
几种生态因子对成体偏顶蛤存活的影响
2015-02-17 08:40宁军号张向向常亚青宋坚经晨晨张伟杰杨立猛
大连海洋大学学报 2015年3期
宁军号,张向向,常亚青,宋坚,经晨晨,张伟杰,杨立猛
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023)
偏顶蛤 Modiolus modiolus(Linnaéus,1758)隶属于软体动物门、瓣鳃纲、贻贝目、壳菜蛤科、偏顶蛤属,俗称毛海红、假海红,主要分布于北半球,在中国主要分布于黄、渤海海域。偏顶蛤个体较大、肉质鲜美、营养丰富,是一种具有较好养殖开发前景的贝类[2]。目前,国内外有关环境因子对贝类影响的研究很多,主要集中在环境因子对贝类能量收支的影响[2],对幼虫、稚贝生长率和存活率的影响[3-8],以及对贝类耐干露和耐淡水浸泡能力的影响[9-11]。迄今尚未见有关环境因子对偏顶蛤存活影响的相关报道。本试验中,研究了海水盐度、温度、pH、干露和淡水浸泡对偏顶蛤存活的影响,旨在为偏顶蛤的增养殖和自然资源保护提供基础资料。
1 材料与方法
1.1 材料
试验用偏顶蛤采自大连市黑石礁海域,运回农业部北方海水增养殖重点实验室暂养1周,挑选健康个体进行耐受性试验。偏顶蛤壳长为 (99.1±6.1)mm,体质量为 (108.8 ± 18.9)g。暂养期间,每天8:00换水1次,换水后投饵,饵料为破壁酵母粉、螺旋藻粉和破壁小球藻粉的混合人工饵料。试验用水为经三级沙滤的自然海水,温度为10~12℃,盐度为31.2,pH为8.05,连续充气。
1.2 方法
1.2.1 盐度试验 以经沙滤、沉淀处理的自然海水与高浓度海水晶和充分曝气的自来水,配制成0、5、10、15、20、25、30、35、40、45、50 共10个盐度组,每个盐度组设置2个平行,每个平行随机放20只偏顶蛤。盐度检测使用 YSI 12F100016型水质仪。试验过程中,海水温度控制在10℃,pH稳定在8.05,连续充气,每天8:00换水1次,换水后温度波动不超过1℃。换水后投饵 (饵料同暂养期间),每次投饵后调整盐度,盐度波动不超过0.5。每天9:00和20:00分别观察和记录偏顶蛤的存活、摄食和排遗情况。
1.2.2 温度试验 试验设 2、5、10、15、21、23、25、26、27、29、30℃共11个温度组,每个温度组设置2个平行。21℃以上温度组以加热棒维持水温,换水时以加热海水和自然海水调节水温;15℃以下温度组用高低温实验箱控制,换水时以冰袋调节海水水温,温度波动不超过0.5℃,其他试验条件同盐度试验。
1.2.3 pH 试验 用 1 mol/L的 HCl溶液和 1 mol/L的NaOH溶液配制成1、2、3、4、5、6、7、8、9、10、11共11个pH组,每个pH组设置2个平行。每次投饵后调整pH,pH波动不超过0.5。自然海水盐度为31.2,其他条件同盐度试验。
1.2.4 干露试验 将偏顶蛤置于干燥水槽底部,保持黑暗环境。用湿度计TH100-16测得空气湿度为85% ~90%。试验设置15、20、25℃共3个气温组,每组设置3个平行,每个槽中放20只贝。气温用空调维持,温度波动不超过0.5℃。每12 h观察和记录偏顶蛤状态及存活情况,偏顶蛤贝壳张开,刺激不能闭合,放入海水中5 h也不能闭合即为死亡。
1.2.5 淡水浸泡试验 试验用淡水为自来水,水温为10~12℃。试验设置投饵和不投饵两组,每组设置3个平行,每个平行放20只贝。每12 h观察和记录偏顶蛤状态及存活情况,判定死亡标准同干露试验。存活率 (R)的计算公式如下:
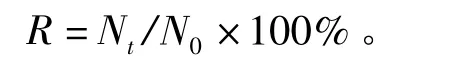
其中:Nt、N0分别为试验t时偏顶蛤的存活个数和试验开始时偏顶蛤的总数。
1.3 数据处理
用Excel 2003软件对试验数据进行初步整理,再用SPSS 17.0软件进行统计分析。将偏顶蛤存活率 (百分数)均进行反正弦 (arsin)转换,对温度、盐度和pH等转换后的试验数据做卡方精确性检验;干露和淡水浸泡试验的试验数据转换后,先经Kolmogorov-Smirnov正态性检验,再用重复度量方差分析对不同试验组偏顶蛤存活率差异进行比较,显著性水平设为0.05。
2 结果与分析
2.1 盐度对成体偏顶蛤存活的影响
从表1和试验观察情况可知:在海水温度为10~12℃、pH为8.05的条件下,淡水组 (对照组)和盐度5组偏顶蛤在第1天内均能100%存活,6 d后成活率不足10%,且两组分别在第7天和第8天时全部死亡;盐度10组和盐度15组偏顶蛤在2 d内能够100%存活,2 d后盐度10组偏顶蛤存活率逐渐下降,到第10天存活率仅为2.5%,盐度15组偏顶蛤在第7天还有超过半数存活,试验结束时 (第10天)仍有20%存活;偏顶蛤在盐度为20~40时均能存活,但观察发现,盐度20组和盐度40组偏顶蛤摄食情况较差,有较多残饵和假粪;盐度45组偏顶蛤虽有死亡,但第5天时还有90.0%存活,第10天存活率达70.0%;盐度50组偏顶蛤在2 d内存活率较高,达95.0%,但在第10天时全部死亡。此外还发现,盐度0~15组及盐度45~50组的偏顶蛤几乎不摄食,也未发现排泄物。

表1 不同盐度对成体偏顶蛤存活率的影响Tab.1 Effects of different salinities on survival rate of adult mussel Modiolus modiolus
2.2 温度对成体偏顶蛤存活的影响
从表2和试验观察情况可知:在海水盐度为31.2、pH为8.05的条件下,温度为2~23℃时,偏顶蛤能够正常生存、摄食和排遗,试验过程中存活率均为100%,但2℃组偏顶蛤有假粪现象;25℃组偏顶蛤在9 d内没有死亡,第10天时有个别死亡,存活率仍高达92.5%,且能够摄食和排遗;26℃组偏顶蛤在5 d内没有死亡,第10天时存活率降为67.5%,偏顶蛤虽能摄食、排遗,但有假粪存在;27℃组偏顶蛤在第1天就有30%以上死亡,偏顶蛤几乎不摄食,到第3天时存活率仅为25.0%,第4天时已全部死亡;温度为29℃和30℃的试验组,偏顶蛤在12 h内全部贝壳大张,刺激不能闭合,软体部颜色变深,海水浑浊,放入海水中5 h也无生命迹象。

表2 不同温度对成体偏顶蛤存活率的影响Tab.2 Effects of different temperature on survival rate of adult mussel Modiolus modiolus
2.3 pH对成体偏顶蛤存活的影响
从表3和试验观察情况可知:在海水盐度为31.2、温度为10~12℃的条件下,pH为1~3时,偏顶蛤在第1天内存活率为100%,3 d内成活率均在70%之上,pH为1、2、3时偏顶蛤分别在第6天、第8天、第8天时全部死亡;pH为4时偏顶蛤在3 d内没有发现死亡个体,到第5天存活率为92.5%,试验结束时,其存活率高达80.0%;pH为5~9时,各试验组均能够正常生存、摄食和排遗,但pH为5时有假粪出现;pH为10时在5 d内没有死亡个体,第7天时存活率高达95.0%,试验结束时,其存活率仍能达到80.0%;pH为11时在第1天内出现死亡个体,第5天时存活率在50%之上,到第8天时已全部死亡。
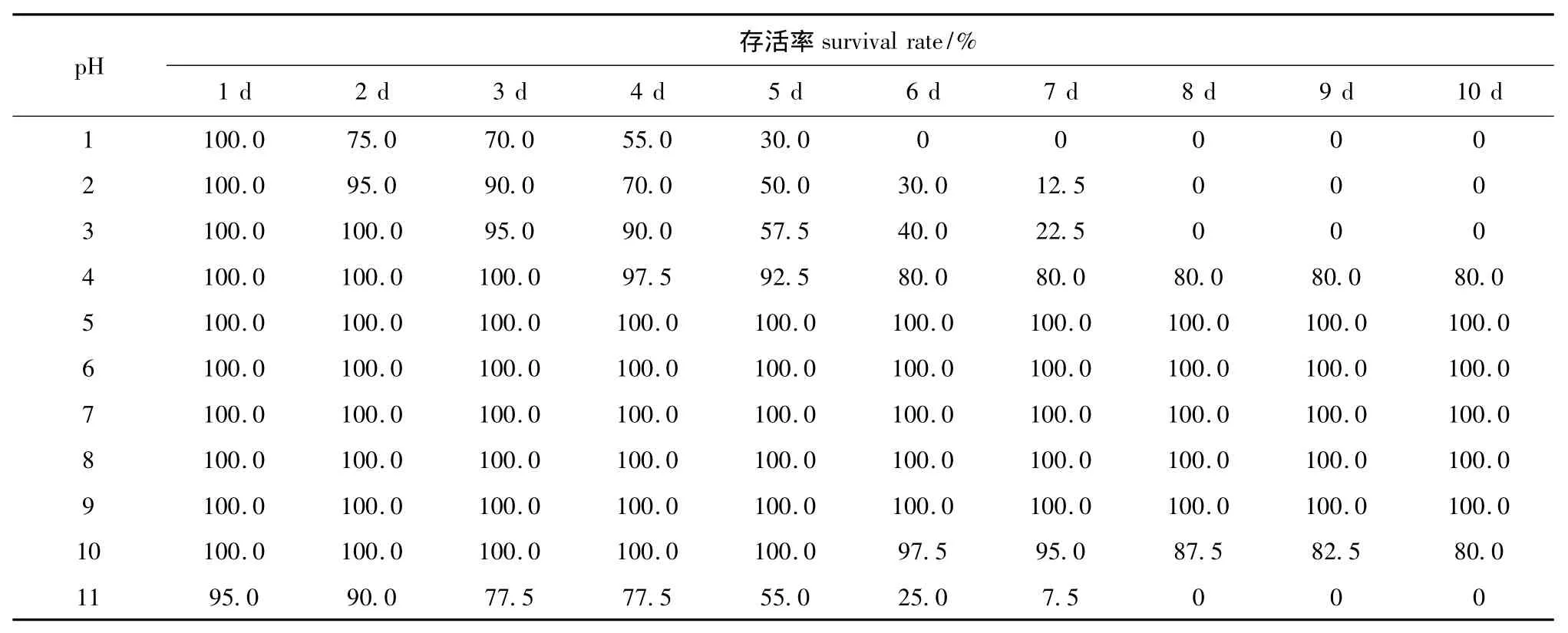
表3 不同pH对成体偏顶蛤存活率的影响Tab.3 Effects of different pH on survival rate of adult mussel Modiolus modiolus
2.4 干露对成体偏顶蛤存活的影响
从图1可见:在湿度为85%~90%的黑暗条件下,随着干露时间的延长,偏顶蛤的存活率呈显著下降趋势 (P<0.05),其中低温条件下的存活率明显高于高温条件下的;25℃组偏顶蛤存活率在48 h出现显著降低 (P<0.05),20℃组在60 h时偏顶蛤存活率出现显著下降 (P<0.05),而15℃组偏顶蛤直到84 h时存活率才出现显著下降(P<0.05),且存活率高达80%;15℃组偏顶蛤存活率在60 h后显著高于其他2组;25℃组偏顶蛤存活率在48 h后显著低于其他2组 (P<0.05)。15、20、25℃组偏顶蛤全部死亡时间分别为144、108、72 h。

图1 干露时间对成体偏顶蛤存活率的影响Fig.1 Effects of air exposure time on survival rate of adult mussle Modiolus modiolus
2.5 淡水浸泡对成体偏顶蛤存活的影响
从图2可见:在水温为10~12℃的黑暗条件下,随着淡水浸泡时间的延长,偏顶蛤存活率呈下降趋势;不投饵组和投饵组偏顶蛤100%存活的最长耐淡水浸泡时间虽均为24 h,但在前3 d内,投饵组出现大量死亡,第3天时存活率仅为41.7%;不投饵组在前3 d内偏顶蛤存活率均在85%之上,第4天时才明显降低 (P<0.05);3 d后,不投饵组偏顶蛤存活率显著高于投饵组 (P<0.05),但两组偏顶蛤均能存活较长时间,分别在第8天和第7天时才全部死亡。

图2 淡水浸泡时间对成体偏顶蛤存活率的影响Fig.2 Effects of fresh-water immersion time on survival rate of adult mussle Modiolus modiolus
3 讨论
3.1 成体偏顶蛤对盐度的耐受性
海水盐度变化可引起贝类体内渗透压改变,在适宜盐度范围内贝类可通过自身调节来适应,但超过自身调节能力时,贝类的代谢速率和代谢效率均受到抑制,更甚者可导致死亡[3,6,8]。本研究中,成体偏顶蛤在盐度为0~15和超过45时均不能正常摄食、排遗和生存,说明盐度超过了其自身调节范围。偏顶蛤能够存活的盐度范围为20~45,适宜盐度范围为25~35,这说明偏顶蛤对盐度的适应范围较广,这与对厚壳贻贝Mytilus coruscus Gould[12]青 蛤 Cyclina sinensis Gmelin[7]、硬 壳 蛤Mercenaria mercenaria[13]等广盐性贝类的研究结果相一致。本试验中发现,在第10天时,盐度10组和盐度50组偏顶蛤的存活率均接近0,推测已是其耐盐能力的下限和上限。贝类的耐盐能力对于其增养殖业具有重要意义,因为海产贝类养殖场和育苗场常分布在沿海、海湾和河口区,海水盐度易受气候和地表径流影响,短时间内会发生较大变化。本研究结果可为偏顶蛤养殖场、育苗场选址提供理论支撑。
3.2 成体偏顶蛤对温度的耐受性
温度是影响海产贝类生长、发育、繁殖和分布的重要因子之一,许多学者对其进行了相关研究[6-7,14-15]。本研究结果表明,成体偏顶蛤能够存活的温度范围为2~26℃,适宜温度范围为5~23℃,与成体小荚蛏 Siliquaminima Gmelin[16]适温范围相似,但比成体橄榄蚶Estellarca olivacea[17]的适温范围窄。水温与偏顶蛤新陈代谢活力密切相关,在适宜水温环境中偏顶蛤代谢旺盛,排放大量粪便;在低温或高温环境中,偏顶蛤代谢异常,很少有代谢物并伴有大量假粪;超出其本身温度适应能力时,会导致其大量死亡。27℃组偏顶蛤第1天死亡率高于30%,第4天就全部死亡,说明已是其能够承受的温度上限,至于其所能承受温度下限的详细数据还有待进一步研究。偏顶蛤自然分布于北半球,在中国主要分布在黄、渤海海域,营底栖固着生活[2],故其对低温的耐受能力要强于对高温的耐受能力,这也是物种对环境长期适应的结果。
3.3 成体偏顶蛤对pH的耐受性
自然海水pH一般变化不大,能够维持较稳定的弱碱性。然而近年来,随着CO2的过度排放与溶解,海洋酸化[18]日趋严重,近岸生物 (尤其含钙质外壳的海洋生物)的生存受到了严重威胁[19-20]。本研究结果表明,成体偏顶蛤能够生存的pH范围为4~10,适宜pH范围为6~9,与厚壳贻贝[12]、彩虹明樱蛤 Moerella iridescens[6]和墨西哥湾扇贝 Argopecten irradians concentricus[21]等贝类对pH耐受范围一致。本研究中发现,成体偏顶蛤对极端pH(pH为1、2、10、11)有一定的耐受性。因此,偏顶蛤亲贝催产前,可利用极端酸、碱环境在短时间内杀死偏顶蛤壳表的细菌、原生动物和寄生虫等致病微生物。从表3可知,pH为3和11时偏顶蛤均在第8天时全部死亡,推测海水pH为3和11时为成体偏顶蛤耐受pH的下限和上限。
3.4 成体偏顶蛤对干露的耐受性
贝类对干露的耐受能力与其种类、大小有紧密联系,并且与干露时长和高温呈负相关[17]。本研究表明,随着温度的升高,偏顶蛤存活率明显下降,耐干露时间显著缩短;随着干露时间的延长,其存活率也显著降低,试验结果印证了上述观点。这种结果是因为贝类干露一段时间后,贝壳张开,体内水分蒸发严重,软体部萎缩,不能获得足够O2和排除多余CO2,使得血液中pH发生不可逆变化,从而导致死亡[9-11]。在气温为15℃和20℃的条件下,成体偏顶蛤开始出现死亡的时间均为48 h,且气温在15℃下60 h时偏顶蛤还有95%的存活率,表明偏顶蛤有较强的露空能力。这是因为偏顶蛤软体部肥厚,双壳具有强大的闭合能力,可有效防止水分大量蒸发,与太平洋牡蛎Crassostrea gigas[9]、硬壳蛤[10]、中国蛤蜊 Mactra chinensis、四角蛤蜊 Mactra veneriformis[11]等贝类具有相似特点。因此,建议偏顶蛤在露空储运或销售等过程中,温度应保持在20℃以下,储运时间不要超过3 d。储运过程中偏顶蛤相互挤压,中下层偏顶蛤贝壳不易张开,水分蒸发减少,再定时喷洒海水维持较高湿度,可延长其耐干露时间。
3.5 成体偏顶蛤对淡水浸泡的耐受性
适当时间的淡水浸泡也可清除贝类壳表、附着基上的敌害生物,可有效预防贝类培育过程中敌害生物暴发引起的稚贝大批死亡现象[10,22]。本研究结果表明,淡水浸泡的前期 (前3 d),投饵组成体偏顶蛤会出现大量死亡,存活率显著低于不投饵组 (P<0.05),这可能与食物颗粒诱导偏顶蛤加快开壳探测外界环境的频率,淡水进入贝壳内引起其体内渗透压改变超过了自身调节能力,最终加速了偏顶蛤的死亡。但3 d后,存活下来的个体都可坚持7~8 d。这可能是偏顶蛤经受长达3 d的淡水浸泡胁迫以后,开壳速率放缓,对淡水浸泡的适应能力有所增强。偏顶蛤一经淡水浸泡等胁迫后,双壳闭合将水分留在外套膜中,主要依赖体内残留的氧气维持生存,直到7~8 d时达到其死亡极限。因此,贝类养殖场一旦注入大量淡水,除及时换水外,还要注意养殖水体在低盐期间应禁止投喂。偏顶蛤在淡水中浸泡24 h能100%存活,不投饵组3 d内存活率还高于85%,比中国蛤蜊和四角蛤蜊耐淡水浸泡能力强[11]。因此,对偏顶蛤催产时,淡水浸泡既可作为诱导产卵的刺激条件,又可用于清除亲贝体表的病原菌和寄生虫等敌害生物。
[1]齐钟彦,马绣同,王祯瑞,等.黄渤海的软体动物[M].北京:农业出版社,1987:164-166.
[2]黄洋,黄海立,林国游,等.盐度、pH和规格对尖紫蛤(Soletellina acuta)耗氧率和排氨率的影响[J].海洋与湖沼,2013,44(1):120-125.
[3]Kinne O.The effects of temperature and salinity on marine and brackish water animals:Ⅱ.salinity and temperature-salinity combination[J].Oceanography Marine Biology Annual Review,1964,2:281-339.
[4]Yan X,Zhang G,Yang F.Effects of diet,stocking density,and environmental factors on growth,survival,and metamorphosis of Manila clam Ruditapes philippinarum larvae[J].Aquaculture,2006,253(1):350-358.
[5]De Bravo M S,Chung K S,Pérez J E.Salinity and temperature tolerances of the green and brown mussels,Perna viridis and Perna perna(Bivalvia:Mytilidae)[J].Revista De Biologia Tropical,1998,46(5):121-125.
[6]顾晓英,尤仲杰,王一农.几种环境因子对彩虹明樱蛤 Moerella iridescens不同发育阶段的影响[J].东海海洋,1998,16(3):40-47.
[7]王丹丽,徐善良,尤仲杰,等.温度和盐度对青蛤孵化及幼虫、稚贝存活与生长变态的影响[J].水生生物学报,2006,29(5):495-501.
[8]何义朝,张福绥.盐度对海湾扇贝不同发育阶段的影响[J].海洋与湖沼,1990,21(3):197-204.
[9]于瑞海,王昭萍,孔令锋,等.不同发育期的太平洋牡蛎在不同干露状态下的成活率研究[J].中国海洋大学学报:自然科学版,2006,36(4):617-620.
[10]李忠泓,王国栋.硬壳蛤稚贝对淡水浸泡、干露和低温的耐受能力[J].水产科学,2004,23(6):14-16.
[11]马贵范,杨凤,郭文学,等.中国蛤蜊和四角蛤蜊对干露和淡水浸泡的耐受性比较[J].大连海洋大学学报,2013,28(1):44-48.
[12]叶莹莹,徐梅英,吴常文.几种环境因子对厚壳贻贝浮游幼虫生长与存活的影响[J].浙江海洋学院学报:自然科学版,2011,30(4):292-296.
[13]常亚青,宋坚,王国栋,等.硬壳蛤的人工育苗技术[J].中国水产科学,2002,9(1):43-47.
[14]林笔水,吴天明.温度与盐度和缢蛏幼体生存、生长及发育的关系[J].水产学报,1990,14(3):171-178.
[15]尤仲杰,徐善良,边平江,等.海水温度和盐度对泥蚶幼虫和稚贝生长及存活的影响[J].海洋学报,2001,23(6):108-113.
[16]柴雪良,张永普,宣瑛,等.若干环境因子对成体小荚蛏存活的影响[J].海洋学研究,2008,26(1):45-51.
[17]周化斌,张永普,肖国强,等.几种环境因子对橄榄蚶成贝存活的影响[J].温州大学学报:自然科学版,2010,31(2):30-35.
[18]Caldeira K,Wickett M E.Oceanography:anthropogenic carbon and ocean pH[J].Nature,2003,425:365.
[19]湛垚垚,黄显雅,段立柱,等.海洋酸化对近岸海洋生物的影响[J].大连大学学报,2013,34(3):79-84.
[20]Hoegh-Guldberg O,Mumby P J,Hooten A J,et al.Coral reefs under rapid climate change and ocean acidification[J].Science,2007,318:1737-1742.
[21]尤仲杰,陆彤霞,马斌,等.几种环境因子对墨西哥湾扇贝幼虫和稚贝生长与存活的影响[J].热带海洋学报,2003,22(3):22-29.
[22]杨凤,谭文明,闫喜武,等.干露及淡水浸泡对菲律宾蛤仔稚贝生长和存活的影响[J].水产科学,2012,31(3):143-146.
猜你喜欢
今日农业(2021年1期)2021-03-19
渔业致富指南(2020年16期)2020-12-18
渔业致富指南(2020年5期)2020-12-18
理化检验-化学分册(2020年5期)2020-06-15
湖南畜牧兽医(2020年6期)2020-03-22
食品与生活(2019年8期)2019-10-30
新农村(浙江)(2018年5期)2018-05-31
益寿宝典(2018年11期)2018-01-27
食品与生活(2014年8期)2014-10-20
中学生物学(2008年12期)2008-12-27
