
黑龙江苹果亚科5属14种植物导管分子形态结构研究
2015-02-28 02:12刘树焕谷利伟张大维
西北植物学报 2015年4期
刘树焕,谷利伟,张大维
(哈尔滨师范大学 生命科学与技术学院,哈尔滨 150025)
黑龙江苹果亚科5属14种植物导管分子形态结构研究
刘树焕,谷利伟,张大维*
(哈尔滨师范大学 生命科学与技术学院,哈尔滨 150025)
利用扫描电子显微镜对黑龙江苹果亚科5属14种植物导管分子的管腔微形态结构特征进行了比较研究。结果显示:(1)该亚科植物导管分子的管腔长度、宽度及端壁斜度角有较大的差别。(2)孔纹导管在该亚科植物种中均存在,网纹导管和螺纹导管仅见于苹果属和花楸属中。(3)导管分子管壁除苹果属及山楂属中的毛山楂、辽宁山楂外均有螺纹加厚。(4)纹孔的排列方式为互列式、对列式及互列、对列同时存在。(5)在所观察的植物中花楸、山荆子、无毛山楂导管端壁具单穿孔板和梯状穿孔板,其余种的导管端壁仅具单穿孔板。(6)导管纹孔膜残留现象普遍存在。研究表明,导管分子管腔的微形态结构特征,可为该亚科植物的系统演化提供形态学依据;导管分子微形态结构特征与其所处的环境存在一定的适应性。
苹果亚科;形态结构;导管分子;扫描电镜;黑龙江
苹果亚科(Maloideae)隶属于蔷薇科(Rosaceae),是以果实特化为梨果为标志的植物类群[1]。该亚科共20属,主要分布于北半球,中国有16属,黑龙江有5属,12种,2变种,1变型[2]。有关该亚科植物形态结构的研究国内外有一些报道,Huckins[3]从形态解剖学特征出发,研究苹果属(MalusM.)植物的进化,认为多胜海棠组(Sect.Docyniopsis Schneid.)是该属中最原始的组;杨晓红等[4-5]通过对苹果属植物花粉形态结构的研究认为苹果系在该属中是较进化的类型;俞德浚[6]通过对苹果亚科植物叶、花形态及果实类型的研究,认为花楸属(SorbusL.)是苹果属的祖先属且梨属(PyrusL.)的演化位置处于二者之间,栒子属(CotoneasterB.)的演化水平高于山楂属(CrataegusL.);刘静等[7]利用扫描电子显微镜对苹果亚科叶表皮微形态特征进行研究,认为梨属要比花楸属、苹果属更为进化。本研究利用扫描电子显微镜对黑龙江苹果亚科14种植物的导管分子管腔主要特征进行比较,旨在为该亚科植物的系统演化提供导管微形态学依据,并从导管分子微形态特征的角度对其与环境的适应性进行探讨。
1 材料和方法
1.1 供试样品
样品均为2013年采自黑龙江境内,取样部位为1年生枝条的节间处。采集相关信息见表1。
1.2 方 法
1.2.1 复型导管样品结构观察 选取一年生枝条节间处,切成10 mm×5 mm大小,用苯乙烯渗透4~6周。渗透好的样品置于60 ℃温箱中使其完全凝固,用刀片切除多余树脂,并将样品切成细杆状后,浸泡于下列2种溶液中,A为高浓度的过氧化氢及等量的冰醋酸混合液,B为浓硫酸。A,B两步骤交替进行,直至充分暴露出复型样品。样品经自然干燥后,粘贴于扫描电镜样品台上,Eiko IB-3离子溅射仪中进行导电处理,HITACHI S-4800型扫描电子显微镜下(加速电压15 kV)进行观察,随机测量30个导管分子管腔长度、宽度、端壁斜度角,求其平均值和标准差。
1.2.2 离析导管样品结构观察 将样品放入装有解离液(10%铬酸与10%硝酸等量混合液)的瓶中,置于60 ℃温箱,每间隔1 h更换1次解离液,时间3~6 h。待样品完全软化,并呈白色透明状,解离完成。解离好的样品吸出解离液,用蒸馏水冲洗后自然干燥粘贴于扫描电镜样品台上,置Eiko IB-3离子溅射仪中进行导电处理,Hitachi S-4800型扫描电子显微镜下(加速电压15 kV)进行观察。
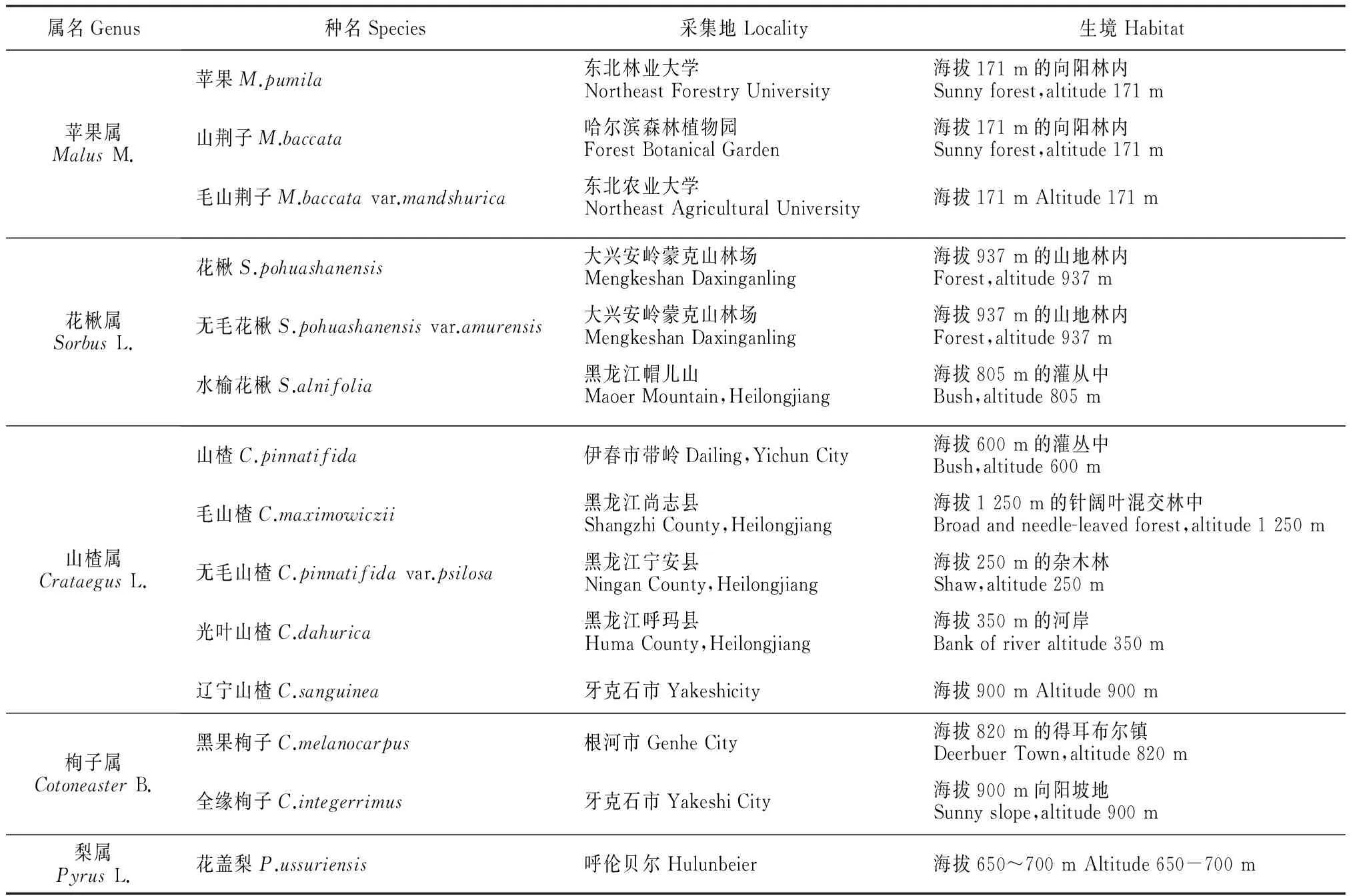
表1 黑龙江苹果亚科14种植物材料信息
2 观察结果
苹果亚科植物导管分子的管腔长度、宽度及端壁斜度角有较大的差别(表2);导管有网纹、孔纹、螺纹3种形态类型,导管次生壁内表面出现螺旋状凸起的螺纹加厚,纹孔排列方式为互列式、对列式及对列、互列同时存在,导管端壁穿孔板为单穿孔板、梯状穿孔板(表3);导管纹孔膜残留现象普遍存在。

表2 扫描电镜下苹果亚科14种植物导管分子管腔的长度、宽度及端壁斜度角

表3 黑龙江苹果亚科14种植物导管分子形态结构特征
注:+和-分别表示有和无。
Note:+and - stand for have and have not.
2.1 导管分子长度、宽度、端壁斜度角
导管分子长度在263.87~436.97 μm之间,其中最长的是毛山楂,最短的是山楂;导管分子管腔宽度范围17.27~27.30 μm,其中光叶山楂最宽,黑果栒子最窄;端壁斜度角在45.25°~61.79°之间,其中毛山楂最大,辽宁山楂最小。
2.2 导管形态类型
导管形态类型共有3种,网纹导管、螺纹导管和孔纹导管。3种类型导管都存在的有山荆子和毛山荆子(图版Ⅰ,1~4);存在螺纹导管和孔纹导管的只有一种,即花楸(图版Ⅰ,5);其余的种均只有孔纹导管(图版Ⅰ,6、7)。
2.3 导管管壁的螺纹加厚
导管分子内表面呈螺纹加厚(图版Ⅰ,8),除苹果属和山楂属中辽宁山楂、毛山楂无螺纹加厚(图版Ⅰ,9)外,其余种均有螺纹加厚。
2.4 纹孔的排列方式
纹孔排列方式为对列式(图版Ⅱ,1)、互列式(图版Ⅱ,2)以及对列、互列同时存在(图版Ⅱ,3)3种类型,其中苹果属、花楸、水榆花楸为对列式,山楂属、花盖梨为互列式排列,无毛花楸、栒子属为对列式、互列式同时存在。
2.5 导管端壁穿孔板类型
在所观察的植物中花楸、山荆子、无毛山楂具单穿孔板和梯状穿孔板(图版Ⅱ,4、5),其余种仅具单穿孔板(图版Ⅱ,6~8)。
2.6 纹孔膜残留
所观察植物木质部导管纹孔膜残留(图版Ⅱ,9)的现象普遍存在。
3 讨 论
3.1 导管微形态特征
导管是植物木质部输导水分和无机盐的管状结构,其输导作用和它的结构特征是紧密联系的。对该亚科植物导管分子测量结果其长度在263.87~436.97 μm之间,管腔宽度变化范围在17.27~27.30 μm之间,端壁斜度角的大小在45.25°~61.79°之间。植物导管分子长度变化幅度相对较大,管腔宽度变化幅度小,端壁斜度角的变化范围不大。斜度角大小直接影响管腔端壁的接触面积,端壁接触面积的大小与植物运输水分的效率呈负相关;孔纹导管在所有植物中均存在,螺纹导管存在于少数种中,网纹导管仅见于苹果属中;纹孔的排列方式为对列式或互列式,同时存在两种排列方式的植物较少;导管管腔内壁螺纹加厚普遍存在,仅少数种中不加厚,螺纹加厚可以加大管壁与水的粘着力,提高植物对水分运输的效率;端壁穿孔板多为单穿孔板,仅少数种中见梯状穿孔板,穿孔板类型的进化可为植物系统演化提供形态学依据;导管纹孔膜普遍存在残留现象,纹孔膜的残留是导管分子原始性的一种表征。
3.2 导管微形态特征与演化
导管是植物适应陆生环境而形成的输导水分和无机盐的最主要组织,它们的结构在系统发育上具有重要地位[8]。在系统演化上,苹果亚科植物导管分子由于发育的先后和侧壁次生增厚的方式不同,形成了螺纹、网纹、孔纹3种类型的导管,其中螺纹导管在个体发育中出现较早,其次是网纹导管,最后发展到孔纹导管,这个规律在个体发育不同时期的木质部中可得到体现[9-10]。管间纹孔式进化的过程为对列→互列(Takhtajan,1948),从这一点来看,互列式纹孔排列是较进化的类型。导管端壁穿孔板的进化顺序为横隔多的梯状穿孔板→单穿孔板(Bailey,1944)且纹孔膜的缺失是导管分子成熟的标志[11-12]。本研究结果显示,导管形态类型以孔纹导管为主,在部分种中有螺纹导管、网纹导管的出现;管间纹孔排列方式既有原始的对列式又有较进化的互列式,也有两种排列方式同时存在的;端壁单穿孔板及纹孔膜的残留见于所有研究的植物种中,梯状穿孔板仅存在于少数种中。从同一亚科植物既出现原始又有进化的导管形态特征来看,本文认为苹果亚科的演化水平处于相对较进化的初级阶段。
3.3 导管微形态特征与其生境适应性
导管位于维管植物的木质部中,是绝大多数被子植物输导水分和矿质元素的重要组织。导管管腔宽度较小,长度较短(即单位长度中横壁较多)端壁斜度角较小(即端壁的面积较大)等是加大水分运输阻力的因素[8]。导管管腔的长度、宽度及端壁斜度角与生境是密切相关的,高海拔和土壤水分缺乏环境下的植物导管一般较长、管腔相对较宽、端壁斜度角较大;低海拔和土壤水分充足环境下导管较短、管腔相对较窄、端壁斜度角较小[13-15]。从导管分子的长度、宽度和端壁斜度角结果显示,毛山楂导管分子最长,管腔相对较宽,端壁斜度角最大,这些形态特征是植物适应海拔较高水分相对较缺乏的土壤环境中的表现。辽宁山楂导管分子相对较短,管腔相对较窄,端壁斜度角最小是其长期生活在低海拔地区和水分含量相对充沛的土壤环境中的表现。其余种类导管分子长度、宽度、端壁斜度角差异明显,说明苹果亚科植物有着广泛的分布地区,这种结构特点差异是其功能长期适应不同环境的结果。
导管分子管壁的螺纹加厚,可以增加管腔表面积,加大管壁与水的粘着力,提高管内水分的内聚力,从而保证水分更有效的运输[16-18],管壁有螺纹加厚的植物,生活在水分含量较多的地区。研究结果显示,苹果亚科导管分子管壁螺纹加厚普遍存在(除苹果属、毛山楂、辽宁山楂),植物导管管壁螺纹加厚这一特征是植物长期适应环境的结果。
[1] 聂绍荃,张艳华.黑龙江省植物志[M].哈尔滨:东北林业大学出版社,1998:81-160.
[2] 周以良,董世林.黑龙江树木志[M].哈尔滨:黑龙江科学技术出版社,1986:267-354.
[3] LI Y N(李育农).Progress in research on the origin and evolution of genusMalusin the word[J].JournalofFruitScience(果树科学),1999,16(S):8-19(in Chinese).
[4] YANG X H(杨晓红),LI Y N(李育农).Studies on morphology and evolution of sect.Maluslangenf and sect.baccatus jiang of genusMalusmill through the pollen grain analysis[J].JournalofSouthwestAgriculturalUniversity(西南农业大学学报),1995,17(4):279-285(in Chinese).
[5] YANG X H(杨晓红),LI Y N(李育农).Studies on the pollen morphology,systematics of genusMalussect.Sorbomalus zabel.and sect,docyniopsis[J].JournalofSouthwestAgriculturalUniversity(西南农业大学学报),1995,17(4):348-354(in Chinese).
[6] YU D J(俞德浚).Origins and evolution of Rosaceae[J].ActaPhytotaxonomicaSinica(植物分类学报),1984,2(6):431-444(in Chinese).
[7] LIU J(刘 静),WANG L(王 磊).Morphology structure of leaf epidermis of Maloideae in Heilongjiang[J].JournalofChineseElectronMicroscopySociety(电子显微学报),2014,33(1):69-76(in Chinese).
[8] ZHANG D W(张大维),SHI F CH(石福臣).Morphological anatomical studies on Betulaceae vessel element from Heilongjiang Province,China[J].BulletinofBotanicalResearch(植物研究),2004,24(2):158-163(in Chinese).
[9] YIN X L(尹秀玲),WANG J X(王金侠).The anatomy observation of the vessel type of the twelve kinds of plants in Rosaceae[J].JournalofHebeiNormalUniversityofScience&Technology(河北科技师范学院学报),2006,20(4):14-17(in Chinese).
[10] POOLE I,FRANCIS J E.The first record of fossil wood of Winteraceae from the upper cretaceous of Antarctica[J].AnnalsofBotany,2000,85:307-315.
[11] LI H F(李红芳),REN Y(任 毅).The variation of perforation plates of vessels in the secondary xylem ofEupteleapleiosperma(Eupteleaceae)[J].ActaPhytotaxonomicaSinica(植物分类学报),2005,43(1):1-11(in Chinese).
[12] ZHU J Y(朱俊义),LU J M(陆静梅).Anatomic study of the secondary xylem inOplopanaxelatussetms[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2005,25(4):776-780(in Chinese).
[13] LI H F(李红芳),TIAN X H(田先华),REN Y(任 毅).Research progress in vessel and perforation plate of vascular plants and some considerations for future research[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2005,25(2):419-424(in Chinese).
[14] ZHU J Y(朱俊义).The type and evolution of vessel element’s penforation plate ofSorbuspohuashanensis[J].BulletinofBotanicalResearch(植物研究),2002,22(3):285-288(in Chinese).
[15] CHEN SH S(陈树思).Study of vessel elements inPterisvittata[J].JournalofWuhanBotanicalResearch(武汉植物学研究),2005,23(2):196-198(in Chinese).
[16] CARLQUIST S.Ecologiical strategies of xylem evolution[M].California:University of Californiusa Press,1975.
[17] CARLQUIST S,SCHNEIDER E L.Origin and natures of vessels in monocotyledons.5.Araceae subfamily Colocasioideae[J].BotanicalJournaloftheLinneanSociety,1998,128:71-86.
[18] CARLQUIST S.Wood anatomy of coriariaecae:phylogenetic and ecological implications[J].Syst.Bot.,1985,10:174-183.
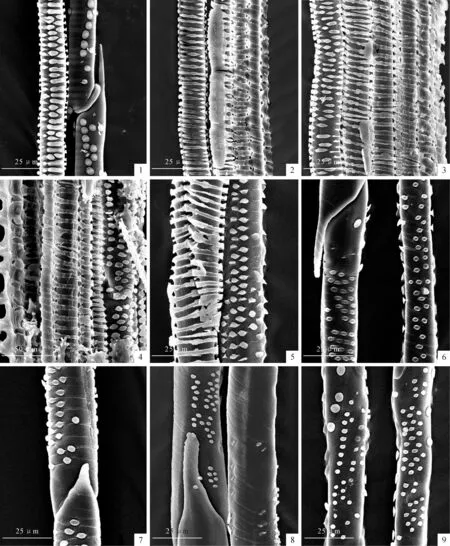
图版 Ⅰ 扫描电镜下导管分子管腔微形态
1.山荆子,×600;2.山荆子,×600;3.毛山荆子,×600;4.毛山荆子,×1000;5.花楸,×600;6.全缘栒子,×600;7.黑果栒子,×600;8.光叶山楂,×600;9.辽宁山楂,×600。
Plate Ⅰ SEM photomicrographs of the inner wall of the vessel molecules
Fig.1.M.baccata,×600;
Fig.2.M.baccata,×600;
Fig.3.M.baccatavar.mandshurica,×600;
Fig.4.M.baccatavar.mandshurica,×1 000;
Fig.5.S.pohuashanensis,×600;
Fig.6.C.integerrimus,×600;
Fig.7.C.melanocarpus,×600;
Fig.8.C.dahurica,×600;
Fig.9.C.sanguinea,×600.

图版 Ⅱ 扫描电镜下导管分子管腔微形态
1.苹果,×600;2.花盖梨,×500;3.无毛花楸,×600;4.无毛山楂,×600;5.无毛山楂,×600;6.水榆花楸,×250;7.毛山楂,×600;8.山楂,×600;9.花盖梨,×1 000。
Plate Ⅱ SEM photomicrographs of the inner wall of the vessel molecules
Fig.1.M.pumila,×600;
Fig.2.P.ussuriensis,×500;
Fig.3.S.pohuashanensisvar.amurensis,×600;
Fig.4.C.pinnatifidavar.psilosa,×600;
Fig.5.C.pinnatifidavar.psilosa,×600;
Fig.6.S.alnifolia,×250;
Fig.7.C.maximowiczii,×600;
Fig.8.C.pinnatifida,×600;
Fig.9.P.ussuriensis,×1 000.
(编辑:潘新社)
Morphological Structure of Vessel Elements in 14 Speciesof Maloideae (Rosaceae) from Heilongjiang,China
LIU Shuhuan,GU Liwei,ZHANG Dawei*
(College of Life Science and Technology,Harbin Normal University,Harbin 150025,China)
The morphological structure of the inner wall of the vessel elements in 14 species of Maloideae (Rosaceae) from Heilongjiang Province was studied by means of scanning electronic microscope (SEM).The result showed that:(1)The length,the width and the slope of vessel elements are various in different species;(2)All studied species have pitted vessel type,while reticulated and spiral vessel only appear in theMalusandSorbus;(3)Helical thickening of the inner wall of the vessel elements occurs in most species except inMalusand inCrataegusmaximowicziiandC.sanguinea;(4)The arrangement of pits is alternate,opposite or both in one element;(5)Simple and scalariform perforations were found on the end wall of the vessel elements inSorbuspohuashanensis,MalusbaccataandCrataeguspinnatifidavar.psilosa,while other species only with simple perforations on the end wall of the vessel elements;(6)The remnants of pit membrane are common in all species.These results indicated that the morphological structure of the inner wall of the vessel elements is systematic significance for Malodieae,and the micro structure of the vessel elements is also adapted to their environment.
Maloideae;morphological structure;vessel element;SEM;Heilongjiang
1000-4025(2015)04-0723-07
10.7606/j.issn.1000-4025.2015.04.0723
2014-10-21;修改稿收到日期:2015-03-05
哈尔滨师范大学硕士研究生创新科研项目(HSDSSCX2014-31);哈尔滨师范大学科技发展预研项目(11XYG-13)
刘树焕(1988-),女,在读硕士研究生,主要从事植物结构形态特征的研究。E-mail:liushuhuan198833@126.com
*通信作者:张大维,博士,教授,主要从事植物解剖学科研工作。E-mail:daniel7491@126.com
Q944.5
A
猜你喜欢
影像研究与医学应用(2022年23期)2023-01-11
粮食加工(2021年6期)2021-12-03
护理实践与研究(2020年23期)2021-01-04
石油钻探技术(2020年3期)2020-08-24
河南医学研究(2020年4期)2020-02-26
武夷科学(2019年2期)2019-12-20
中国医学装备(2019年11期)2019-11-27
石油地质与工程(2019年4期)2019-09-10
中成药(2018年3期)2018-05-07
西南石油大学学报(自然科学版)(2016年2期)2016-12-01
