
海南野生豹纹鳃棘鲈染色体核型及银染研究
2015-02-17 08:40高宁蔡岩周永灿李根浪袁卫刘海天谢珍玉王世锋
大连海洋大学学报 2015年3期
高宁,蔡岩,周永灿,李根浪,袁卫,刘海天,谢珍玉,王世锋
(海南大学热带生物资源可持续利用省部共建国家重点实验室培育基地,海南省热带水生生物技术重点实验室,海南 海口570228)
豹纹鳃棘鲈Plectropomus leopardus俗称东星斑,隶属于鲈形目、鮨科、石斑鱼亚科、鳃棘鲈属,为暖水性岛礁鱼类,主要分布于西太平洋至印度洋海区,日本、澳洲、斐济等地也有少量分布。豹纹鳃棘鲈因体色艳丽、肉质鲜美、市场价值高,导致野生资源遭到过度捕捞,已被国际自然保护联盟(International Union for the Conservation of Nature,IUCN)列为濒危物种[1]。近年来,国内外学者对豹纹鳃棘鲈的生态学、分类学、胚胎发育、人工育苗及养殖技术等研究较多[2-5]。仅见 Pinthong等[6]对台湾海域采集的豹纹鳃棘鲈进行了核型及银染分析,但与从海南采集的样品在银染研究结果上存在差异。本研究中,对海南野生豹纹鳃棘鲈的染色体核型特征进行研究,旨在为石斑鱼类染色体进化分析等提供资料,同时也为其种群种质资源的开发与利用、遗传育种与系统发育关系的探讨奠定基础。
1 材料与方法
1.1 材料
试验用鱼为捕获于海南东岸海域的野生鱼(图1,全长为14.5 cm),共5尾,体质量为30~50 g,活体运至实验室充气暂养。
1.2 方法
1.2.1 染色体标本的制备 按照吉华松等[7]的方法并稍作修改。从样品鱼尾柄处以10 μL/g(鱼体质量)抽血,在胸鳍基部以10 μg/g(鱼体质量)注射植物血球凝集素 (PHA);3.5 h后于胸鳍基部以1 μg/g(鱼体质量)注射秋水仙素;30 min后在水中剪鳃放血,然后在解剖盘中取其头肾,于0.85%生理盐水中制备细胞悬液,静置片刻后吸取上层细胞悬液放入5 mL离心管中,以1000 r/min离心5 min,弃上清后轻轻弹散沉淀,缓慢加入低渗液,于37℃下水浴25~35 min;离心后缓慢加入适量新配制的卡诺氏固定液,室温固定20 min,之后重复固定2次;加入适量新配制的卡诺氏固定液,置于冰箱 (-20℃)中过夜;第二天用吸管小心将上清吸出后,打散沉淀,加适量新配制的固定液制成细胞悬液,用吸管吸取细胞悬液滴于干净的载玻片上,过酒精灯3次后自然晾干。用Giemsa染液染色15~20 min后,将玻片冲洗干净并自然晾干,在显微镜下观察并拍照。
1.2.2 核型分析 在显微镜下选取分散良好的中期分裂相进行染色体观察和计数,确定豹纹鳃棘鲈二倍体染色体数目。选取染色体细长、分散良好和形态清晰的中期分裂相进行拍照和测量,根据统计分析结果确定染色体的相对长度和臂比,根据Levan等[8]的标准确定染色体类型。

图1 豹纹鳃棘鲈Fig.1 Leopard coraltrout Plectropomus leopardus
1.2.3 Ag-NORs显带 参照 Howell等[9]的方法并略加修改。在培养皿底部放置一层滤纸,然后用蒸馏水润湿,在滤纸上平行放置2根牙签,置60℃水浴锅中待用;玻片标本细胞面朝上,将50%AgNO3溶液与2% 明胶溶液 (内含1%甲酸)以2∶1的比例滴加到染色体标本上,混匀后盖上盖玻片,然后平放在培养皿中的牙签上。染色体标本在水浴锅中温浴至标本呈棕黄色时取出,用自来水冲去盖玻片,干燥后在显微镜下观察并拍照。
2 结果与分析
2.1 豹纹鳃棘鲈体细胞染色体数目
本试验对182个染色体分散良好、形态清晰的中期分裂相进行观察和计数,二倍体染色体数目为48的中期分裂相最多,有138个,占75.82%(表1),由此可确定豹纹鳃棘鲈二倍体染色体众数为48,即 2n=48。

表1 豹纹鳃棘鲈染色体中期分裂相的统计结果Tab.1 Diploid chromosome number of leopard coraltrout Plectropomus leopardus during metaphase
2.2 染色体核型
选择6个分散良好、形态清晰和数目完整的中期分裂相进行拍照,经测量和统计分析确定染色体组中各条染色体相对长度和臂比,进而判定染色体类型,结果如图2和表2所示。由图2和表2可见,在豹纹鳃棘鲈染色体标本中,可观察到两种核型:A型染色体较短,由24对端部着丝点染色体(t)构成 (图2-A、B);B型染色体较为细长,由1对亚端部着丝粒染色体和23对端部着丝粒染色体构成 (图2-C、D),这可能是不同中期分裂相受秋水仙素作用时间不同所致,因此,豹纹鳃棘鲈的核型公式为2n=2st(t)+46t,NF=48。豹纹鳃棘鲈染色体组中,第1对染色体相对长度明显长于其他23对染色体,第24对明显短于其他染色体,而其他22对染色体相对长度呈逐渐递减的趋势,相邻染色体之间无明显的大小差别。
2.3 Ag-NORs带型
对豹纹鳃棘鲈61个二倍体染色体数目为48、银染位点清晰的中期分裂相进行统计分析,结果表明,在第1对染色体上具有1对NORs,位于染色体长臂靠近着丝粒的位置 (图3)。对112个间期细胞核中的核仁组织区数量进行统计,具有1个核仁组织区的间期细胞核有78个,占总数的70%;具有2个核仁组织区的有34个,占总数的30%。
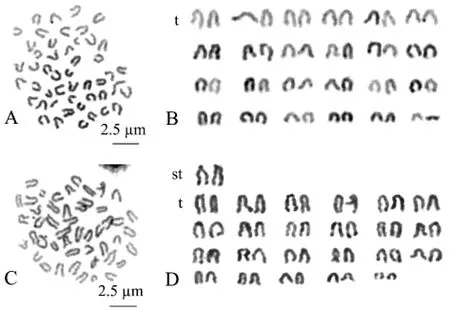
图2 豹纹鳃棘鲈染色体中期分裂相 (A、C)和核型(B、D)Fig.2 Chromosome metaphase(A,C)and karyotypes(B,D)in leopard coraltrout Plectropomus leopardus

图3 豹纹鳃棘鲈间期细胞核 (A)和中期分裂相的Ag-NORs(B)Fig.3 Nuclei at interphase and Ag-NORs in metaphase chromosomes in leopard coraltrout Plectropomus leopardus
3 讨论
海南豹纹鳃棘鲈二倍体染色体数目为48,核型公式为2n=2st(t)+46t,NF=48,与 Pinthong等[6]对台湾采集样品的研究结果基本一致。鳃棘鲈属作为石斑鱼亚科中较为原始的类群[10],其染色体组组成与石斑鱼属原始核型特征一致[11],可见在进化过程中,石斑鱼类染色体核型具有较高的保守性。这种核型的保守性,不仅仅存在于科这一分类阶元,2n=48t也被认为是鲈形目基本的原始核型特征[12]。
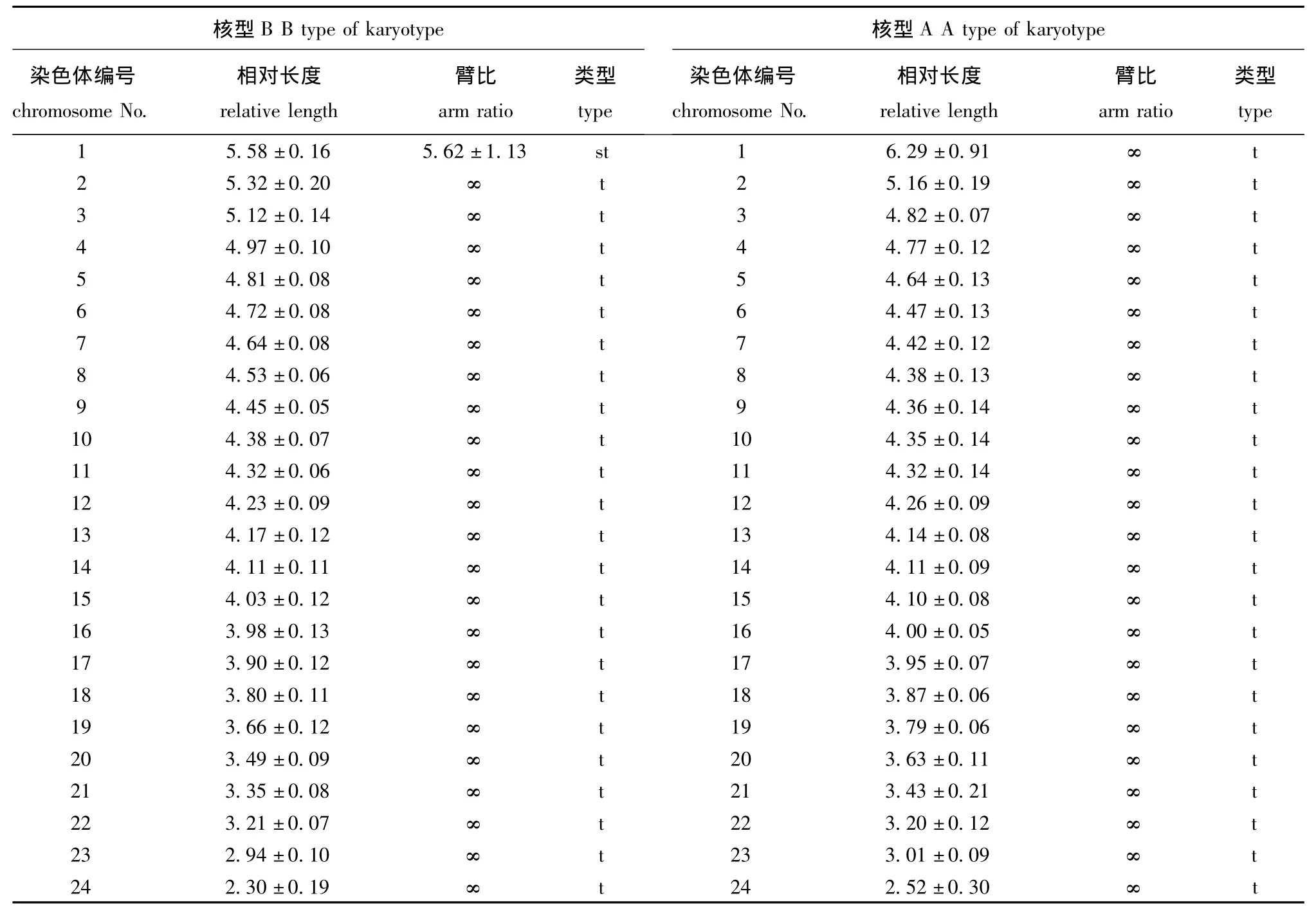
表2 豹纹鳃棘鲈染色体相对长度和臂比Tab.2 Relative length and arm ratio of chromosomes in leopard coraltrout Plectropomus leopardus
海南豹纹鳃棘鲈染色体组中,除第1对和第24对染色体与其他染色体在长度上稍有差异外,其他22对染色体相对长度呈逐渐递减的趋势,相邻染色体形态、大小差异不明显,容易造成同源染色体的错配。染色体显带技术可显著提高核型分析的准确率[13]。本研究中对海南豹纹鳃棘鲈的Ag-NORs研究发现,NORs位于第1对染色体长臂靠近着丝粒的位置,而Pinthong等[6]研究结果显示,台湾地区豹纹鳃棘鲈的NORs位于第20对染色体靠近着丝粒的位置,这可能是豹纹鳃棘鲈不同地理群体间NORs分布位置具有多态性[14]。
不同作者对同一鱼类进行核型分析,可能会由于采用的染色体制备方法不同、利用体细胞或胚胎细胞等材料不同、采集样品的地理区域不同等原因造成试验结果存在差异[15]。卓孝磊等[13]对中国研究过核型的海水鱼类进行了总结,在77种海水鱼类中,有8种海水鱼类的核型公式在不同的研究报道中存在差异,包括星康吉鳗、大黄鱼、斜带髭鲷、纹缟鰕虎鱼、许氏平鮋、斑头鱼、欧氏六线鱼、红鳍东方鲀等。在石斑鱼亚科中,鲑点石斑鱼[16-17]、云纹石斑鱼[18-19]、赤点石斑鱼[20-21]、三斑石斑鱼等[15,22]也存在上述情况。对上述报道进行比较分析,发现同一物种在不同报道中所显示的核型公式上的差异,多为染色体组中不同染色体类型在数量上的差异。如赤点石斑鱼,在王云新等[20]的研究报道中染色体组由10条亚端部着丝粒染色体 (st)和38条端部着丝粒染色体构成,而在王世锋[21]的研究中则由2条亚中部着丝粒染色体 (sm)、8条亚端部着丝粒体 (st)和38条端部着丝粒染色体构成。本研究中发现,豹纹鳃棘鲈同一染色体标本的不同中期分裂相之间也存在上述现象,在染色体相对细长的中期分裂相中,第1对染色体具有短臂,为亚端部着丝染色体 (st),而在凝缩较为严重的中期分裂相中,第1对染色体为端部着丝粒染色体 (t)。这可能是由于豹纹鳃棘鲈的头肾细胞分裂不同步,秋水仙素对不同中期分裂相中的染色体作用时间不同,从而导致染色体凝缩程度不同,造成染色体形态上的差异[23]。因此,在不同报道中同一种海水鱼类在核型上的差异,是否同样是由于秋水仙素作用时间不同,以及测量时所选取的中期分裂相染色体凝缩程度不同造成的,还有待进一步研究。
[1]Zgliczynski B J,Williams I D,Schroeder R E,et al.The IUCN red list of threatened species:an assessment of coral reef fishes in the US Pacific islands[J].Coral Reefs,2013,32:627-650.
[2]Sun Z,Xia S,Feng S,et al.Effects of water temperature on survival,growth,digestive enzyme activities,and body composition of the leopard coral grouper Plectropomus leopardus[J].Fish Sci,2015,81:107-112.
[3]Cai X,Qu M,Ding S,et al.Differentiation of coral trout(Plectropomus leopardus)based on an analysis of morphology and complete mitochondrial DNA:are cryptic species present?[J].Acta Oceanol Sin,2013,32(6):40-46.
[4]陈超,吴雷明,李炎璐,等.豹纹鳃棘鲈(Plectropomus leopardus)早期形态与色素变化及添加剂对其体色的影响[J].渔业科学进展,2014,35(5):83-90.
[5]刘天密,王永波,符书源,等.豹纹鳃棘鲈室外大型水泥池的人工育苗技术[J].海洋渔业,2012,34(4):400-405.
[6]Pinthong K,Gomontean B,Kongim B,et al.Cytogenetic comparison and chromosome localization of the nucleolar organizer region of four grouper genera(Pisces,Epinephelinae)from Thailand[J].Cytologia,2013,78:223-234.
[7]吉华松,周永灿,蔡岩,等.六带石斑鱼染色体核型和银染研究[J].水产科学,2011,30(8):465-466.
[8]Levan A,Fredga K,Sandberg A A.Nomenclature for centromeric position on chromosomes[J].Hereditas,1964,52(2):201-220.
[9]Howell M W,Black D A.Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer:a1-step method[J].Experientia,1980,36(8):1014-1015.
[10]丁少雄,王颖汇,王军,等.基于16S rDNA部分序列探讨中国近海30种石斑鱼类的分子系统进化关系[J].动物学报,2006,52(3):504-513.
[11]Wang S F,Su Y Q,Ding S X,et al.Cytogenetic analysis of orange-spotted grouper,Epinephelus coioides,using chromosome banding and fluorescence in situ hybridization[J].Hydrobiologia,2010,638(1):1-10.
[12]Galetti Jr P M,Molina W F,Affonso P R A M,et al.Assessing genetic diversity of Brazilian reef fishes by chromosomal and DNA markers[J].Genetica,2006,126:161-177.
[13]卓孝磊,邹记兴.我国海水鱼类核型及染色体显带研究进展[J].热带海洋学报,2007,26(5):73-80.
[14]任修海,余其兴,韦萍.黄鳝染色体Ag-NORs多态性的研究[J].遗传学报,1991,18(4):304-311.
[15]舒琥,魏秋兰,罗丽娟,等.广东沿海4种石斑鱼的染色体组型分析[J].广东农业科学,2012(8):124-129.
[16]郑莲,刘楚吾,李长玲.四种石斑鱼染色体核型研究[J].海洋科学,2005,29(4):51-55.
[17]陈毅恒,容寿柏,刘绍琼.鲑点石斑鱼的核型[J].福建水产,1990(1):23-25.
[18]郭丰,王军,苏永全,等.云纹石斑鱼染色体核型研究[J].海洋科学,2006,30(8):1-3.
[19]Guo M L,Wang S F,Su Y Q,et al.Molecular cytogenetic analyses of Epinephelus bruneus and Epinephelus moara(Perciformes,Epinephelidae)[J].Peer J,2014,2:e412.
[20]王云新,王宏东,张海发,等.斜带石斑鱼与赤点石斑鱼的核型研究[J].湛江海洋大学学报,2004,24(3):4-8.
[21]王世锋.六种石斑鱼核型特征比较和染色体进化研究[D].厦门:厦门大学,2007.
[22]蔡岩,周永灿,张本,等.海南野生三斑石斑鱼染色体核型分析[J].热带生物学报,2011,2(2):97-100.
[23]曹丽萍,丁炜东.半微量全血培养法制备罗非鱼染色体标本的条件探索[J].上海水产大学学报,2005,14(4):457-459.
猜你喜欢
当代水产(2022年1期)2022-04-26
生物技术进展(2022年1期)2022-02-11
小资CHIC!ELEGANCE(2021年44期)2021-01-11
当代水产(2020年3期)2020-06-15
当代水产(2019年7期)2019-09-03
学生天地(2019年35期)2019-08-25
当代水产(2019年5期)2019-07-25
科学之谜(2018年9期)2018-12-17
花卉(2015年1期)2015-02-21
遗传(2015年4期)2015-02-04
