
吉林西部退化草地中羊草的生长率和异速生长关系的研究
2015-02-27 09:00赵一嬴李月芬郭冬艳王冬艳李晴宇
安徽农业科学 2015年24期
赵一嬴,李月芬* ,郭冬艳,2,王冬艳,李晴宇
(1.吉林大学地球科学学院,吉林长春130061;2.中国国土资源经济研究院,北京101149)
生态化学计量学是从元素计量的角度研究生态过程和生态作用中能量和化学元素平衡的科学,是当前生态学领域的研究热点。生长率假说是当前生态化学计量学研究的热点问题,该假说认为生物个体的生长速率与体内的氮磷比(N∶P)、碳磷比(C∶P)具有负相关的关系,与体内的P含量呈显著的正相关关系[1-3]。这是因为快速生长的生物需要众多的核糖体,而生物体内磷的主要仓库恰好是核糖体,于是核糖体的大量增加就必然导致P的大量增加[2,4-5]。生长速率假说在浮游动物、节肢动物、细菌的研究中得到了广泛的验证,然而在陆生植物的研究中并没有得出一致的结论[6]。目前,对植物生长率假说的研究多集中在地上部分,对于地下部分的研究较少[7]。
异速生长是指生物体某一特征的相对生长速率不等于第二种特征的相对生长速率的特性,是生物体生长的一种特性,由遗传物质所决定,同时也受外界条件的影响[8]。异速生长关系简单来说就是不成比例的生长关系,通常以幂函数形式表示[6]。研究表明,植物体内C、N、P元素含量存在显著的相关生长关系,N随C呈等速增长,P随C呈3/4幂指数增长[6]。不同物种或相同物种的不同发育阶段对元素的吸收、运输、分配和利用过程存在显著差异,吉林西部退化草地C、N、P元素含量之间的异速生长关系需要进一步研究。
草地是我国最重要的生态系统类型之一。吉林省西部草地的优势种是羊草,它生态适应性广,可塑性强,对干旱和盐碱化环境具有较强的耐受性,被认为是该区草地恢复和重建最有前景的牧草之一[9]。对于退化草地来说,羊草的生长高度和覆盖程度随着退化程度的加剧而降低[10]。C∶N∶P是分析生态系统的有力工具[4]。C∶N∶P化学元素比率可以影响种群稳定性和群落结构[11-12],研究 C:N:P的比值与羊草生长率的关系,不仅具有重要的理论意义,还将为退化草地的治理给出有益的启示。笔者对吉林西部退化草地不同退化演替阶段羊草的生长率假说进行验证,并对根、茎、叶中C、N、P元素含量之间的异速生长关系进行研究。
1 材料与方法
1.1 研究区概况 吉林西部位于松嫩平原西南,包括松原市与白城市2个地级市,10个县(市、区)。地理坐标123°09'~124°22'E,44°57'~45°46'N,总面积约 47 000 km2。该区为典型的半湿润半干旱季风气候,四季分明,春季多风干燥,夏季高温多雨,秋季晴朗多日照,冬季寒冷漫长。多年平均降水量约400 mm,集中于7、8月份,平均蒸发量远大于降水量。研究区多年平均气温约4.5℃,7月气温最高,为23℃,1月气温最低,为-17.5℃。该区主要河流有第二松花江、嫩江、霍林河和洮儿河等,松花江和嫩江位于该区边界,且海拔较低,不易利用,洮儿河和霍林河近年来水量显著减少,全区地表水缺乏。该区主要地貌类型为平坦的河流低阶地、倾斜的河流高阶地、河漫滩、湖积冲积平原、沙丘等。土壤类型有淡黑钙土、草甸土、风沙土、黑钙土、碱土和栗钙土等。在地势较低的地区往往分布着盐碱湖泊,湖泊周围为盐碱土或盐碱化的草甸土。
1.2 样品采集与分析 根据土壤的退化程度和羊草群落的不同密度,采用以空间序列代替草地退化时间序列的方法进行样方调查,样方大小为100 cm×100 cm,样方基本上处于同一海拔高度,地势平坦,且土壤类型一致。分别于2010年6月和2012年6月在吉林省西部大安市、长岭县和前郭县选取具有代表性的采样点,共采集羊草样品73件,包括根28件、茎17件、叶28件。植物样品采集时,将羊草植株全部取出,尽量减少根系损失量,拣出植物的枯死体,采用四分法取样,装入牛皮纸信封保存并标记。采回的新鲜羊草样品带回实验室后,须立即将根、茎、叶3部分剪开,以免养分运转,用蒸馏水冲洗后,用吸水纸吸干;在105℃鼓风干燥箱中杀青15 min,降温至65℃烘干12 h,使水分去除干净,利用电子天平称量样品干重。
1.3 测定项目与方法 植物样品经60℃烘干,用粉碎机粉碎后保存于干燥器内,用于测定羊草C、N、P含量。植物全碳测定采用高温催化氧化消解法;全氮采用H2SO4-H2O2消煮,蒸馏法测定;全磷采用H2SO4-H2O2消煮,钼锑抗比色法测定[13]。
1.4 数据处理与分析 试验数据用Excel 2010统计软件进行整理。采用简约主轴回归(reduced major axis regression,RMA)(即第II类回归模型),进行生长率假说验证和元素间异速生长关系分析。RMA回归采用软件Standardized Major Axis Tests and Routines(SMATR)[14]进行计算。原始数据经对数变换以符合正态分布。简约主轴回归,又叫做几何平均数回归,与最小二乘法回归(ordinaryleast squares regression,OLS)(即第Ⅰ类回归模型)最大的不同,是考虑到解释变量也是存在随机误差的。因此,对于随机变量来说,这种相对严谨的分析方法能得到更科学的结果。普通最小二乘回归多用于预测2个变量之间的变化趋势,当需要了解2个变量之间的功能关系时,RMA回归是最合适的方法,而且该方法也是异速生长分析的标准方法[15]。
2 结果与分析
2.1 生长率假说的验证 植物生长所需要的化学元素主要来自于土壤。植物各个器官对植物生长的影响是有区别的,根系是吸收作用,茎是运输作用,叶片是光合作用。植物生长时,各个器官的作用不同导致C、N、P元素含量在植物体内分布不同[16]。生长率假说是研究生物生长率与化学计量特征之间关系的理论[17]。引入生长率公式计算羊草绝对生长率,分析其与C、N、P元素比值之间的联系,研究退化草地在N、P缺乏的情况下是否符合生长率假说。绝对生长率方程为 μ=Ln(Mt/M0)/t[5],式中 Mt和M0为前后两个时间的单位面积样方中羊草生物量干重;t为时间,d。这里,Mt为2012年8月3日的羊草植株、地上或地下的生物量;M0为2012年7月2日羊草植株、地上或地下的生物量。结果表明,N、P元素限制下,地下部分生长率与根系C∶P之间存在显著正相关关系,回归斜率αRMA=0.000 175 4,显著水平P=0.025,决定系数R2=0.854(表1);地上部分生长率与茎N∶P之间存在显著正相关关系,回归斜率αRMA=0.023 05,显著水平P=0.038,决定系数R2=0.809(表2)。由表1~3可知,羊草地下部分生长率与根系C∶N、N∶P无明显的相关关系,羊草地上部分生长率与茎C∶N、C∶P无明显的相关关系,羊草地上部分生长率与叶C∶N、C∶P、N∶P均无明显的相关关系,不符合生长速率假说。

表1 羊草地下部分生长率与根系C∶N∶P比值的关系
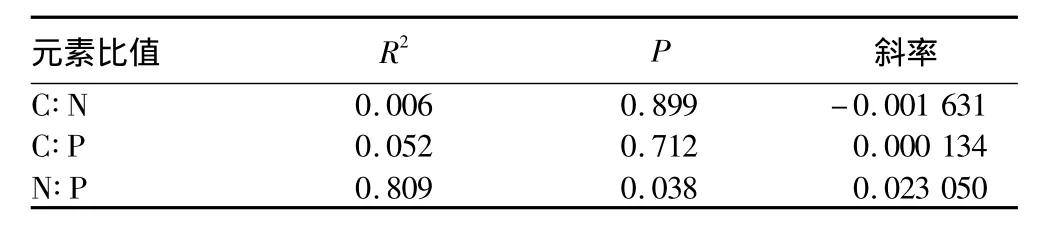
表2 羊草地上部分生长率与茎C∶N∶P比值的关系

表3 羊草地上部分生长率与叶C∶N∶P比值的关系
针对吉林西部的退化草地而言,N、P营养元素缺乏,羊草地上部分生长率与茎N∶P之间存在正相关关系,这与庾强[7]对内蒙古草原维管植物物种内生长率的研究一致,但与生长速率理论不符。庾强对内蒙古草原维管植物物种内生长率研究表明,内蒙古草原维管植物物种内生长率与N∶C和P∶C呈正相关,符合生长率假说[7]。磷限制条件下,生长率与N∶P呈负相关,也符合生长率假说。但是,氮限制条件下生长率与N∶P呈正相关,与生长率假说不符[6]。这可能是由于生长率假说是在无营养元素限制条件下针对浮游生物和细菌提出的,而该研究是针对吉林西部的退化草地而言,N、P营养元素缺乏,并且羊草植物属于自养生物。生长率假说对光合自养生物并不是完全一致的,主要因为光合自养生物尤其是植物存在储藏养分的器官,掩盖了C∶N∶P化学计量特征与生长速率的关系[5,18]。植物的生长率与N∶P的关系目前尚未明确,因此关于高等植物生长率的调控机制尚需进一步研究。
2.2 羊草根、茎、叶中C、N、P的异速生长关系 异速生长是指生物体各部分器官不均匀和不成比例地生长,用于研究生长中的整体与部分或部分与部分之间的关系[8]。异速生长方程表示为:y=βxα,式中x、y是生物属性,如羊草各器官中C、N、P含量及其比率;β为异速生长常数,α为异速生长指数[19]。将上式进行以10为底的对数转换,得到如下方程logy=αlogx+logβ。然后,用简化主轴回归方法(reduced major axis,RMA)计算异速生长关系的指数αRMA和常数logβRMA。由表4可知,羊草茎中N、P呈显著的异速生长关系(αRMA=0.675 8,P=0.005),羊草叶片中C、P以及N、P呈显著的异速生长关系(C、P:αRMA=0.242 4,P=0.005;N、P:αRMA=0.832 8,P=0.004)。其余器官C、N、P之间没有显著的相关关系。该研究结果与Niklas等得出的异速生长关系并不一致[20]。因此,吉林西部退化草地中羊草的生长不符合3/4法则。
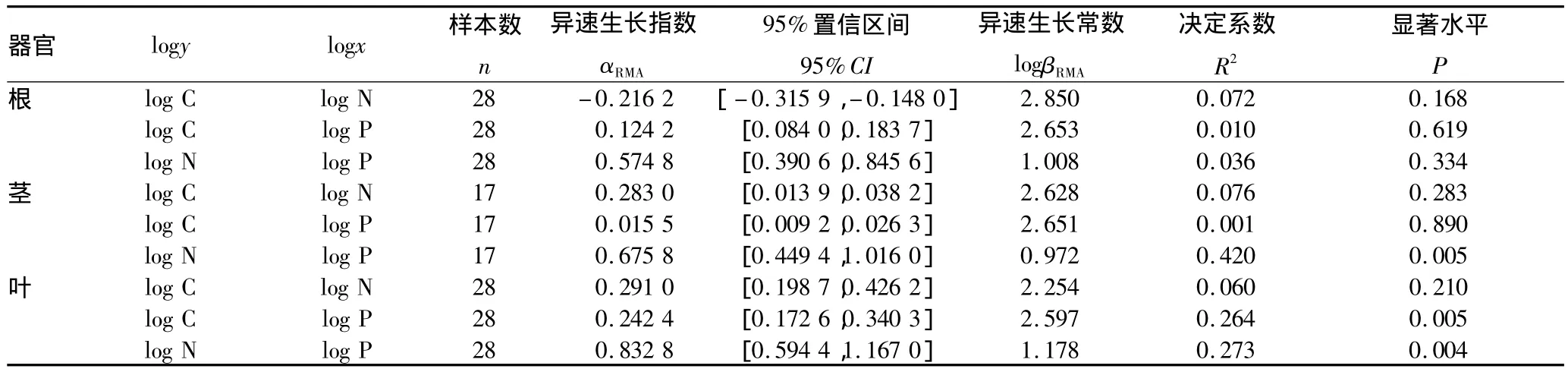
表4 羊草根、茎、叶、中C、N、P之间的异速生长关系
前人在研究C、N、P的异速生长关系时,多是研究不同物种的异速生长关系。比如,Niklas等通过搜集131种禾草的元素数据,分析得出叶片中N含量与C含量呈显著的等速生长,叶片中N含量与P含量呈幂指数为3/4的异速生长[20]。这里研究了单个物种的异速生长关系,因此异速生长规律在物种水平的应用需要进一步研究。
3 结论
(1)随着吉林西部草地退化演替的进行,在N、P元素限制下,羊草地下部分生长率与根系C∶P之间存在显著正相关关系,地上部分生长率与茎N∶P之间存在显著正相关关系。羊草地下部分生长率与根系根C∶N,N∶P无明显的相关关系,羊草地上部分生长率与茎C∶N、C∶P无明显的相关关系,羊草地上部分生长率与叶C∶N、C∶P、N∶P均无明显的相关关系,不符合生长速率假说。
(2)羊草茎中的N、P呈显著的异速生长关系,羊草叶片中C、P以及N、P呈显著的异速生长关系,其余器官C、N、P之间没有显著的相关关系。吉林西部退化草地中羊草的生长不符合3/4法则。其他植物是否符合生长率假说和异速生长关系还需要进一步研究。
(3)研究了单个物种的生长率假说和异速生长关系,为进一步研究不同物种、群落或生态系统的化学计量特征提供了参考。
[1]HESSEN D O,LYCHE A.Inter-and intraspecific variations in zooplankton element composition[J].Archiv für Hydrobiologie,1991,121(3):343-353.
[2]ELSER J J,DOBBERFUHL D R,MACKAY N A,et al.Organism size,life history,and N∶P stoichiometry[J].Bio Science,1996,46(9):674-684.
[3]STERNER R W,SCHULZ K L.Zooplankton nutrition:Recent progress and a reality check[J].Aquatic Ecology,1998,32(4):261-279.
[4]ELSER J J,FAGAN W F,DENNO R F,et al.Nutritional constraints in terrestrial and freshwater food webs[J].Nature,2000,408(6812):578-580.
[5]STERNER R W,ELSER J J.Ecological stoichiometry:The biology of elements from moleculaes to the biosphere[M].Princeton:Princeton University Press,2002.
[6]严正兵,金南瑛,韩廷申,等.氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响[J].植物生态学报,2013,37(6):551-557.
[7]庾强.内蒙古草原植物化学计量生态学研究[D].北京:中国科学院植物研究所,2009
[8]刘秀香,杨允菲.松嫩平原不同生境芦苇生殖分株的异速生长分析[J].草业学报,2012,21(4):313-318.
[9]李晓宇,蔺吉祥,李秀军,等.羊草苗期对盐碱胁迫的生长适应及Na+,K+代谢响应[J].草业学报,2013,22(1):201.
[10]桂花.呼伦贝尔草原不同程度退化下羊草群落特征[J].南方农业学报,2012,43(12):2035-2039.
[11]STERNER R W,HESSEN D O.Algal nutrient limitation and the nutrition of aquatic herbivores[J].Annu Rev Ecol Syst,1994,25:1-29.
[12]ANDERSEN T.Pelagic nutrient cycles:Herbivores as sources and sinks[M].Berlin:Springer,1997.
[13]李月芬,王冬艳,杨小琳,等.基于土壤化学性质与神经网络的羊草碳氮磷含量预测[J].农业工程学报,2014,30(3):104-111.
[14]FALSTER D S,WARTON D I,WRIGHT I J.(S)MATR:Standardised major axis tests and routines(Version2.0)[EB/OL].[2015-05-06].http://www.Bio.mq.edu.au/ecology/SMATR.
[15]张元明.准噶尔荒漠6种类短命植物生物量分配与异速生长关系[J].草业学报,2014,23(2):38-48.
[16]印婧婧,郭大立,何思源,等.内蒙古半干旱区树木非结构性碳,氮,磷的分配格局[J].北京大学学报:自然科学版,2009(3):519-527.
[17]徐劲草,许新宜.羊草生长率的研究和生长率假说的验证[J].草业科学,2013,30(1):74-79.
[18]ÅGREN G I.Stoichiometry and nutrition of plant growth in natural communities[J].Annual Review of Ecology,Evolution,and Systematics,2008,39:153-170.
[19]NIKLAS K J.Plant allometry:The scaling of form and process[M].Chicago:University of Chicago Press,1994.7 –34.
[20]NIKLAS K J,COBB E D.N,P and C stoichiometry of Eranthis hyemalis(Ranunculaceae)and the allometry of plant growth[J].American Journal of Botany,2005,92(8):1256-1263.
猜你喜欢
现代畜牧科技(2021年7期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
幼儿100(2020年31期)2020-11-18
防护林科技(2020年9期)2020-11-09
疯狂英语·初中版(2019年4期)2019-09-10
中南林业调查规划(2018年3期)2018-11-24
小太阳画报(2018年6期)2018-05-14
现代畜牧科技(2016年9期)2016-02-21
中南林业调查规划(2015年3期)2015-12-20
生态学报(2014年8期)2014-05-05
