
大帛斑蝶成虫行为学特征
2015-02-11 02:06王翻艳李承哲陈晓鸣周成理
生态学报 2015年23期
王翻艳,李承哲,陈晓鸣,周成理
中国林业科学研究院资源昆虫研究所,国家林业局资源昆虫培育与利用重点实验室,昆明 650224
大帛斑蝶成虫行为学特征
王翻艳,李承哲,陈晓鸣*,周成理
中国林业科学研究院资源昆虫研究所,国家林业局资源昆虫培育与利用重点实验室,昆明 650224
在网室内跟踪观察了大帛斑蝶Idealeuconoe成虫的活动规律。结果表明:适于大帛斑蝶羽化的温度为≥23 ℃,羽化主要集中在8:00—9:00,羽化当天蝴蝶极少飞行。羽化第2天开始飞行和觅食,飞行活动最早于7:30便开始有活动,最晚于18:40停止活动,阴天则会推迟开始活动时间,提前结束活动时间。雌蝶日飞行有一个高峰期,在10:00—12:00时段内最活跃,雄蝶则在10:00—12:00和14:00—16:00有2个活跃高峰期。羽化后至产卵前的8d中,大帛斑蝶雌蝶总的飞行次数、时间分别为146次和186.23 min,平均每天飞行次数为18次,飞行时间为23.28 min,单次平均飞行时间为1.28 min;雄蝶总的飞行次数、时间分别为131次和114.43 min,平均每天飞行次数16次,飞行时间为14.30min,单次平均飞行时间为0.87 min。飞行次数和飞行时间在雌雄虫间均无显著性差异(P>0.05);而雌、雄虫每天飞行规律有显著性差异(P<0.05);蝴蝶种群的飞行次数与飞行时间呈显著正相关(P<0.01),飞行次数和单次飞行时间也呈显著正相关(P<0.01)。羽化后至产卵前大帛斑蝶雌蝶访花53次,占飞行次数的36.30%;访花时间258.87 min,超过飞行时间139.01%;平均每天访花7次,占飞行次数的38.89%;访花时间32.36 min,单次访花时间4.88 min。雄蝶访花40次,占飞行次数的30.53%;访花时间344.41 min,超过飞行时间300.98%;平均每天访花5次,占飞行次数的31.25%;访花时间43.05 min,单次访花时间8.61 min。访花次数和访花时间在雌雄虫间无显著性差异(P>0.05),雌、雄虫的每天访花规律有显著差异(P<0.05)。蝴蝶种群的访花次数与访花时间呈显著正相关(P<0.01)。交配出现在羽化后的第6天,主要发生在12:00—16:00时段内,大部分蝴蝶交配持续时间很长,雌雄蝶均可多次交配。雌蝶最早于交配完第2天产卵,卵散产于叶背,产卵可持续2—3个月。
大帛斑蝶;飞行;访花;交配;产卵
大帛斑蝶(Idealeuconoe)隶属于鳞翅目(Lepidoptera)斑蝶科(Danaidae)帛斑蝶属(Idea)。别名:大白斑蝶、大胡麻斑蝶、大笨蝶、傻蝶、熊猫斑蝶。主要分布于台湾,琉球、中南半岛、马来半岛、菲律宾等地区也有分布[1]。其数量较多,且全年可见期长,体型较大,斑纹鲜明,飞行缓慢、优雅,极具观赏价值,是供给蝴蝶生态园的重要品种之一,亦是科普教育的好材料。对大帛斑蝶的研究,国内只有生物学特性[1]和觅食过程中视觉和嗅觉的行为反应研究[2],国外在信息素[3-4]、生物碱[5]、飞行动力学[6]等方面研究较多,在大帛斑蝶行为学研究方面,日本学者结合环境因子对大帛斑蝶的访花[7]、飞行、睡眠[8]行为和日活动节律[9]等进行研究,然而,还未见到对大帛斑蝶进行全面系统的行为学研究报道。
活动节律和活动时间分配是动物行为学研究的两个重要方面[10],动物根据自己的需要和能量与代谢之间的权衡把时间分配到各种活动中去,从而形成了某一种类在某个生境中特殊的活动节律和活动时间的分配方式[11]。本研究将对大帛斑蝶成虫的羽化、飞行、访花、求偶、交配和产卵行为进行观察研究,摸清其日活动规律、逐日活动规律,进而根据其活动特点将成虫期分为不同发育阶段。旨在为蝴蝶旅游景观营造、提高大帛斑蝶人工繁育效率提供理论依据。
1 材料与方法
1.1 试验地点
实验在云南省元江哈尼族彝族傣族自治县中国林业科学研究院资源昆虫研究所元江试验站进行(102°00′46"E,23°36′11"N,海拔400m),年平均气温21 ℃,年平均降水量770mm,属于干热气候类型。实验观察在一个长、宽和高为8 m×8 m×5 m的网室内进行。
1.2 材料
供试蝴蝶:大帛斑蝶Idealeuconoe。
蜜源及寄主植物:马利筋Asclepiascurassavica与爬森藤Parsonsialaevigata。
1.3 试验方法
(1)羽化
在网室内观察大帛斑蝶蛹的羽化状况,雌雄虫样本30头(雌∶雄=1∶1),记载雌雄虫的羽化时间、羽化时的温度。
(2)成虫行为观察
在网室内随机布置20盆开花的马利筋植物作为蜜源植物,1盆爬森藤植物供其产卵。
大帛斑蝶羽化后,用记号笔在翅上标记雌雄虫30头(雌∶雄=1∶1),在布置有蜜源植物和寄主植物的网室中放飞,选择雌雄蝶各3头定位跟踪观察,每天从7:00到19:00进行持续观察,记录大帛斑蝶飞行、访花、求偶、交配和产卵等行为。其余放飞的大帛斑蝶用摄像记录,分析和补充行为学观察。
1.4 数据分析
采用Excel 2010和SPSS 20.0进行数据统计分析,用Kruskal-Wallis进行差异性检验。
2 结果与分析
2.1 羽化
大帛斑蝶临近羽化时,蛹壳逐渐褪去金色,变干变透明,显现出虫体的轮廓和颜色。羽化时,蛹体于背部开裂,头和足依次伸出,足紧紧抓住蛹壳,然后胸腹部随之而出,并停息在蛹壳上,从蛹壳开裂到成虫完全脱离蛹壳只需1 min左右,此时翅柔软而皱缩,腹部膨大,不久即开始慢慢抖动翅膀向上爬行,找合适的位置停息,常有数滴浑浊液体从腹部排出,约5 min左右翅膀完全展开,叠于背部,约1 h后翅变硬。在羽化后至起飞前,蝴蝶经常有向上爬行和扑打翅膀的行为。
大帛斑蝶成虫在白天上午羽化,开始羽化的时间在7∶31,主要集中在8:00—9:00,雄蝶比雌蝶早羽化。成虫羽化与温度和蛹的发育程度密切相关,大帛斑蝶羽化时温度均高于23 ℃(表1)。
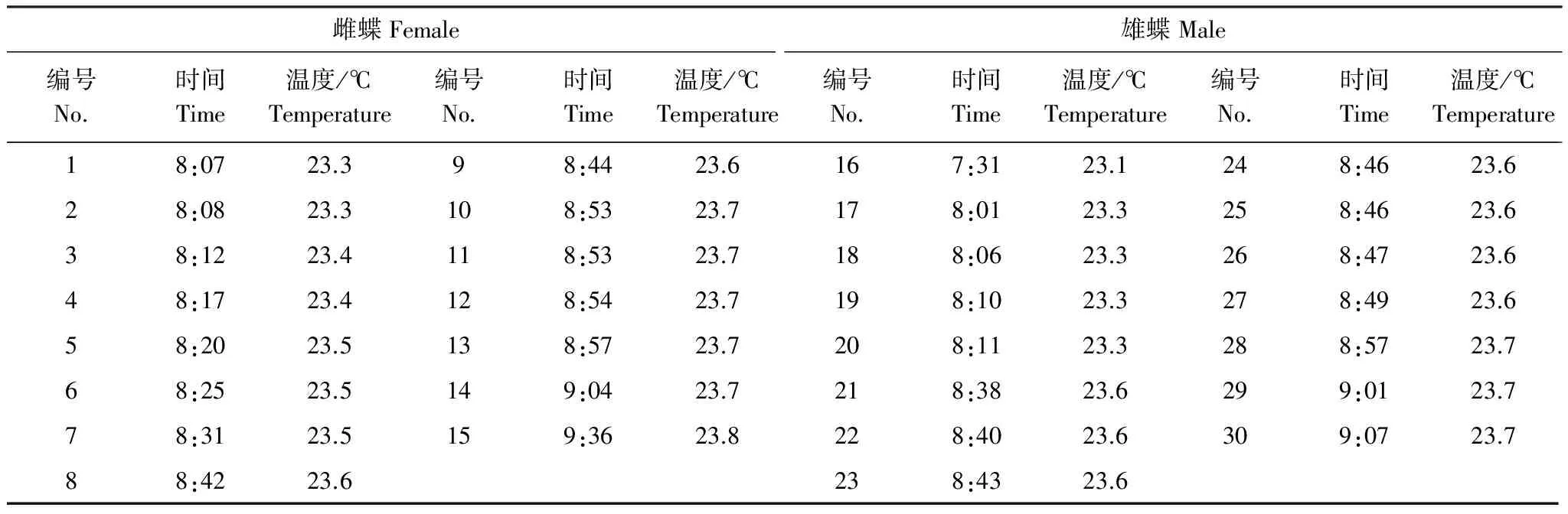
表1 大帛斑蝶羽化时间及温度Table 1 The eclosion time and temperature of Idea leuconoe
2.2 飞行行为
成虫于羽化当天不活跃,羽化5h后开始飞行,只有0—4次飞行,其余时候均处于停息状态。从羽化第2天开始,除第4天和第7天为阴天出现低峰外,随着蝴蝶生长发育,雌雄蝶的飞行次数和时间逐渐增加(图1)。
大帛斑蝶成虫飞行活动最早于7:30便开始有活动,最晚于18:40停止活动,蝴蝶于10:00前活动较少,雌蝶在10:00—12:00时段内最活跃,雄蝶则在10:00—12:00和14:00—16:00有2个活跃高峰期(图2)。阴天则会推迟开始活动时间,提前结束活动时间。
大帛斑蝶在羽化后至产卵前的8d中,雌蝶总的飞行次数、时间分别为146次和186.23 min,平均每天飞行次数为18次,飞行时间为23.28 min,单次平均飞行时间为1.28 min;雄蝶总的飞行次数、时间分别为131次和114.43 min,平均每天飞行次数16次,飞行时间为14.30min,单次平均飞行时间为0.87 min。飞行次数和飞行时间在雌雄虫间均无显著性差异(P>0.05);而雌、雄虫的每天飞行规律有显著性差异(P<0.05);蝴蝶种群的飞行次数与飞行时间呈显著正相关(雌蝶:P<0.01,r=0.838;雄蝶:P<0.01,r=0.855),飞行次数和单次飞行时间也呈显著正相关(P<0.01,r=0.873)。
大帛斑蝶是热带蝴蝶,一天实验中,温度>23 ℃后开始飞行,10:00—12:00飞行活动活跃,此阶段的平均温度为27 ℃,12:00以后,超过30℃飞行活动降低;雄虫在30℃高温下飞行活动较雌虫活跃;羽化后第4天的温度<25 ℃,雌雄蝶的飞行活动受影响(图3,图4)。说明温度与大帛斑蝶飞行活动关系密切。大帛斑蝶飞行活跃时的相对湿度在60%—80%,超过80%,飞行活动受影响(图5,图6)。
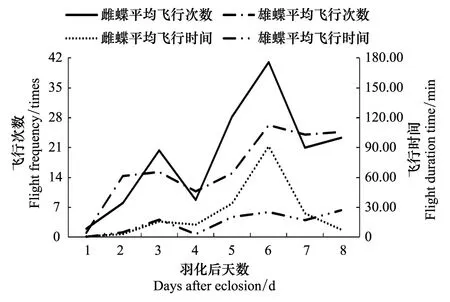
图1 雌雄蝶羽化后日飞行次数和时间 Fig.1 The daily flight frequency and duration time of female and male from eclosion to oviposition

图2 大帛斑蝶的日飞行活动节律Fig.2 The daily flight rhythm
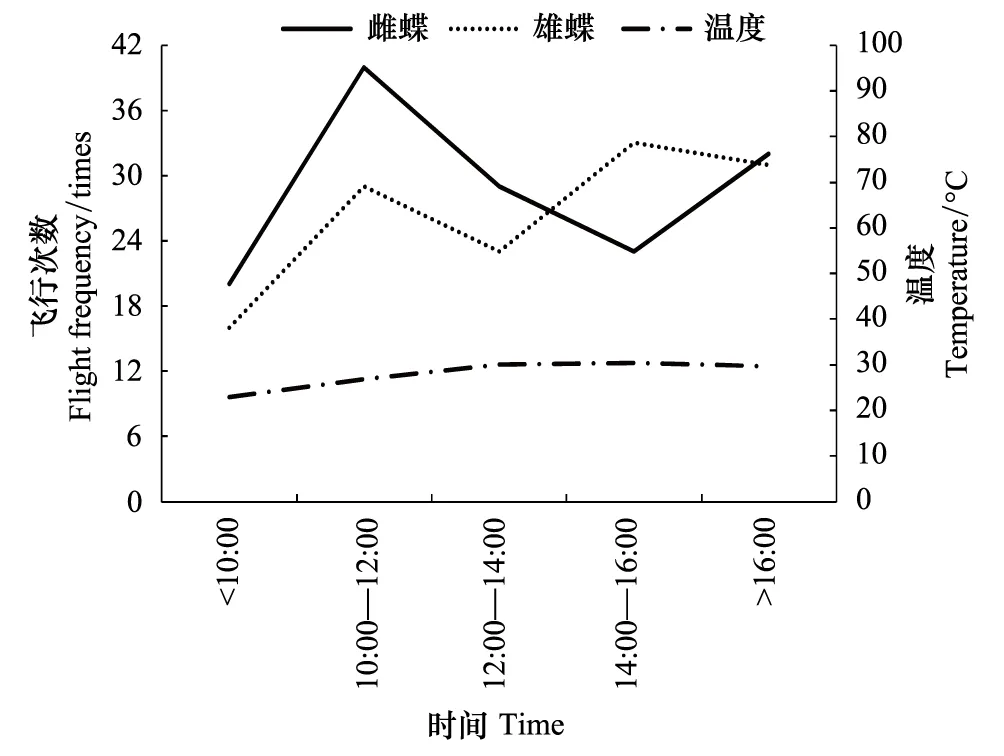
图3 大帛斑蝶日飞行次数与温度Fig.3 The temperature and daily flight frequency

图4 大帛斑蝶羽化后日飞行次数与温度 Fig.4 The temperature and daily flight frequency from eclosion to oviposition
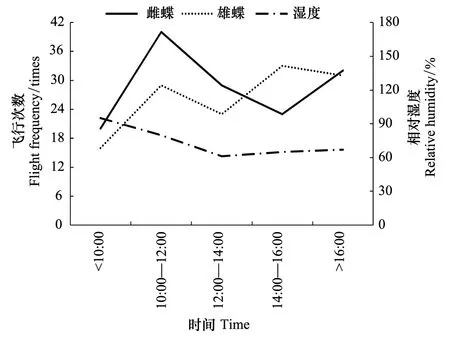
图5 大帛斑蝶日飞行次数与湿度Fig.5 The relative humidity and daily flight frequency
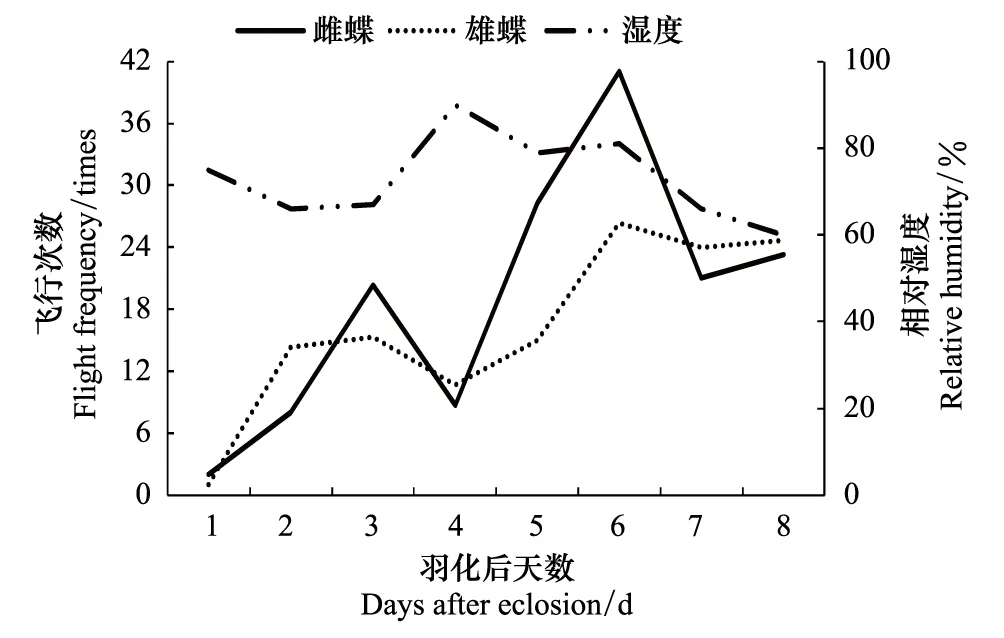
图6 大帛斑蝶羽化后日飞行次数与湿度 Fig.6 The relative humidity and daily flight frequency from eclosion to oviposition
2.3 访花行为
大帛斑蝶雌雄蝶均于羽化后第2天开始出现少量的访花行为,在一天的访花中,10:00前的访花行为最少,10:00后蝴蝶访花活跃,一直持续到16:00时(图7)。
在大帛斑蝶产卵前的成虫期,雌蝶于羽化后第2天开始,雄蝶于第4天开始,种群的访花次数和时间随着成虫生长发育呈现出波浪型增加趋势(图8)。
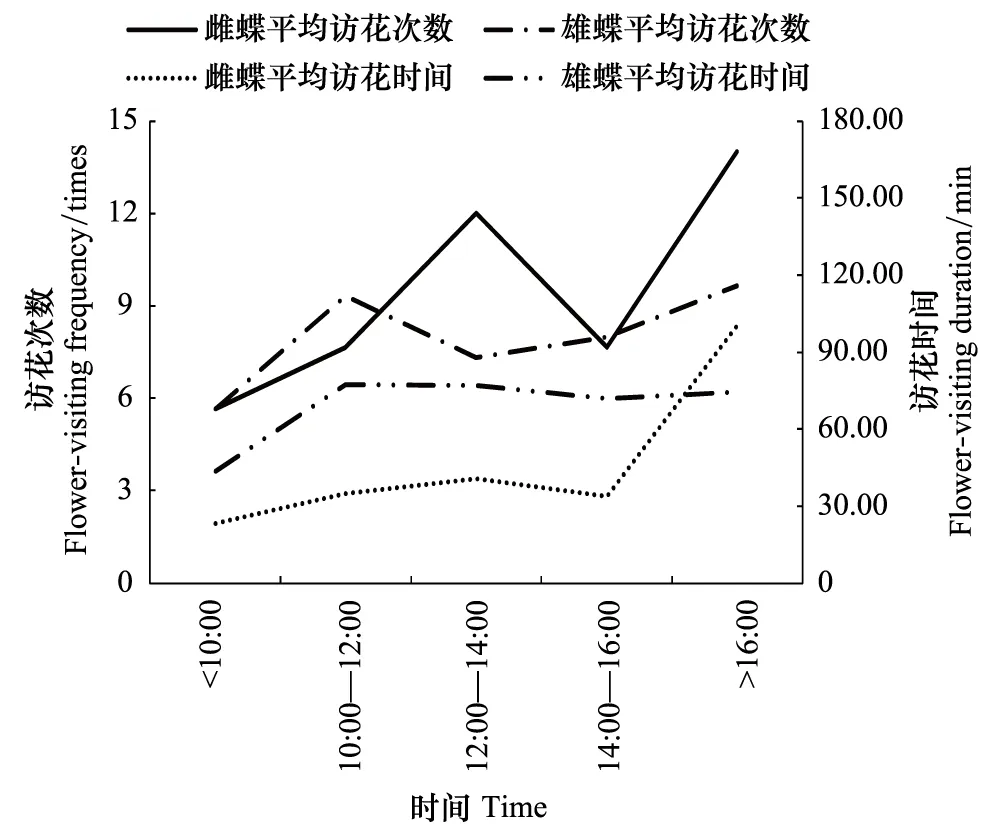
图7 大帛斑蝶日访花节律Fig.7 The daily flower-visiting rhythm
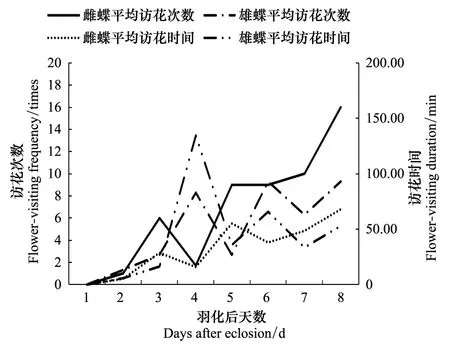
图8 雌雄蝶羽化后日访花次数和时间 Fig.8 The daily flower-visiting frequency and duration time of female and male from eclosion to oviposition
大帛斑蝶在羽化后至产卵前的8d中,雌蝶访花53次,访花时间258.87 min,平均每天访花7次,访花时间32.36 min,单次访花时间4.88 min;雄蝶访花40次,访花时间344.41 min,平均每天访花5次,访花时间43.05 min,单次访花时间8.61 min。访花次数和访花时间在雌雄虫间无显著性差异(P>0.05),雌、雄虫的每天访花规律有显著差异(P<0.05)。蝴蝶种群的访花次数与访花时间呈正相关(雌蝶:P<0.01,r=0.922;雄蝶:P<0.01,r=0.880)。
大帛斑蝶访花次数占飞行次数的比例在20%—40%之间,随着生长发育,访花与飞行的比例逐步提高,雄蝶在羽化后第4天,温度较低时的飞行与访花次数均高于雌蝶,可能是雄蝶对温度的适应性较雌蝶强(图9,图10)。
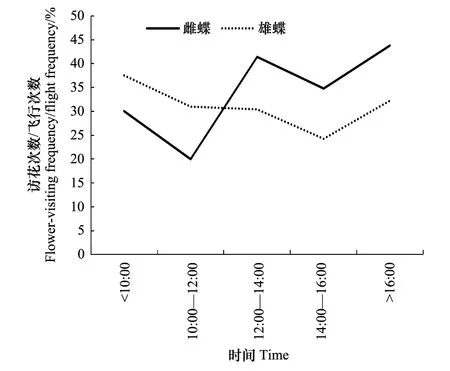
图9 一天不同时段内雌雄虫访花次数与飞行次数比率 Fig.9 The ratio between the flower-visits and the flight times of female and male in different time during a day
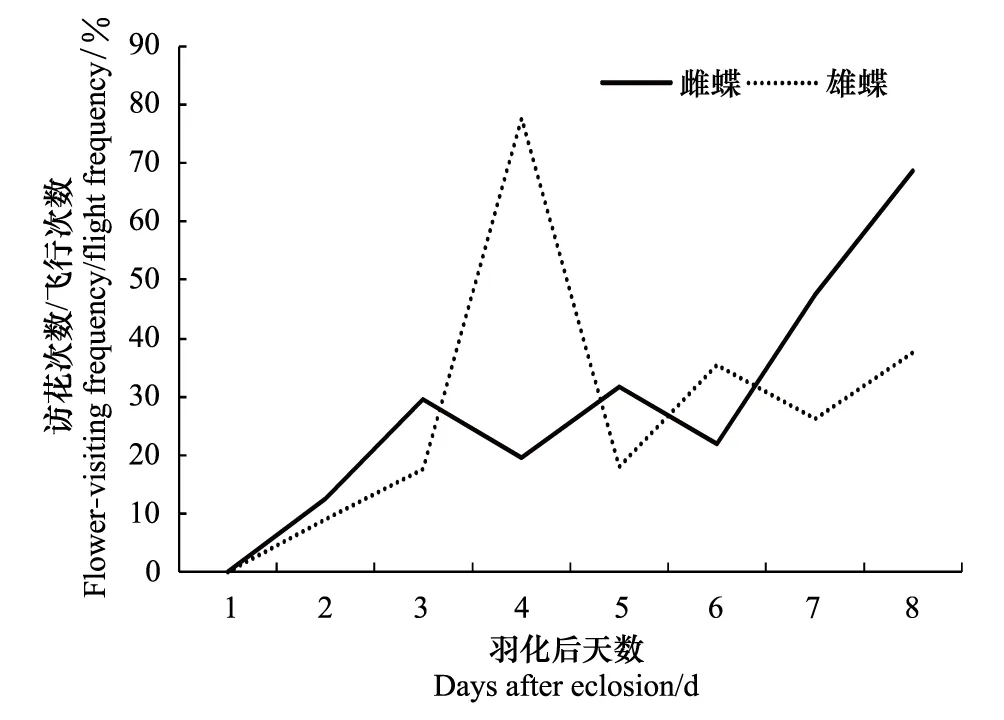
图10 大帛斑蝶羽化后日访花次数与飞行次数比率 Fig.10 The ratio between the flower-visits and the flight times during the days from eclosion to oviposition
2.4 求偶、交配与产卵
大帛斑蝶雄蝶的求偶行为属于巡游型。求偶时,雌蝶在雄蝶的追逐下,若雌蝶有交配意愿时,便会停在网壁、地面等处,雄蝶紧随而至,停靠在雌蝶旁,尾部弯曲伸至雌蝶腹末,交尾时雌蝶也会在后翅下弯曲腹末,这样有利于交尾的顺利进行,否则雌蝶飞离,甩开雄蝶。在求偶高峰,经常会出现多只蝴蝶互相追逐的情景。
大帛斑蝶交配姿势有一字型和并排型,双翅展开或叠合。若交配期间受到干扰,能以未分离状态飞到别处继续交配。大帛斑蝶于羽化后第6天出现交配行为,交配主要发生在一天中的12:00—16:00内。在观察时间段内,观察到2对大帛斑蝶的交配始末时间,交配持续时间分别为185 min和229 min,大多数蝴蝶从白天开始交配,并过夜,到第2天8:00之前才结束交配,交配持续时间很长,有的可达10h以上。大帛斑蝶雌雄虫均能进行多次交配。
大帛斑蝶交配完第2天雌蝶即可产卵,也有交配完第4天产卵的。观察中发现大帛斑蝶有两次交配后产卵和产卵后继续交配的现象。大帛斑蝶卵散产,主要产于叶背面,也有将少数卵产于叶正面、茎杆、盆沿等处。

图11 大帛斑蝶成虫的不同发育阶段Fig.11 The different developmental stages of adults
3 讨论
蝴蝶进入成虫期后,整个生命活动都是围绕着繁衍后代进行,根据蝴蝶成虫的行为,可以将蝴蝶成虫期分为4个阶段,分别是运动器官成熟阶段、补充营养和促进生殖器官发育阶段、求偶和交配阶段、产卵阶段,各阶段内行为有交叉(图11)。
运动器官成熟阶段:大帛斑蝶于羽化5h后开始飞行,且在羽化当天飞行很少,多有爬行、展翅行为,这可能与其运动器官还未成熟、羽化时能量消耗太多有关。蝴蝶于羽化当天并不访花,可能是蝴蝶羽化后,运动器官需要通过飞翔而达到发育成熟,以锻炼飞翔能力和适应环境,这时蝴蝶主要依靠蛹积累的能量来完成,而不需要访花来补充营养。这个阶段影响蝴蝶行为的主要因素是温度。蝴蝶是日出性昆虫,它们的体温与活动都受外界温度的支配,在阴雨天一般休止不飞,有时太阳突然被乌云遮蔽,蝴蝶也会立即作出反应,太阳出来后便又开始活动[12-14]。大帛斑蝶雄蝶对温度的适应性较雌蝶强,在一天的飞行活动中,雌虫高峰期在上午,之后活动逐步下降,而雄虫有2个高峰期,除中午温度(12:00—14:00)太高活动减少外,均处于活跃期(图3)。雌、雄虫的每天飞行规律有显著差异,雄虫活动时间较雌虫长。
补充营养和促进生殖器官发育阶段:羽化后的第2天飞行活动逐渐增多,出现少量的访花行为,开始补充营养,第3天开始,蝴蝶飞行访花行为活跃,访花次数、时间随着羽化后天数逐渐增加(图8),这个阶段蝴蝶以补充营养为主,通过飞翔和补充营养促进生殖器官的发育。这个阶段温度仍然是飞翔活动的主要影响因子,随着生长发育,大帛斑蝶的飞行活动呈现波浪型增长(图1),而原因是温度呈现波浪型变化造成的。这个阶段大帛斑蝶的访花和飞行次数的比例增加(图10),访花时间稳步增加,蜜源植物充足对生殖器官发育有重要影响[15-19]。
求偶和交配阶段:大帛斑蝶于羽化后第6天发生交配行为,此前访花次数和访花时间总体呈上升趋势(图8),说明蝴蝶在进入交配期后,蝴蝶继续补充营养,促进精子和卵的发育和成熟,可增大产卵量[20]。这个阶段大帛斑蝶的飞行次数的降低,可能是为蝴蝶交配积蓄能量。蝴蝶飞行是通过间接飞行肌的收缩,产生胸体积的变化,进而引起的翅膀运动[21]。飞行行为随着飞行肌的增加而增强,但飞行肌的发育可阻碍其它行为,尤其是雌虫的繁殖力[22]。飞行肌的降解是一种普遍现象,降解的能量和产物会参与生殖行为,但伴随着飞行肌的降解,蝴蝶飞行能力开始下降,尤其是雌蝶[23]。
产卵阶段:大帛斑蝶羽化后第9天开始产卵。产卵高峰较交配高峰延后1—2 d[3]。大帛斑蝶交配和产卵期很长,可延续2—3个月,在这期间,大帛斑蝶不断地访花补充营养,重复交配和产卵。在较长的产卵期中,在云南元江7—8月高温时,常在树下栖息不动,待温度降低后才活动,持续高温对大帛斑蝶生殖影响巨大(不产卵或者产不孕卵),高温是否对大帛斑蝶成虫有滞育作用,有待研究。
[1] 佘震加. 厦门岛蝴蝶种类调查与大帛斑蝶生物学特性研究 [D]. 福建农林大学, 2008:16- 26.
[2] Tang Y C, Zhou C L, Chen X M, Zheng H. Visual and olfactory responses of seven butterfly species during foraging. Journal of Insect Behavior, 2013, 26(3):387- 401.
[3] Nishida R, Schulz S, Kim C S, Fukami H, Kuwahara Y, Honda K, Hayashi N. Male sex pheromone of a giant danaine butterfly,Idealeuconoe. Journal of Chemical Ecology, 1996, 22(5):949- 972.
[4] Mineeva I V. Methyl (3R,5R)-3,5-dihydroxydecanoate in the asymmetric synthesis ofIdealeuconoepheromone and formal syntheses of (+)-(3R,5R)-3-hydroxydecano-5-lactone, verbalactone, and Tolypothrix pentaether. Russian Journal of Organic Chemistry, 2013, 49(6):838- 842.
[5] Honda K, Hayashi N, Abe F, Yamauchi T. Pyrrolizidine alkaloids mediate host-plant recognition by ovipositing females of an old world danaid butterfly,Idealeuconoe. Journal of Chemical Ecology, 1997, 23(7):1703- 1713.
[6] Wilson T, Albertani R. Wing-flapping and Abdomen Actuation Optimization for Hovering in the ButterflyIdealeuconoe//Region VI AIAA Student Paper Conference, Salt Lake City, UT, 2013:1- 11.
[7] 瀬田和明, 落合米子, 寺口芳一. マダラチョウの訪花活動について:(2)温室内におけるオオゴマダラ,スジグロカバマダラの日周活動と環境条件. 蝶と蛾, 1999, 50(3):235- 242.
[8] 津吹卓, 瀬田和明. 温室内におけるオオゴマダラの飛翔および睡眠に関する活動と環境条件. 蝶と蛾, 2001, 52(3):171- 182.
[9] 瀬田和明, 落合米子, 寺口芳一. 冬期の放蝶温室におけるオオゴマダラの日周活動. 蝶と蛾, 2004, 55(3):160- 165.
[10] Halle S, Stenseth N C. Activity Patterns in Small Mammals:An Ecological Approach. Berlin Heidelberg:Springer-Verlag, 2000:39- 39.
[11] 王力军, 于丰军, 洪美玲, 吴山, 陈旭. 人工饲养条件下蜡皮蜥昼间行为时间分配及活动规律. 四川动物, 2005, 24(1):9- 13.
[12] 杨慧英, 黄模华, 刘良源. 大自然的舞姬——蝴蝶的生物学习性. 江西植保, 2010, 33(2):88- 89.
[13] 霍晓杰. 青海祁连山地区四川绢蝶的生物学特性及各因素对其活动影响的研究 [D]. 西宁:青海师范大学, 2010:19- 31.
[14] Cormont A, Malinowska A H, Kostenko O, Radchuk V, Hemerik L, WallisDeVries M F, Verboom J. Effect of local weather on butterfly flight behaviour, movement, and colonization:significance for dispersal under climate change. Biodiversity and Conservation, 2011, 20(3):483- 503.
[15] 王竑晟, 徐洪富, 崔峰. 成虫期营养对甜菜夜蛾生殖力及卵巢发育的影响. 西南农业学报, 2004, 17(1):34- 37.
[16] Fischer K, O′Brien D M, Boggs C L. Allocation of larval and adult resources to reproduction in a fruit-feeding butterfly. Functional Ecology, 2004, 18(5):656- 663.
[17] O′Brien D M, Boggs C L, Fogel M L. Making eggs from nectar:connections between butterfly life history and the importance of nectar carbon in reproduction. Oikos, 2004, 105(2):279- 291.
[18] Mevi-Schütz J, Erhardt A. Amino acids in nectar enhance butterfly fecundity:a long-awaited link. American Naturalist, 2005, 165(4):411- 419.
[19] Bauerfeind S S, Fischer K, Hartstein S, Janowitz S, Martin-Creuzburg D. Effects of adult nutrition on female reproduction in a fruit-feeding butterfly:The role of fruit decay and dietary lipids. Journal of Insect Physiology, 2007, 53(9):964- 973.
[20] Boggs C L. Reproductive strategies of female butterflies:variation in and constraints on fecundity. Ecological Entomology, 1986, 11(1):7- 15.
[21] Correa-Fernandez F, Cruz-Landim C. Differential flight muscle development in workers, queens and males of the eusocial bees,ApismelliferaandScaptotrigonapostica. Journal of Insect Science, 2010, 10(1):85- 85.
[22] Marden J H. Variability in the size, composition, and function of insect flight muscles. Annual Review of Physiology, 2000, 62(1):157- 178.
[23] Stjernholm F, Karlsson B. Flight muscle breakdown in the green-veined white butterfly,Pierisnapi(Lepidoptera:Pieridae). European Journal of Entomology, 2008, 105(1):87- 91.
Behavior observation ofIdealeuconoeadults
WANG Fanyan, LI Chengzhe, CHEN Xiaoming*, ZHOU Chengli
TheResearchInstituteofResourcesInsectoftheChineseAcademyofForestry;KeyLaboratoryofBreedingandUtilizationofResourceInsect,StateForestryAdministration,Kunming650224,China
Adult behaviors ofIdealeuconoewas observed by the tracking method in a netted room. The results showed that the initial temperature of eclosion was 23 °C and that eclosion mainly occurred at 08:00—09:00. The pupal shell faded to a gold color, and became dry and transparent as animals approached emergence, which revealed the contour and color of the butterfly. Just 1 min was required from the cracking of the puparium to adult emergence from the puparium. The wings were fully extended after approximately 5 min, and were stacked on the back, with a further 1 h being required for the wings to harden. We observed some flight, but no foraging behavior, on the first day of eclosion, with adult insects remaining in a resting state for most of the time. Both male and female butterflies began to forage on the second day after eclosion. Flight activity began as early as 07:30, and stopped as late as 18:40. However, on cloudy days, flight activity began later and stopped earlier. The daily peak time for flying by females was at 10:00—12:00, while two flying peaks were observed for males, at 10:00—12:00and 14:00—16:00. Flight behavior was affected when temperatures fell below 25 °C or rose above 30°C. Male butterflies tolerated higher temperatures compared to female butterflies since males were more active than females at 30°C. Except for cloudy days, the flight frequency and flight duration of both males and females increased gradually until mating. In the eight days from eclosion to oviposition, the total flight frequency and accumulated flying time of females were 146 times and 186.23 min, flying on average 18 times a day with a cumulative time of 23.28 min per day, resulting in 1.28 min per flight. Similarly, males flew 131 times for 114.43 min during these eight days, on average 16 times and 14.30min per day, resulting in 52 seconds per flight. There was no significant difference (P> 0.05) between males and females with respect to total flight frequency or flight duration, while this difference was significant among different days (P< 0.05). A positive significant correlation existed between flight frequency and duration time (P< 0.01). In addition, a positive correlation was seen between flight frequency and single flight time (P< 0.01) in the butterfly population. During the eight days from eclosion to oviposition, females visited flowers 53 times, which took up 36.30%of the total flying times. Total feeding time on flowers was 258.87 min, which was equivalent to 139%of total flight duration time. Seven visits took up 38.9%of total daily flying times, representing 32.36 min per day for flower visits and 4.88 min per visit. Males visited flowers 40times, which was 30.53%of total flying times, with 344.41 min in total spent feeding on flowers, representing 300.98%of overall flying time. Five visits took up 31.25%of total daily flying times, representing 43.05 min per day for flower visits and 8.61 min per visit. There was no significant difference (P> 0.05) between males and females with respect to flight and feeding times. On the sixth day after eclosion, mating mainly occurred at 12:00—16:00. Most mating encounters were very long, some of which lasted more than 10hours. Both male and female butterflies mated more than once. If the mating process was disturbed, butterflies were able to change location by flying elsewhere in an unseparated state. The first oviposition occurred on the second day after mating, while the last oviposition occurred on the fourth day after mating. Most eggs were deposited in a dispersed manner on the back of host leaves.
Idealeuconoe; flight frequency; flower-visiting; mating; oviposition
云南省社会发展计划(2011CA023)
2014- 04- 24; < class="emphasis_bold">网络出版日期:
日期:2015- 05- 19
10.5846/stxb201404240812
*通讯作者Corresponding author.E-mail:cafcxm@139.com
王翻艳,李承哲,陈晓鸣,周成理.大帛斑蝶成虫行为学特征.生态学报,2015,35(23):7642- 7649.
Wang F Y, Li C Z, Chen X M, Zhou C L.Behavior observation ofIdealeuconoeadults.Acta Ecologica Sinica,2015,35(23):7642- 7649.
猜你喜欢
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
湖北植保(2017年4期)2017-08-31
小溪流(画刊)(2017年6期)2017-07-10
小学阅读指南·低年级版(2017年1期)2017-03-13
文理导航·科普童话(2016年4期)2016-05-31
小朋友·快乐手工(2015年11期)2016-01-07
中国马铃薯(2015年3期)2015-12-19
广西蚕业(2015年3期)2015-11-30
