期,整个花期持续约5个多月。异裂菊属种子成熟时间与花期重合,开花旺期时已有部分种子成熟,种子成熟期仍有零星的开花(表1,图1)。
2.1.2 花部形态特征观察 (1)小花异裂菊 总苞钟状圆柱形,长约8.12 mm,宽约2.26 mm,4层,外层长约2.18 mm,中层长约2.93 mm,内层长约4.01 mm。雌花边缘1层,4~5个,管长约2.53~4.12 mm,舌片极小,花柱高于花面约0.49 mm。两性花位于中间,4个,管长约6.18 mm,舌片长约1.07 mm,花柱高于舌片约2.15 mm。
(2)绢叶异裂菊 总苞钟状圆柱形,长约7.66 mm,宽约3.14 mm,总苞片4层,外层长约2.11 mm,中层长约2.50 mm,内层长约3.17 mm。雌花位于边缘1层,4~5个,管长约3.46 mm,舌片长约0.32 mm,花柱高于舌片约0.84 mm。两性花位于中间,5~6个,花冠管状,管长约5.17 mm,舌片长约1.01 mm,花柱高于舌片约2.16 mm。
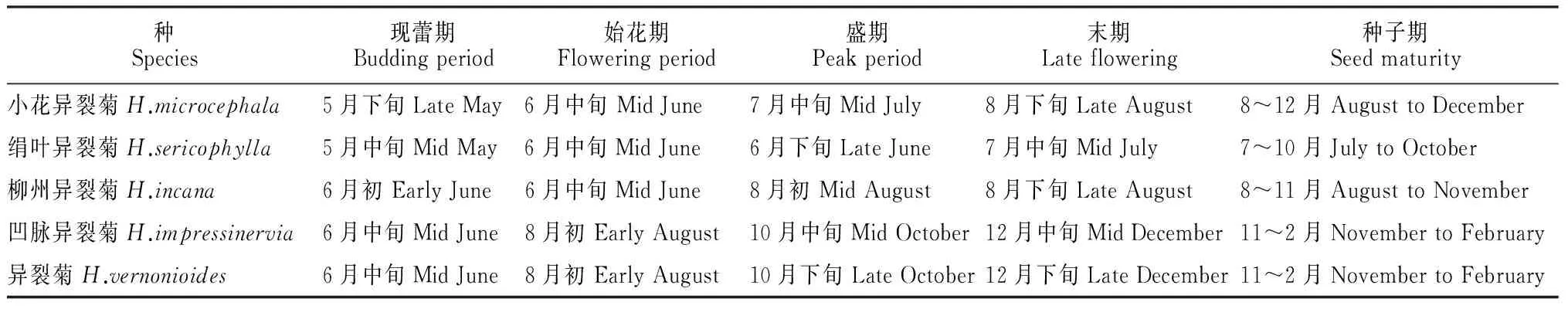
表1 异裂菊属开花物候

图1 柳州异裂菊开花过程
(3)柳州异裂菊 总苞长约10.3 mm,宽约2.58 mm,4~5层,外层长约2.72 mm,中层长约3.32 mm,内层长约4.22 mm。雌花边缘1层,3~4个,管长约3.76 mm,舌片约5 mm,花柱高于舌片0.6 mm。两性花位于中间,3~4个,管长5.18~6.33 mm,舌片长约5 mm,柱头高于舌片约2.58 mm。
(4)凹脉异裂菊 总苞长约8.59 mm,宽约2.13 mm,4~5层,外层长约4.29 mm,中层长约3.71 mm,内层长约2.69 mm。雌花边缘1层,4个,管长约3.93 mm,舌片极小,柱头高于花面约0.81 mm。两性花位于中间,4~5个,管长约5.52 mm,舌片长约1.02 mm,柱头高于花面约0.75 mm。
(5)异裂菊 总苞长约10.46 mm,宽约2.36 mm,5层,外层长约5.97 mm,中层长约3.64 mm,内层长约2.93 mm。雌花边缘1层,多为4个,管长约5.48 mm,柱头高于花药长约1.71 mm。两性花位于中间,2~4个,管长约4.84 mm,柱头高于花药长约1.24 mm。
2.1.3 开花动态 异裂菊属为头状花序,单生于枝端叶腋或2~3个簇生于茎或分枝的顶端。开花顺序为从上到下,花序顶端最先开放,然后渐及于两侧开放。主干上的各枝节或主干枝节的副枝节上的花开放无一定的顺序。边缘花为雌性单性花,中间为两性花。边缘雌性花先开,在两性花开花前,雌性花已有2~3个柱头展开。两性花开放时间不一,一般2~3个同时先开,持续3 d左右,1~2个后续开放。两性花开花后,柱头有很多花粉,花柱和柱头呈亮黄色。约18 h后花粉明显减少,花柱开始变为黄褐色。约24 h后,花柱仅有少量花粉,呈青色,花柱绝大部分变为黄褐色。40 h后,基本无花粉,花药变为浅绿色,花柱全部变为褐色,花药前段分叉明显。
2.2 花粉活力和柱头可授性
异裂菊属花粉在花药开裂时花粉活力处于最强状态,此后随着时间的推移花粉活力逐渐下降,18 h后仅剩少量花粉,剩余的花粉基本无活力。柱头在花药开裂前具有可授性,花药开裂后2 h左右柱头可授性最强,随着时间推移柱头可授性下降,可授性可持续到开花后48 h左右(表2)。
2.3 杂交指数
异裂菊属花序直径全部大于6 mm,OCI记为3;雌蕊先熟,OCI记为0;花药与柱头空间分离,OCI记为1。所以,异裂菊属的杂交指数(OCI)为4。按照Dafni(1992)的标准判断:异裂菊属的有性繁育系统为部分自交亲和,异交,需要传粉者。
2.4 花粉胚珠比
表3显示,异裂菊属平均每朵花的花粉数在1 450~2 250粒之间,胚珠数每朵花1个,P/O值为1 450~2 250。按照Cruden(1977)的标准,异裂菊属的繁育系统为兼性自交或兼性异交。
2.5 访花与传粉昆虫
异裂菊属的访花昆虫主要有眼蝶科稻眉眼蝶(MycalesisgotamaMoore)、寄蝇科日本追寄蝇(Exoristajaponica)、食蚜蝇科灰带管食蚜蝇(Eri-staliscerealis)、胡蜂科黑尾胡蜂(Vespaducalis)、蚁科哀弓背蚁(CamponotusdolendusForel)、灰蝶科亮灰蝶[(Lampidesboeticus(Linnaeus)]、丽蝇科丽蝇(Calliphoridae)、蝽科大臭蝽[Metonymiaglandulosa(Wolff)]等。观测结果显示异裂菊属的传粉昆虫主要是食蚜蝇科灰带管食蚜蝇和胡蜂科黑尾胡蜂等。
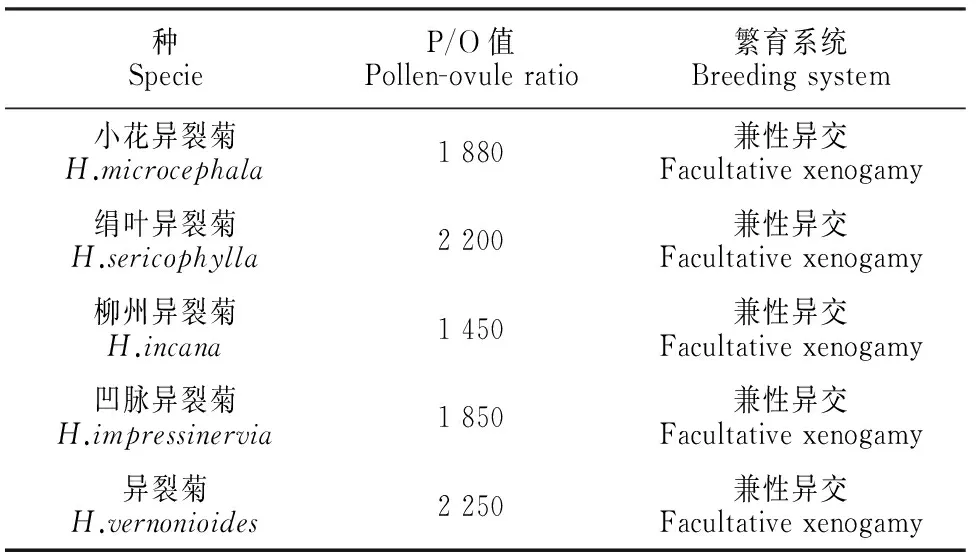
表3 异裂菊属花粉-胚珠比
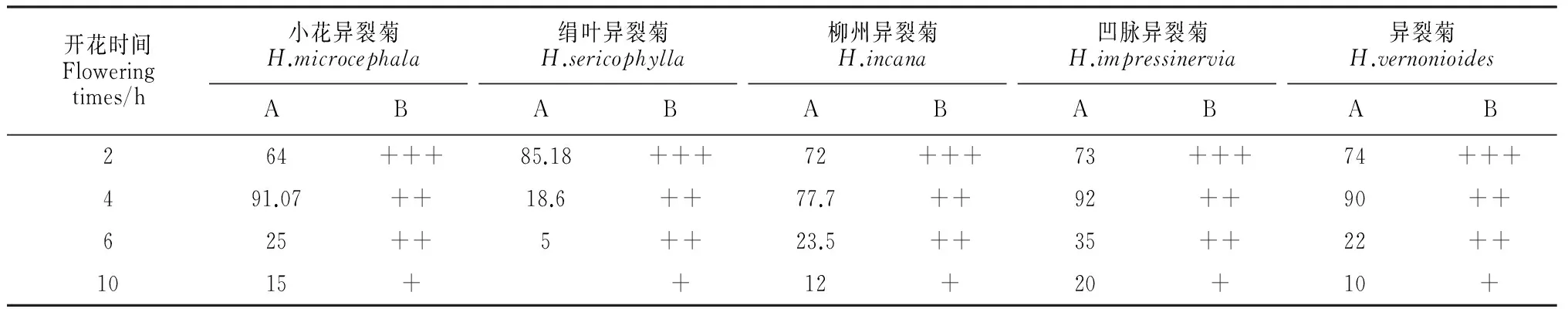
表2 异裂菊属花粉活力和柱头可授性
注:A.花粉活力;B.柱头可授性;+++表示柱头具最强可授性;++柱头具较强可授性;+表示具有可授性。
Note:A.Pollen activity;B.Stigma receptivity;+++ means stigmas have the highest receptivity;++ means stigmas have the higher receptivity;+ means sigmas have receptivity.
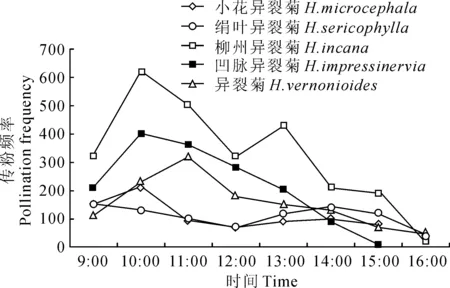
图2 主要传粉昆虫访花统计
由图2可知,昆虫访花集中在10:00~14:00 期间,访花高峰期在中午10:00~12:00。食蚜蝇和黑尾胡蜂等访花时,头腹部可以接触到花药,成熟的花粉粒粘附在昆虫头腹部,昆虫离开时花粉即被带走。昆虫每次可连续访问多朵花,每朵花访问时间约为2~10 s。
3 讨 论
异裂菊属为头状花序,平均9朵单花聚合在一起组成,单生于枝端叶腋或2~3个簇生于茎或分枝的顶端。异裂菊属在开花过程中,花的颜色由淡黄色渐变成褐色,花冠较小,不具芳香气味,单花寿命为2~3 d不等,单花开放不同步。这种集成花序结构的花朵对昆虫的吸引比单花效应的总和要高[20],开花持续时间长且开花不同步。一方面保证植株在较长的时间内能成功完成异花传粉受精过程而保证生殖成功,另一方面也可降低花期恶劣的自然环境对其生殖成功造成的不良影响,成为其长期适应环境影响所形成的一种生殖策略,这与许多研究结果相吻合[2,21-22]。
按照Dafni等建议的标准进行OCI测定,异裂菊属的繁育系统为异交、部分自交亲和、需要传粉者;依据Cruden以花粉/胚珠比评判繁育系统的标准,异裂菊属的繁育系统属于兼性异交类型。2种方法检测到的繁育系统结果基本一致。异裂菊属的繁育系统为异交、部分自交亲和、需传粉者。
自交不亲和、雌雄异位和雌雄异熟是促进异交,避免单花自交的重要机制[22]。异裂菊属具有保证异交进行的许多特征,由于雌花不产生花粉,雌花和两性花分离,降低了花粉落置在雌花柱头上自交的比例。而且,在异裂菊属中雌花位于两性花的边缘一轮,传粉昆虫的运动方式是由外向内,边缘的雌花由于被访花的传粉者优先访问其异交率增大[23]。
从柱头的可授性强弱变化与花粉活性变化规律来看,在异裂菊属花药刚开裂前,雌花的柱头已经成熟。当两性花的花药开始枯萎时,柱头都具有微弱可授性。由此看出柱头的可授性好,花粉活力较差,其柱头具有可授性时期与花粉成熟期具有部分重叠期,雌蕊的成熟先于雄蕊,其适应异花传粉的机制。
异裂菊属的主要有效传粉媒介为食蚜蝇科灰带管食蚜蝇和胡蜂科黑尾胡蜂等昆虫,访花主要集中在10:00~12:00。许多因素如光照、大风、温度、阴雨天气等都可以影响访花者的数量、行为和频率并进而影响植物的传粉和结实。食蚜蝇和胡蜂的活动受气候条件的影响明显,上午12:00前数目较多,活动频率相对高,当温度升高和光照增强后,访问频率低。异裂菊属分布于石灰岩石山山顶,其开花期山顶温度可以高达40 ℃左右且持续时间长,光照强烈,白天和夜晚的风速都相对较高。石灰岩石山山顶气候下影响传粉的主要气象因子是风速和温度。强风和高温会导致柱头表面干燥和花蜜快速蒸发,使传粉者的活动减少。如果盛花期遇到持续阴雨天气,雨水会冲掉花粉并使昆虫活动减少。气象因素可能还会影响生境昆虫区系,进而影响访花者的种类和数量。表明异裂菊属具有不稳定的传粉环境,成为异裂菊属能否成功传粉的重要影响因子[2]。
自交衰退作为物种的濒危机制的重要原因已在很多物种中有过报道[24]。一方面,异裂菊属的雌雄同花可进行自发自交,另一方面,传粉昆虫也可对异裂菊属进行协助自交。异裂菊属居群仅分布于石灰岩石山山顶部分,两个居群之间距离较远。传粉昆虫很难在不同居群之间进行飞行。异裂菊属居群的植株数量一般在10~50之间,其中只有部分开花,昆虫随机的访花方式极易引起同株异花授粉,从而导致自交。异裂菊属自然分布十分狭窄,居群规模很小,长期的自交在一定程度上导致居群遗传多样性降低[21]。野外调查发现,异裂菊属小苗非常少,表明其居群自然更新能力很差,暗示异裂菊属可能存在近交衰退[2]。异裂菊属的有性繁殖能力低,种群复壮最主要的途径就是提高有性繁殖能力。因此,保存现有异裂菊属居群面积或者人工促进居群内与居群间的传粉,增加后代的基因杂合度,可在很大程度上达到复壮种群的目的。
[1] WANG J(王 洁),YANG ZH L(杨志玲),YANG X(杨 旭).Advance in the studies of endangered plants breeding system[J].JournalofNorthwestA&FUniversity(Nat.Sci.Edi.)(西北农林科技大学学报·自然科学版),2011,39(9):207-213(in Chinese).
[2] WANG Y B(王玉兵),LIANG H W(梁宏伟),MO N B(莫耐波),etal.Flower phenology and breeding system of rare and endangeredDayaoshaniacotinifolia[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2011,31(5):958-965(in Chinese).
[3] GAO T T(高婷婷),LI Q H(李清河),XU J(徐 军),etal.Sexual reproduction system characteristics of rare eremophyteHelianthemumsoongoricum[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2010,30(10):1 982-1 988(in Chinese).
[4] WANG L L(王立龙),WANG G L(王广林),LIU D Y(刘登义),etal.Pollination biology of endangeredMagnoliasieboldii[J].ChineseJournalofEcology(生态学杂志),2005,24(8):853-857(in Chinese).
[5] LIU L D(刘林德),TIAN G W(田国伟).The pollination biology ofEleuthero-coccussenticosus(Araliaceae)[J].ActaPhytotaxonomicaSinica(植物分类学报),1998,36(1):19-27(in Chinese).
[6] QI W Q(奇文清),CHEN X L(陈晓麟).Pollination biology inCimicifugananchuanensis,an endangered species(Ranunculaceae)[J].ActaBotanicaSinica(植物学报),1998,40(8):688-694(in Chinese).
[7] HUANG SH Q(黄双全),GUO Y H(郭友好),CHEN J K(陈家宽).Pollination rates and pollen tube growth in a vulnerable plant,liriodendronchinense(Hemsl.) Sarg.(Magnoliaceae)[J].ActaPhytotaxonomicaSinica(植物分类学报),1998,36(04):23-29(in Chinese).
[8] LIANG J Y(梁健英).Two new species ofHeteroplexisChang(Compositae)[J].Guihaia(广西植物),1994,14(2):126-129(in Chinese).
[9] WANG C M(王才明),HUANG SH X(黄仕训),WANG Y(王 燕).A preliminary study on ecological and biologcal characteristics ofHeteroplexismicrocephala——A protected and endemic species from Guangxi[J].Guihaia(广西植物),1994,14(3):277-288(in Chinese).
[10] 傅立国.中国红皮书——稀有濒危植物[M].北京:科学出版社,1992:228-229.
[11] ZHANG G L(张国莉),GUO X(龚 洵).The karyotype analysis ofAnemoclemaglaucifoliumandHeteroplexismicrocephalaboth endemic to China[J].ActaBotanicaYunnanica(云南植物研究),2002,24(6):765-768(in Chinese).
[12] FAN X N(樊晓娜),LIN SH(林 生),ZHU CH G(朱承根),etal.Terpenoids ofHeteroplexismicocephalaand their bioactivities[J].ChinaJournalofChineseMateriaMedica(中国中药杂志),2010,35(3):315-322(in Chinese).
[13] HUANG SH X(黄仕训),LUO W H(骆文华),TANG W X(唐文秀),etal.Study on the adaptability of the rare and threatenedl imestone plants ex- situ conservation[J].Guihaia(广西植物),2002,22(02):136-139(in Chinese).
[14] GAO Z R(高则睿),ZHANG CH Q(张长芹),HAN ZH Q(韩智强).Pollination biology and breeding system ofCraigiayunnanensisin fragmented habitat[J].ChineseJournalofEcology(生态学杂志),2012,31(9):2 217-2 224(in Chinese).
[15] ZHANG Y(张 洋),YE Q G(叶其刚).Breeding system of the endangered speciesPsilopeganumsinenseand its pollination process in a botanical garden[J].PlantScienceJournal(植物科学学报),2011,29(5):599-606(in Chinese).
[16] DAFNI A.Pollination Ecology[M].New York:Oxford Unit Press,1992:1-57.
[17] CRUDEN R W.Pollen-ovule ratio:A conservative indicator of breeding systems in flowering plants[J].Evolution,1977,31:32-46.
[18] JIANG B(姜 波),SHEN Z G(沈宗根),RUAN X L(阮仙利),etal.Floral biology and breeding system ofHypericumperforatum[J].Guihaia(广西植物),2012,32(04):457-463(in Chinese).
[19] HU H Y(胡红岩),CHEN H(陈 欢),XU H L(徐环李).Main pollinators and their foraging behaviors on a sand-fixing legume,Thermopsislanceolata,in Mu Us Sandland[J].BiodiversityScience(生物多样性),2012,20(3):354-359(in Chinese).
[20] MULLIGAN G A,KEVAN P G.Color,brightness,and other floral characteristics at tracting in sects to the blossoms of some Canadian weeds[ J].CanadianJournalofBotany,1973,51:1 939-1 952.
[21] ZHANG J J(张金菊),YE Q G(叶其刚),YAO X H(姚小洪),etal.Preliminary studies on the floral biology,breeding system and reproductive success ofSinojackiahuangmeiensis,an endangered plant in a fragmented habitat in Hubei Province,China[J].JournalofPlantEcology(Chinese Version)(植物生态学报),2008,32(4):743-750(in Chinese).
[22] LÜ H Y(吕海英),CAO M H(曹满航),LI J(李 进).Study on the breeding system of the rare plantAmmodendronargenteum(Pall.) Kuntze[J].Guihaia(广西植物),2014,34(6):763-767(in Chinese).
[23] LU Y(卢 洋),HUANG SH Q(黄双全).Adaptive advantages of gynomonoecious species[J].ActaPhytotaxonomicaSinica(植物分类学报),2006,44(2):231-239(in Chinese).
[24] SHI X(施 翔),LIU H L(刘会良),ZHANG D Y(张道远),etal.The flower sydrome and pollination adaptation of desert rare speciesEremospartonsongoricum(litv) Vass.(Fabaceae)[J].ActaEcologicaSinica(生态学报),2013,33(18):5 516-5 522(in Chinese).
(编辑:潘新社)
Study on the Floral Biology and Breeding System ofHeteroplexisPlants
SHI Yancai,ZOU Rong,TANG Jianmin,CHEN Zongyou,CHAI Shengfeng,WEI Xiao*
(Guangxi Institute of Botany,Guangxi Zhuangzu Autonomous Region and the Chinese Academy of Sciences,Guilin,Guangxi 541006,China)
Field investigation was performed on the floral syndrome,pollination characteristics,pollinator behaviors and breeding system ofHeteroplexisby continuous observations,TTC method,Benzidine-H2O2method,out-crossing index,pollen-ovule ratio.The result as follows:(1)Under natural conditions,the flowering stage ofHeteroplexisvernonioideswas about 60 days from April to June,HeteroplexismicrocephalaandHeteroplexisincanawere about 60 days from April to August,HeteroplexisimpressinerviaandHeteroplexisvernonioideswere about 150 days from August to December.The life span of a single flower was about 3 days and the inflorescence was 5 days.(2)Pollen had vigor after the 1-18 h of anthesis.Stigma was receptive after 48 hours of anthesis.The out-crossing index was 4,P/O was 1 450-2 250.The breeding system of five species were given priority to out-crossing,part self-compatibility,insects pollinator dependent.(3)Eristalis cerealis and vespa ducalis are effective pollinators.The visiting frequency showed triple-climax was 10:00-12:00 and the visiting time was 2-10 s per flower.
Heteroplexis;floral syndrome;out crossing index;pollen-ovule ratio;breeding system
1000-4025(2015)04-0824-06
10.7606/j.issn.1000-4025.2015.04.0824
2014-10-27;修改稿收到日期:2015-03-05
广西自然科学基金项目(2013GXNSFBA019054;2014GXNSFAA118093);广西植物研究所基本业务费项目(桂植业12011)
史艳财(1984-),男,硕士,助理研究员,主要从事保护生物学和药用植物引种驯化研究。E-mail:shiyancainan@163.com
*通信作者:韦 霄,研究员,主要从事保护生物学研究。E-mail:weixiao@gxib.cn
文献标志码:A
猜你喜欢
中国科学探险(2022年8期)2022-12-31
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
铜仁学院学报(2018年7期)2018-09-08
大科技·百科新说(2018年3期)2018-04-11
当代工人(2018年21期)2018-03-06
现代园艺(2017年22期)2018-01-19
中国稻米(2017年6期)2017-12-28
红蜻蜓·低年级(2017年6期)2017-10-30
读写算·小学低年级(2017年8期)2017-08-11
作物研究(2015年4期)2015-03-24
