
猪APOBEC3F 基因外显子8 单核苷酸的多态性及其与猪繁殖和呼吸综合征病毒易感性的关联性
2014-12-23 11:30朱前明孟春花茆达干王慧利李静心婧1王亚磊1
江苏农业学报 2014年5期
朱前明,孟春花, 茆达干, 王慧利, 李静心, 王 婧1,, 王亚磊1,,
曹少先1,3
(1.江苏省农业科学院畜牧研究所,江苏 南京210014;2.南京农业大学动物科学与技术学院,江苏 南京210095;3.江苏省农业科学院动物品种改良和繁育重点实验室,江苏 南京210014)
载脂蛋白质B mRNA 编辑酶催化多肽3F(APOBEC3F)是APOBEC 家族的一员,具有广泛的抗病毒活性,可有效地抑制人类免疫缺陷病毒(HIV)、乙型肝炎病毒(HBV)、猪内源性逆转录病毒(PERV)等病毒的复制,在宿主抗病毒天然免疫应答中发挥着重要作用[1-4]。APOBEC3F 含有2 个相同的胞嘧啶脱氨基模体即His/Cys-X-Glu-X23-28-Pro-Cys-X2-Cys,分别位于基因编码蛋白质的第85 ~119 位和265 ~299 位。C 端脱氨基模体是其发挥脱氨基作用的主要活性区域,N 端模体脱氨基作用较弱,主要与APOBEC 分子衣壳化、RNA 结合及二聚体的形成有关,但两者都是发挥脱氨基功能必不可少的部分[5-8]。其主要作用机制是细胞内APOBEC3F 分子通过gag 蛋白质[6]、病毒RNA[7]、细胞RNA[8]的媒介进入成熟病毒粒子的核心,并在随后感染的靶细胞中使新合成的负链cDNA 发生C/T 突变,从而导致新合成的DNA 降解或产生大量致死性的G/A 突变来抑制病毒的复制与感染[9-12]。
有研究结果表明APOBEC3F基因的抗逆转录病毒特性具有普遍性[13-14]。猪繁殖与呼吸综合征(Porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(PRRSV)感染引起的一种高度传染性疾病。2006 年中国南方暴发的猪高热病对中国养猪业造成巨大的经济损失,其病原就是变异的高致病性PRRSV[15-16]。通过分离猪肺泡巨噬细胞体外攻毒PRRSV 的方法来分析猪PRRSV 易感性差异是一种有效的试验模型[17]。猪APOBEC3F基因虽然已经被克隆[18-19],但关于APOBEC3F基因的多态性及其与PRRSV 易感性的关联研究尚未见报道。本研究对多个猪种APOBEC3F基因外显子3、4、8 和内含子3 进行测序,寻找多态性位点,并分析其与PRRSV 易感性的关联性,以期检测到与PRRSV 易感性相关的分子标记,为抗PRRSV 猪育种提供依据。
1 材料与方法
1.1 材料
试验猪包括9 个品种,共193 头,其中杜洛克猪36 头(购自杭州大关猪场)、长白猪42 头(购自杭州大关猪场)、大白猪38 头(购自杭州大关猪场)、苏钟猪34 头(购自江苏省农业科学院动物科学基地猪场)、姜曲海猪14 头(购自江苏姜曲海猪场)、定远猪12 头(购自安徽定远县猪场)、二花脸猪5 头(购自江苏省常熟市畜禽良种有限责任公司)、梅山猪5 头(购自江苏太仓市种猪场)、三元杂交猪7 头(购自镇江丹徒区云立牧业有限公司)。每个个体取耳组织于冻存管中,置于液氮中保存。PRRSV NJGC 株由江苏省农科院兽医研究所提供。
1.2 方法
1.2.1 组织样DNA 提取 耳组织样的DNA 提取采用酚/氯仿抽提法,TE buffer 溶解后,-20 ℃保存。
1.2.2APOBEC3F基因外显子3、4 和8 扩增 参照猪APOBEC3F基因外显子3、4、8 和内含子3 序列分别设计A3F-A、A3F-B 2 对引物,由上海捷瑞公司合成,引物信息见表1。PCR 反应总体系20.0 μl,其中含模板DNA 为60 ng,2.5 U/μl PrimeSTAR® HS DNA Polymerase 为0.2 μl,2 mmol/L dNTPs 为1.6 μl,上、下游引物(10 μmol/L)各0.4 μl,5× Prime-STAR®Buffer(包含Mg2+)为4.0 μl,最终加灭菌蒸馏水至20.0 μl。
PCR 反应程序:98 ℃变性10 s;各引物最适温度复性5 s,72 ℃延伸1 min 20 s,35 个循环;最后72 ℃延伸10 min。PCR 产物经1%琼脂糖凝胶电泳,溴化乙锭(EB)染色,检测扩增结果。
1.2.3 多态位点的测序和筛选 每6 头猪DNA 等量混合构建DNA 池,构建3 个不同的DNA 池,进行PCR 扩增,电泳纯化后的PCR 产物送上海美吉生物医药科技有限公司测序。
1.2.4 PCR-RFLP 分析 根据测序结果选择外显子8 的5 bp 处的多态位点,建立PCR-RFLP 检测方法,引物Nrul-SNP 序列见表1。PCR 反应总体系20.0 μl,含模板DNA 60 ng,Taq聚合酶(5 U/μl)0.2 μl,dNTPs(10 mmol/L)0.4 μl,上、下游引物(10 μmol/L)各0.6 μl,MgCl2(25 mmol/L)1.4 μl,10 ×缓冲液2.0 μl,加灭菌蒸馏水至20.0 μl。PCR 扩增条件为:95 ℃预变性5 min;95 ℃变性30 s,引物最适温度复性30 s,72 ℃延伸30 s,共35 个循环;最后72 ℃延伸7 min。PCR 产物经2%琼脂糖凝胶电泳,溴化乙锭(EB)染色,检测扩增结果。
用Nrul酶对引物Nrul-SNP 扩增的PCR 产物进行酶切分型。酶切反应体系为16.0 μl:含PCR 产物5.0 μl,Nrul(10 U/μl)0.5 μl,10 × T buffer 1.0 μl,灭菌蒸馏水9.5 μl,37 ℃反应4 h。2%琼脂糖凝胶电泳检测酶切产物,EB 染色,培清JS-780 全自动凝胶成像分析仪进行拍照分析。
1.2.5 PRRSV 易感性测定 参照Ait-Ali 等[17]的方法进行。简要步骤如下:分离猪肺泡巨噬细胞,接种PRRSV 病毒,分别在接毒后6 h、12 h、18 h、24 h和36 h 收集细胞,提取RNA 反转录成cDNA。针对β-actin基因、PRRSV 毒株N基因序列分别设计特异性引物β-actin和PRRSV-N(表1,由上海捷瑞公司合成)。以cDNA 为模板PCR 扩增基因片段,克隆至T 载体,以此为模板,SYBR Green I 染料法进行定量PCR(ABI7300),绘制标准曲线。以PRRSVN基因的拷贝数与β-actin基因拷贝数的比来计算病毒载量,作为易感性的参数。每组3 个重复。反应体系为20.0 μl,其中SYBR PremixEx Taq10.0 μl,上、下游引物(10 μmol/L)各0.6 μl,ROX 0.4 μl,模板2.0 μl,灭菌去离子水6.4 μl。反应条件为:95℃预变性30 s;95 ℃5 s,56 ℃31 s,40 个循环,每个循环结束时收集荧光信号。

表1 引物信息Table 1 The information of primers used in this study
1.2.6 数据分析 不同猪种群体不同基因型的PRRSV 易感性数据分别用SPSS17.0 软件的One-Way ANOVA 进行关联分析。
2 结果与分析
2.1 APOBEC3F 基因片段的扩增及SNP 筛选
对猪APOBEC3F基因内含子3 进行PCR 扩增,在1 000 ~2 000 bp 间出现1 条特异性条带(图1)与预期结果一致。对猪APOBEC3F基因外显子8 进行PCR扩增,在1 000 ~2 000 bp 间出现1 条特异性条带(图2)与预期结果一致。测序结果显示,在内含子3 的16 bp 处出现1 个A/G 突变(图3)。在外显子8 的5 bp处出现1 个G/A 突变(图4)。经分析外显子8 的5 bp处出现G/A 突变处于Nrul酶切位点内,而内含子3 的16 bp 处A/G 突变引物不易设计,即使用人为创造酶切位点的方法也不能找到合适的限制性内切酶。
2.2 PCR-RFLP 分析
针对外显子8 的5 bp 处G/A 的单碱基突变设计特异性引物,命名为Nrul-SNP。对猪DNA 进行PCR 扩增,在500 bp 与750 bp 之间得到1 条特异性条带(图5),与预期结果一致。用限制性内切酶Nrul对引物Nrul-SNP 扩增的PCR 产物进行酶切分型,以PCR 产物作为对照电泳,AA 型为1 个572 bp片段;GG 型为379 bp 和193 bp 2 个片段;GA 型为572 bp、379 bp 和193 bp 3 个片段(图6)。
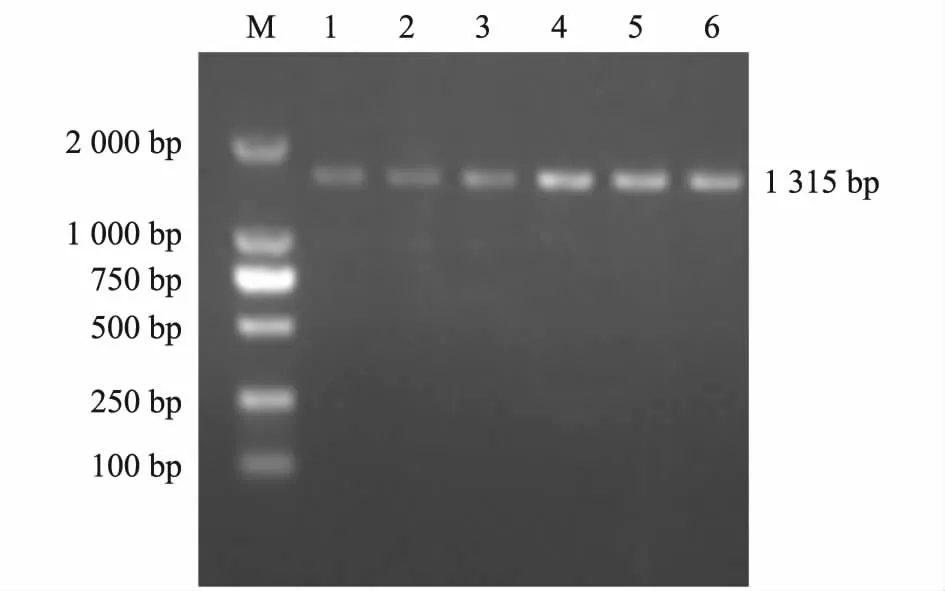
图1 猪APOBEC3F 基因内含子3 PCR 扩增产物电泳图Fig.1 PCR amplification of APOBEC3F intron three
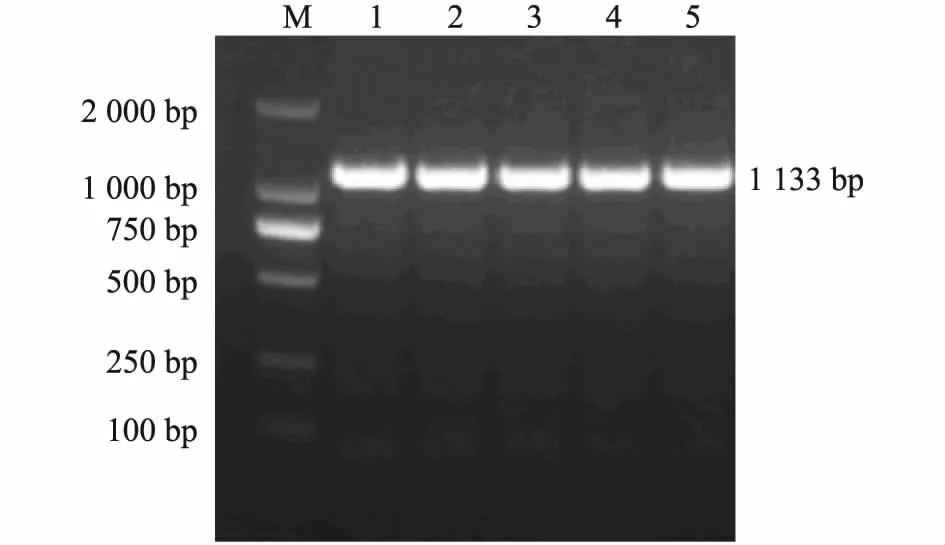
图2 猪APOBEC3F 基因外显子8 PCR 扩增产物电泳图Fig.2 PCR amplification of APOBEC3F exon eight

图3 APOBEC3F 基因内含子3 单碱基突变Fig.3 Single-base mutation in intron three of APOBEC3F gene

图4 APOBEC3F 基因外显子8 单碱基突变Fig.4 Single-base mutation in exon eight of APOBEC3F gene

图5 Nrul-SNP 引物PCR 产物电泳图Fig.5 PCR amplification with primer Nrul-SNP
2.3 APOBEC3F 基因群体的遗传特征
2.3.1 基因频率及基因型频率在猪群体中的分布
在各猪种群体中外显子8 的5 bp 处可以检测到3种基因型,以GG 型居多,GA 型次之,AA 最少;定远猪和梅山猪群体中检测到3 种基因型;二花脸猪群体中检测到GG 和GA 2 种基因型;其他猪群体只检测到GG 1 种基因型。在所有被检测猪种群体中,G等位基因的频率均高于A等位基因,因此G为优势等位基因(表2)。

图6 PCR 产物的Nrul 酶切分型Fig.6 Identification of PCR products with Nrul

表2 APOBEC3F 基因外显子8 的5 bp 处基因型频率、等位基因频率及群体遗传特征Table 2 Genotypic frequency and allelic frequency at 5 bp of exon eight of APOBEC3F gene and population inheritance character

表3 不同基因型猪PAM 接毒后PRRSV 拷贝数Table 3 The relative amount of PRRSV in porcine alveolar macrophage(PAM)of different genotypes of pig
2.3.2 遗传特性 外显子8 的5 bp 处在定远猪、梅山猪和二花脸猪群体中为中度多态(0.25 <PIC<0.50),而在其他群体中未表现出多态性,表明该多态位点在定远猪、梅山猪和二花脸猪群体中存在较强的选择潜力(表2)。采用χ2适合性检验检测发现该基因座位在定远猪、二花脸猪和梅山猪群体中均未偏离Hardy-Weinberg 平衡(P>0.05)。
2.4 不同基因型猪种群PRRSV 易感性
比较APOBEC3F基因外显子8 的5 bp 处不同基因型猪肺泡巨噬细胞(PAM)接毒后PRRSV 拷贝数(表3)发现,AA 基因型猪PAM 细胞接毒后12 h PRRSV 拷贝数显著高于GG 型(P<0.05),极显著高于GA 基因型(P<0.01)。
3 讨论
大量研究结果表明APOBEC3F基因具有抑制HIV[1]、PERV[3]、MLV[20]等反转录病毒复制的作用。Jonsson 等[14]研究发现偶蹄动物牛、羊和猪APOBEC3F 主要定位在细胞质,并能抑制HIV-1 和MLV 的复制。王雄虎等[21]研究发现APOBEC3F基因第4 外显子536 位点T/C 突变可能与感染HBV易感性有关。而猪APOBEC3F基因多态性及其与PRRSV 抗性相关性的研究尚未见报道。本研究以APOBEC3F基因为候选基因,结合DNA 池PCR 扩增产物测序方法,发现了猪APOBEC3F基因2 个单碱基突变(内含子3 的16 bp 处出现A/G 突变和外显子8 的5 bp 处出现G/A 突变)。外显子8 的5 bp处可以检测到3 种基因型,以GG 型居多,GA 型次之,AA 最少,且该位点多态性在不同猪种间存在明显的差异。其他种猪群体中没有多态性,可能与长期的人工选育有关。内含子3 的16 bp 处的A/G 突变找不到合适的限制性内切酶来设计PCR-RFLP 或者CRS-PCR-RFLP 方法,因此该位点还需探索其他的检测方法以进一步研究。
比较外显子8 的5 bp 处不同基因型猪PAM 细胞接毒后PRRSV 拷贝数发现,AA 型猪PAM 细胞接毒后12h PRRSV 拷贝数显著高于GG 基因型(P<0.05),极显著高于GA 基因型。表明AA 型猪PRRSV 易感性高于GG 型和GA 型,提示G等位基因更有利于抵抗PRRSV 的感染。
通过数据库查询对比可以得出,本研究检测到外显子8 的5 bp 处G/A 突变是有义突变,导致APOBEC3F基因所编码蛋白质的第391 位的丙氨酸(Ala)变成苏氨酸(Thr)。有研究结果[12]显示APOBEC3F 含有两个胞嘧啶脱氨基模体,并且C 端脱氨基模体是其发挥脱氨基作用的主要活性区域,N 端模体脱氨基作用较弱,主要与APOBEC 分子衣壳化、RNA 结合及二聚体的形成有关。虽然该处的氨基酸改变不在脱氨基结构域上,但有研究显示失去胞苷脱氨基作用的APOBEC3F 分子依然具有抗病毒作用[22],APOBEC3F 分子的脱氨基模体是其发挥抗病毒作用的分子基础,但并不是其抗病毒作用的唯一途径。A5T 突变是否通过其他途径改变APOBEC3F 的抗病毒作用还有待进一步研究和探讨。
[1] HULTQUIST J F,LENGYEL J A,REFSLAND E W,et al.Human and rhesus APOBEC3D,APOBEC3F,APOBEC3G,and APOBEC3H demonstrate a conserved capacity to restrict Vif-deficient HIV-1[J].J Virol,2011,85(21):11220-11234.
[2] XIANG SL,MA Y,YAN Q,et al.Construction and characterization of an infectious replication competent clone of porcine endogenous retrovirus from Chinese miniature pigs[J].J Virol,2013,10:228.
[3] LEE J,CHOI J Y,LEE H J,et al.Repression of porcine endogenous retrovirus infection by human APOBEC3 proteins[J].Biochem Biophys Res Commun,2011,407(1):266-270.
[4] HARRIS R S,BISHOP K N,SHEEHY A M,et al.DNA deamination mediates innateimmunity to retroviral infection[J].Cell,2003,113(6):803-809.
[5] HACHE G,LIDDAMENT M T,HARRIS R S.The retroviral hypermutation specificity of APOBEC3F and APOBEC3G is governed by the C-terminal DNA cytosine deaminase domain[J].J Biol Chem,2005,280(12):10920-10924.
[6] CEN S,GUO F,NIU M,et al.The interaction between HIV-1 Gag and APOBEC3G[J].J Biol Chem,2004,279:33177-33184.
[7] LUO K,LIU B,XIAO Z,et al.Amino-terminal region of the human immuno-deficiency virus type 1 nucleocapsid is required for human APOBEC3G packaging[J].J Virol,2004,78:11841-11852.
[8] IWATANI Y,TAKEUCHI H,STREBEL K,et al.Biochemical activities of highly purified,catalytically active human APOBEC3G:correlation with antiviral effect[J].J Virol,2006,80:5992-6002.
[9] WANG T,ZHANG W,TIAN C,et al.Distinct viral determinants for the packaging of human cytidine deaminases APOBEC3G and APOBEC3C[J].Virology,2008,377:71-79.
[10] HARRIS R S,LIDDAMENT M T.Retroviral restriction by APOBEC proteins[J].Nat Rev Immunol,2004,4(11):868-877.
[11] LACKEY L,LAW E K,BROWN W L,et al.Subcellular localization of the APOBEC3 proteins during mitosis and implications for genomic DNA deamination[J].Cell Cycle,2013,2(5):762-772.
[12] SONG C,SUTTON L,JOHNSON M E,et al.Signals in APOBEC3F N-terminal and C-terminal deaminase domains each contribute to encapsidation in HIV-1 virions and are both required for HIV-1 restriction[J].J Biol Chem,2012,287(20):16965-16974.
[13] FRANCA R,SPADARI S,MAGA G.APOBEC deaminases as cellular antiviral factors:a novel natural host defense mechanism[J].Med Sci Monit,2006,12(5):92-98.
[14] JONSSON S R,HACHE G,STENGLEIN M D,et al.Evolutionarily conserved and non-conserved retrovirus restriction activities of artiodactyl APOBEC3F proteins[J].Nucleic Acids Res,2006,34(19):5683-5694.
[15] TIAN K,YU X,ZHAO T,et al.Emergence of fatal PRRSV variants:unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark[J].PLoS One,2007,2(6):e526.
[16] 童光志,周艳君,郝晓芳,等.高致病性猪繁殖与呼吸综合征病毒的分离鉴定及其分子流行病学分析[J].中国预防兽医学报,2007(5):323-326.
[17] AIT-ALI T,WILSON A D,WESTCOTT D G,et al.Innate immune responses to replication of porcine reproductive and respiratory syndrome virus in isolated swine alveolar macrophages[J].Viral Immunol,2007,20(1):105-118.
[18] 孙 宇,罗 佳,马玉媛,等.猪APOBEC3F基因克隆及真核表达载体的构建与鉴定[J].第三军医大学学报,2011,33(18):1904-1906.
[19] 吴小霞,马义才.猪APOBEC3F基因电子克隆[J].生物信息学,2008(3):100-112.
[20] DOEHLE B P,SCHAFER A,WIEGAND H L,et al.Differential sensitivity of murine leukemia virus to APOBEC3-mediated inhibition is governed by virion exclusion[J].J Virol,2005,79(13):8201-8207.
[21] 王雄虎,游志毅,周军华,等.APOBEC3F基因多态性与HBV感染易感性关系的探讨[J].中国热带医学,2013(4):471-483.
[22] REBECCA K H,FRANSJE A K,KATE N B,et al.APOBEC3F can inhibit the accumulation of HIV-1 reverse transcription products in the absence of hypermutation[J].J Biol Chem,2007,282(4):2587-2595.
猜你喜欢
河北医学(2021年10期)2021-10-27
昆明医科大学学报(2021年6期)2021-07-31
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
中国临床医学影像杂志(2019年6期)2019-08-27
生物工程学报(2019年6期)2019-07-10
生物学通报(2019年1期)2019-02-15
生物学通报(2018年12期)2018-10-10
中华实验和临床病毒学杂志(2016年3期)2016-08-09
医学研究杂志(2015年6期)2015-07-01
医学研究杂志(2015年8期)2015-06-22
