
水稻分蘖与主茎同伸叶片大小的定量关系及其对氮素的响应
2014-12-23 11:30:22魏广彬孙和平王绍华
江苏农业学报 2014年5期
魏广彬, 徐 蕊, 孙和平, 王绍华
(1.金坛市农林局,江苏 金坛213200;2.南京农业大学农学院,江苏 南京210095)
叶面积动态的模拟是作物生长模拟的基础组成部分,现有的群体叶面积模拟模型多采用干物质分配与比叶面积相结合的方法[1-5],部分研究采用有效积温法[6-10],或前期采用积温法、后期采用干物质分配与比叶面积相结合的方法[11-12]。这些方法虽然具有较高的准确性,但难以满足虚拟植物构建的需要。水稻分蘖叶片的发生与主茎出叶具有同伸关系[13],由于同伸叶片的生长发育处于同一生长期,受共同的环境影响,分蘖叶的面积与其同伸主茎叶的面积就存在一定的相关性,只要明确了分蘖叶与同伸主茎叶之间叶面积的定量关系,就可根据主茎叶的面积估算出与其同伸分蘖叶的叶面积。一些学者认为不同茎蘖上同伸的叶片最终具有相同的大小[13-14],也有一些学者提出不同的结论,认为分蘖叶与主茎同伸叶片大小的关系与分蘖叶的叶龄即分蘖叶在分蘖上的叶位有关[15]。为此,本研究重点分析分蘖叶片大小(长度、宽度)与同伸主茎叶片大小的定量关系及其对氮素的响应。
1 材料与方法
试验于2009 年在江苏省丹阳市延陵镇宝林试验农场(31o54' N,119o28' E,海拔7 m)进行。供试品种为常规粳稻南粳44(NJ44),杂交籼稻II 优107(EY107)。试验采用塑料桶露天盆栽,桶口径30 cm,桶底径22 cm,桶高30 cm。每桶装过筛细土15 kg,土壤质地为沙壤土,含有机质1.88%、全氮0.76 g/kg、碱解氮66.31 mg/kg,速效磷62.45 mg/kg,速效钾45.52 mg/kg。每个品种设4 个氮肥水平(每桶0 g、0.9 g、1.8 g、2.7 g;按1 hm2耕层土壤干质量2.25 ×106kg 计算,则分别相当于0 kg/hm2、135 kg/hm2、270 kg/hm2、405 kg/hm2),依次用N0、N1、N2、N3 表示,基追肥比为8 ︰2,每处理20 桶。5 月22 日浸种催芽,5 月25 日选“根长一粒谷,芽长半粒谷”的种子,按每桶3 粒均匀播于桶中心附近土壤表面,并用细土覆盖。待幼苗发育至2 叶1 心期进行间苗,每桶保留一株生长健壮的幼苗,同时尽量保证同品种各处理间整齐一致。基肥于播种前1 d 施入桶中,并搅拌均匀,分蘖肥于3 叶期施用,穗肥在倒4 叶期(南粳44 约16 叶期,Ⅱ优107 约15 叶期)施用。磷肥和钾肥作基肥一次施入,施用量分别为每桶0.9 g(P2O5)和1.3 g(K2O)。
测定项目及方法:每处理选10 株,每隔3 d 标记新生叶片的叶位,观测主茎、一次分蘖、二次分蘖和三次分蘖每张叶片全展时的叶长和最大叶宽(简称叶宽)。
2 结果与分析
2.1 同伸叶片叶长的相关关系
水稻叶片生长遵循明确的同伸关系,即主茎i叶抽出时,i-3 叶位的第1 叶、i-4 叶位分蘖的第2叶同时抽出[13]。同伸叶片生长期间的环境相同,因而其叶片长度间存在一定的相关性。基于此,本研究系统测定了两供试品种在不同施氮量处理下主茎、一次分蘖、二次分蘖及三次分蘖各叶片的叶长,并计算出分蘖各叶位叶片与主茎同伸叶片叶长的比值(以下简称叶长比)。现列举与主茎第8 叶和第10 叶同伸的各分蘖叶片的叶长及叶长比(表1,表2),分析叶长与施氮量的关系及叶长比与蘖位和叶位的关系。
分析表1 和表2 中主茎叶长与分蘖叶长的关系,发现随着施氮水平的提高,主茎叶叶长变大,而与该主茎叶同时抽出的分蘖叶的长度也明显增加,同伸叶片的叶长表现出一定程度的正相关性。例如,南粳44 主茎第8 叶同伸的分蘖叶片有0/1分蘖第5 叶、0/2 分蘖第4 叶、0/3 分蘖第3 叶、0/4分蘖第2 叶、0/5 分蘖第1 叶、0/1/1 分蘖第2 叶、0/1/2 分蘖第1 叶和0/2/1 分蘖第1 叶等8 张,在N3 处理下主茎第8 叶的叶长较N0 增加了31.1%,而与该主茎叶同伸的8 张分蘖叶片的叶长也较N0 处理对应叶片分别增加了30.8%、32.9%、26.4%、32.2%、34.6%、35.5%、42.7%和49.9%,分蘖叶长的增加幅度与主茎第8 叶较接近。南粳44 主茎第10 叶、Ⅱ优107 主茎第8 叶和10 叶同伸的分蘖叶片也表现出类似情况,表明主茎叶叶长的增长与分蘖叶长的增长同步性是一种较为普遍的现象,可以依据主茎叶的叶长预测分蘖叶长。
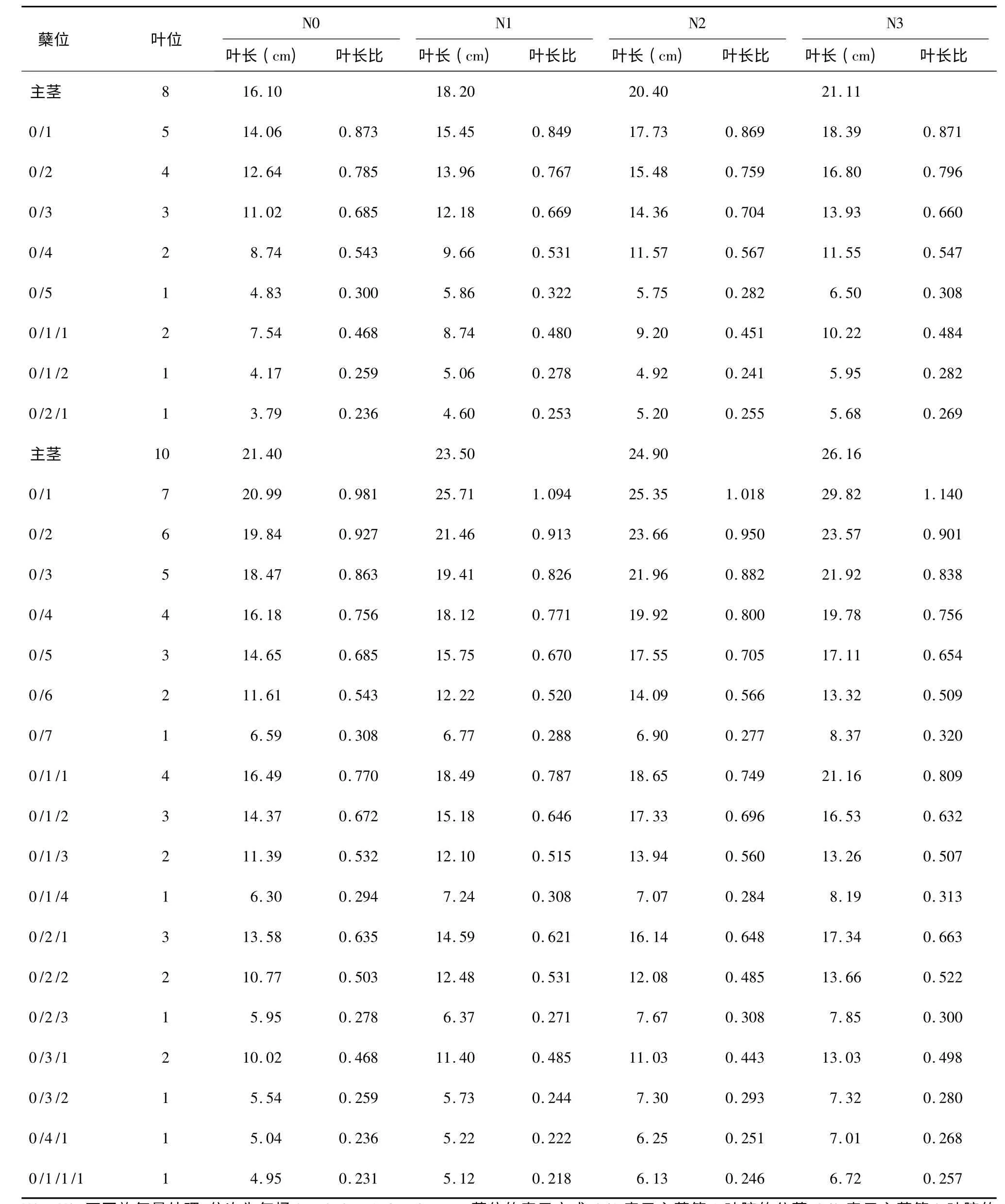
表1 南粳44 与主茎第8 和第10 叶同伸分蘖叶片的叶长和叶长比Table 1 Length and length ratio of leaves synchronously emerged with 8th and 10th leaves on main stem of Nanjing44
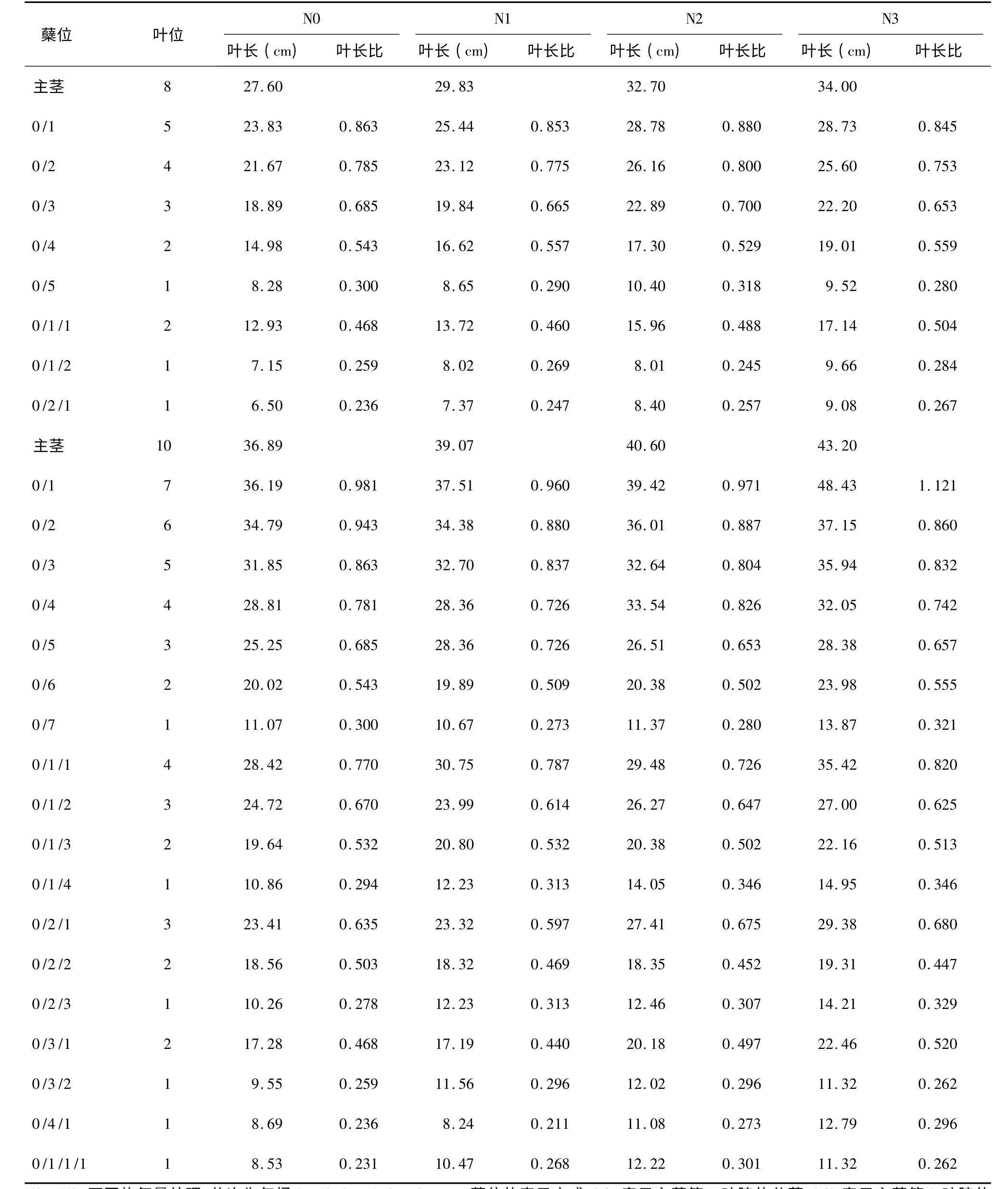
表2 II 优107 与主茎第8 和第10 叶同伸分蘖叶片的叶长和叶长比Table 2 Length and length ratio of leaves synchronously emerged with 8th and 10th leaves on main stem of Eryou107
分析分蘖叶叶长与同伸主茎叶叶长的比例关系,发现其变化具有一定的规律性。以南粳44 N0处理主茎第10 叶同伸的分蘖叶片为例,该叶同伸的分蘖叶片有18 张。其中,0/1、0/2、0/3、0/4、0/5、0/6、0/7 等7 个分蘖均为一级分蘖,在这7 个分蘖上分别为第7、6、5、4、3、2、1 叶位的7 张叶片与主茎第10 叶同伸,这7 张叶的叶长比分别为0.981、0.927、0.863、0.756、0.685、0.543、0.308,叶片所在分蘖上的叶位从7 下降到1,叶长比从0.981 下降到0.308,表现为随叶位降低叶长比降低。再看0/1/1、0/1/2、0/1/3、0/1/4 等4 个二级分蘖,该4个分蘖上分别是第4、3、2、1 叶位的叶片与主茎第10 叶同伸,这4 张叶的叶长比分别为0.770、0.670、0.532、0.294,也表现出随叶位降低叶长比降低的趋势。进一步比较叶位相同的一级分蘖与二级分蘖,0/4 和0/1/1 的叶位均为4,叶长比前者为0.756,后者为0.770,二者十分接近;0/5 与0/1/2 的叶位均为3,叶长比分别为0.685 和0.670,也十分接近;0/6和0/1/3 的叶位同为2,叶长比分别为0.543 和0.532,很接近;0/7 和0/1/4 的叶位同为1,叶长比分别为0.308 和0.294,仍然很接近。进一步比较叶位为1 的6 个分蘖,即0/7、0/1/4、0/2/3、0/3/2、0/4/1 和0/1/1/1,他们的叶长比分别为0.300、0.294、0.278、0.259、0.236 和0.231,这6 个分蘖有一级分蘖,有二级分蘖,也有三级分蘖,总体上看叶长比有随分蘖级别降低而降低的趋势,但不十分明显,对表1 和表2 中其他数据进行仔细分析,都出现了类似的结果,表明同龄分蘖的同伸叶片具有相近的叶长,叶长比主要与叶片在分蘖上的叶位有关,而与分蘖的位置关系不密切。
进一步分析叶长比与对应叶片在分蘖上叶位的定量关系(图1),可以看出,分蘖叶片与主茎同伸叶片的叶长比随蘖龄提高而增大,约在蘖龄达到7 时,叶长比达到1,氮肥和品种的不同对这一关系没有明显的影响。同伸叶片叶长之间的这种规律性关系可用式(1)来定量表达。
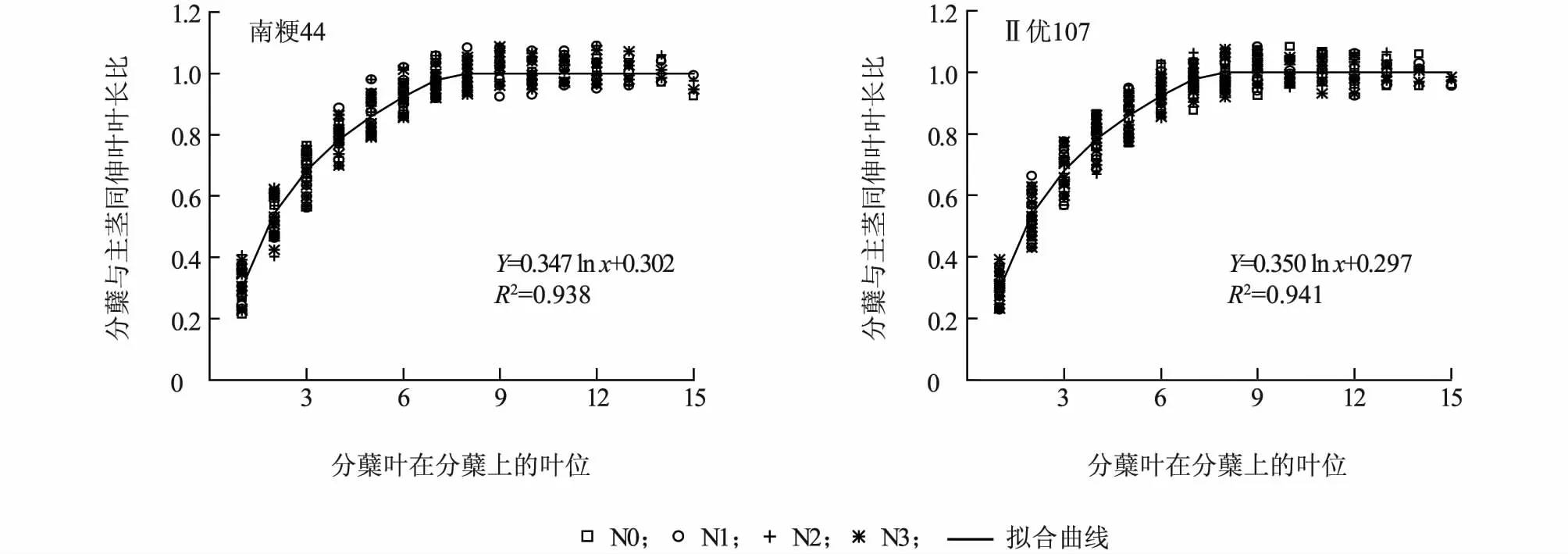
图1 叶长比随分蘖叶在分蘖上的叶位变化(数据为主茎4 ~18 叶期)Fig.1 Length ratio of synchronously emerged leaf on main stem and on tiller changed with tiller-leaf position (data from leaf age 4 to 18)
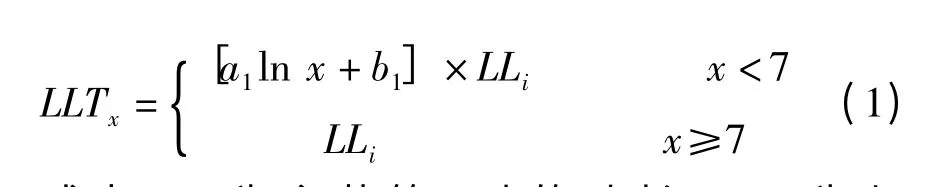
式中,LLi为主茎第i叶的叶长,LLTx为与主茎第i叶同伸叶位为x的分蘖叶的叶长,a1、b1为系数。利用图1 的数据,分品种拟合式(1)中 的a1、b1,南 粳4 4 分 别 为0.3 4 7 和0.3 0 2,R2= 0.9 3 8,Ⅱ优1 0 7 分别为0.3 5 0和0.2 9 7,R2= 0.9 4 1,两品种的模型系数较为接近。
2.2 同伸叶片叶宽的相关关系
分析表3 和表4 中主茎叶宽与分蘖叶宽的关系,发现随着施氮水平提高,主茎叶叶宽没有明显的变化,而与该主茎叶同时抽出的分蘖叶的宽度也没有明显的变化。
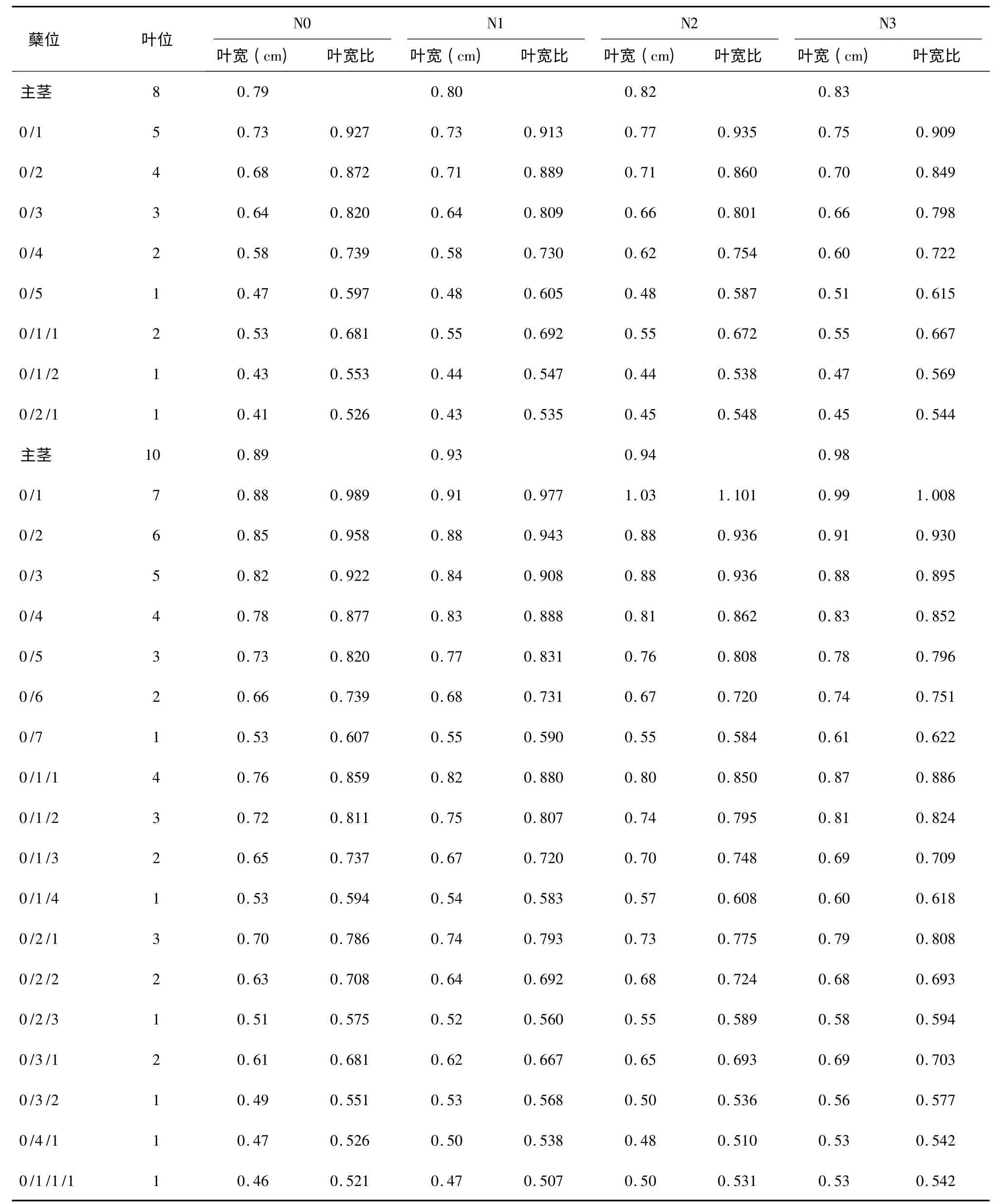
表3 南粳44 与主茎第8、第10 叶同伸分蘖叶片叶宽和叶宽比Table 3 Width and width ratio of leaves synchronously emerged with 8th and 10th leaves on main stem of Nanjing44
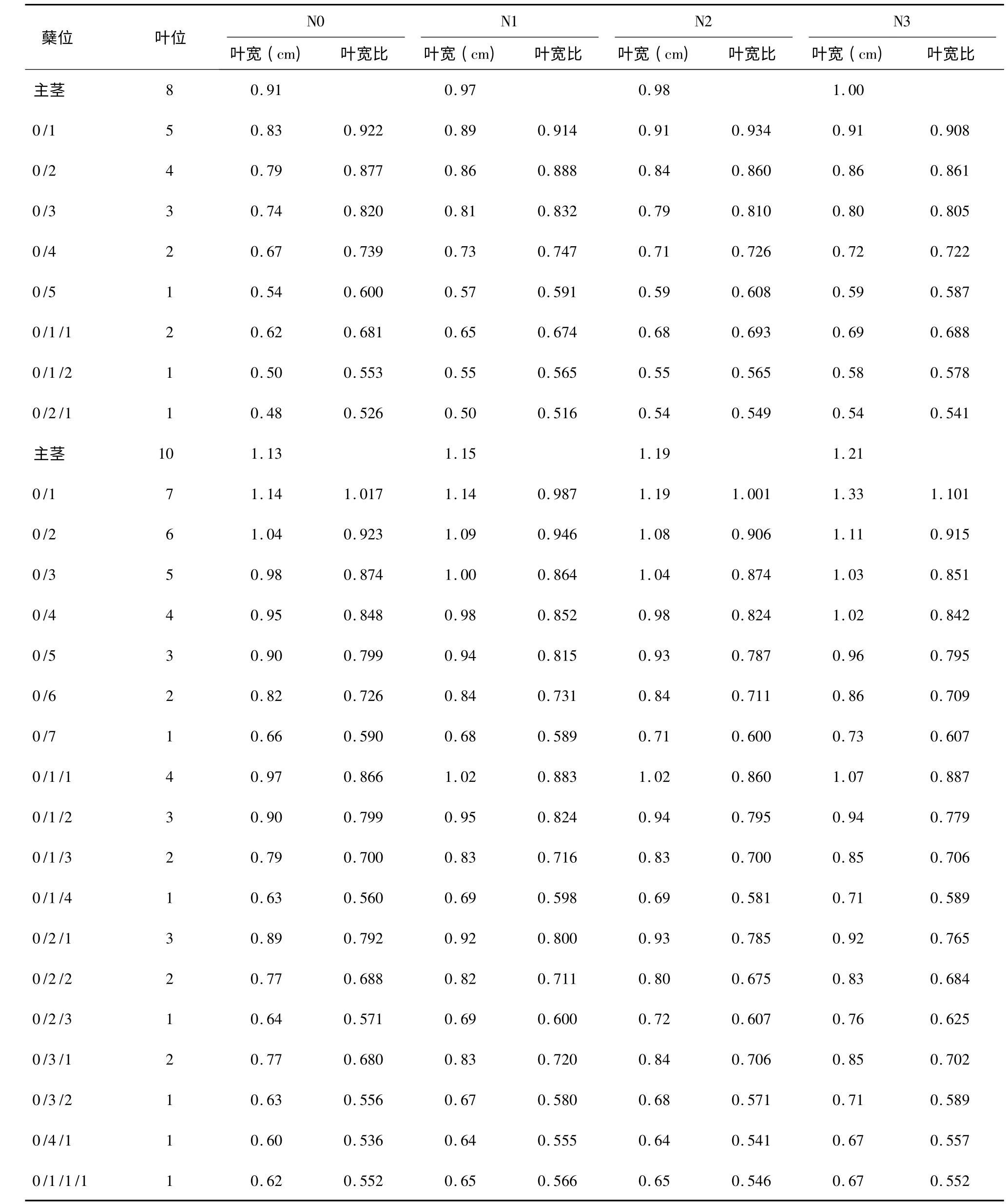
表4 II 优107 与主茎第8、第10 叶同伸分蘖叶片叶宽和叶宽比Table 4 Width and width ratio of leaves synchronously emerged with 8th and 10th leaves on main stem of Eryou107
分析分蘖叶叶宽与同伸主茎叶叶宽的比例关系,发现其变化也具有一定的规律性。同样以南粳44 N0 处理与主茎第10 叶同伸的分蘖叶片为例,0/1、0/2、0/3、0/4、0/5、0/6、0/7 等7 个一级分蘖上与主茎第10 叶同伸叶片的叶宽比分别为0.989、0.958、0.922、0.877、0.820、0.739、0.607,叶片所在分蘖上的叶位从7 下降到1,叶宽比从0.989 下降到0.607,表现为随叶位降低叶宽比降低。再看0/1/1、0/1/2、0/1/3、0/1/4 等4 个二级分蘖,该4个分蘖上分别是第4、3、2、1 叶位的叶片与主茎第10 叶同伸,这4 张叶的叶宽比分别为0.859、0.811、0.737、0.594,也表现出了随叶位降低叶宽比降低的趋势。进一步比较叶位相同的一级分蘖与二级分蘖,0/4 和0/1/1 的叶位均为4,叶宽比前者为0.877,后者为0.859,二者十分接近;0/5 与0/1/2的叶位均为3,叶宽比分别为0.820 和0.811,也十分接近;0/6 和0/1/3 的叶位同为2,叶宽比分别为0.739 和0.737,很接近;0/7 和0/1/4 的叶位同为1,叶宽比分别为0.607 和0.594,仍然很接近。进一步比较叶位为1 的6 个分蘖,即0/7、0/1/4、0/2/3、0/3/2、0/4/1 和0/1/1/1,他们的叶宽比分别为0.607、0.594、0.575、0.551、0.526、0.521,这6 个分蘖有一级分蘖,有二级分蘖,也有三级分蘖,总体上看叶宽比有随分蘖级别降低而降低的趋势,但不十分明显。对表3 和表4 中其他数据进行仔细分析,都出现了类似的结果,表明同龄分蘖的同伸叶片具有相近的叶宽,叶宽比主要与叶片在分蘖上的叶位有关,而与分蘖的位置关系不密切。
进一步分析叶宽比与对应叶片在分蘖上的叶位定量关系(图2),可以看出,分蘖与主茎同伸叶片的叶宽比随分蘖叶在分蘖上叶位的提高而增大,约在分蘖叶叶位达到7 时,叶宽比达到1。氮肥和品种的不同对这一关系没有明显的影响。同伸叶片叶宽之间的这种规律性关系可用式(2)来定量表达。

图2 叶宽比随分蘖叶在分蘖上的叶位变化(数据为主茎4 ~18 叶期)Fig.2 Width ratio of the synchronously emerged leaves on main stem and on tiller changed with tiller-leaf position (data from leaf age 4 to 18)
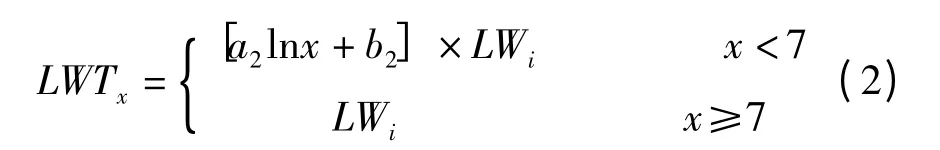
式中,LWi为主茎第i叶的叶宽,LWTx为与主茎第i叶同伸叶位为x的分蘖叶叶宽,a2、b2为系数。利用图2 数据,分品种拟合式(2)中的系数a2、b2,南粳44 分别为0.209 和0.584,R2= 0.886;Ⅱ优107 分别为0.198 和0.601,R2= 0.849。两品种的模型系数较为接近。
3 讨论
有学者提出,不同茎蘖上同伸的叶片最终具有相同的大小[13-14],但本研究的结果却不支持这一结论。分蘖上第1 叶的全展叶长大约相当于与其同伸的主茎叶叶长的30%,之后随着分蘖上叶位的提高,分蘖与主茎的同伸叶长比逐渐增加;当分蘖叶片数达到7 时,其比值近似为1,即分蘖与主茎同伸叶片长度趋于一致。这与石春林等[15]的研究结果一致。分蘖叶叶宽与主茎同伸叶叶宽之间也有与叶长相类似的定量关系,这使得通过主茎叶片大小可以估计分蘖叶片大小,进而估计单株和冠层的叶面积。由于本模型能够模拟任意单个分蘖叶片的大小,与叶片伸长过程模型[16]相结合,可以为虚拟植物的构建提供一定的支持[17-18]。
[1] 刘 岩,陆建飞,曹宏鑫,等.基于生物量的水稻叶片主要几何属性模型研究[J].中国农业科学,2009,42(11):4093-4099.
[2] CHARLES-EDWARDS D A,FISHER M J.A physiological approach to the analysis of crop growth data I.Theoretical considerations[J].Annals of Botany,1980,46:413-423.
[3] VAN KEULEN H,SELIGMAN N G.Simulation of water use,nitrogen nutrition and growth of spring wheat crop[M].Wageningen:Simulation Monographs Pudoc,1987.
[4] PENNING DE VRIES F W T,JANSEN D M,TEN BERGE H F M,et al.Simulation of ecophysiological processes of growth in several annual crops[M].Wageningen,The Netherlands:Pudoc and Los Banos,Philippines:International Rice Research Institute,1989.
[5] GOUDRIAAN J,VAN LAAR H H.Modeling potential crop growth processes[M].Dordrecht,The Netherlands:Kluwer Academic Publishers,1994.
[6] HUNT R,PARSONS I T.A computer program for deriving growth functions in plant growth analysis[J].J Appl Ecol,1974,11:297-307.
[7] DALE R F,COELHO D T,GALLO K P.Prediction of daily green leaf area index for corn[J].Agron J,1980,72:999-1005.
[8] JONES C A,KINIRY J R.CERES-maize:a simulation model of maize growth and development[M].College Station,TX:Texas A&M University Press,1986.
[9] HAMMER G L,CARBERRY P S,MUCHOW R C.Modeling genotypic and environmental control of leaf area dynamics in grain sorghum.I.Whole plant level[J].Field Crops Res,1993,33:293-310.
[10] JAMIESON P D,SEMENOV M A,BROOKING I R,et al.Sirius:a mechanistic model of wheat response to environmental variation[J].Eur J Agron,1998,8:161-179.
[11] KROPFF M J.Modeling the effects of weeds on crop production[J].Weed Res,1988,28:465-471.
[12] SPITTERS C J T,VAN KEULEN H,VAN KRAALINGEN D W G.A simple and universal crop growth simulator:SUCROS87[M]//RABBINGE R,WARD S A,VAN LAAR H H.Simulation and systems management in crop protection.Wageningen:Pudoc,1989:147-181.
[13] KATAYAMA T.Studies on tillering of rice,wheat and barley[M].Tokyo:Yokendo,1951.
[14] KEMP D R.Comparison of growth rates and sugar and protein concentrations of the extension zone of main shoot and tiller leaves of wheat[J].Journal of Experimental Botany,1981,32:151-158.
[15] 石春林,朱 艳,曹卫星.水稻叶片几何参数的模拟分析[J].中国农业科学,2006,39(5):910-915.
[16] 常丽英,顾东祥,张文宇,等.水稻叶片伸长过程的模拟模型[J].作物学报,2008,34(2):311-317.
[17] 孙爱珍,杨红云,何火娇,等.水稻叶片三维可视化建模[J].安徽农业科学,2008,36(4):1320-1321.
[18] 杨红云,罗 威,何火娇,等.水稻叶片形态三维建模与计算机模拟[J].农机化研究,2008 (12):33-35.
猜你喜欢
农业与技术(2023年11期)2023-06-19 02:28:18
干旱地区农业研究(2022年4期)2022-07-28 05:37:08
四川蚕业(2022年1期)2022-06-06 02:03:42
新疆农业科学(2020年1期)2020-02-14 03:22:46
亚热带植物科学(2019年2期)2019-07-22 13:16:12
江苏农业学报(2018年5期)2018-09-10 22:36:44
江苏农业科学(2016年10期)2017-02-05 14:45:09
西南农业学报(2016年5期)2016-05-17 05:42:39
家家乐·上半月(2013年6期)2013-04-29 00:44:03
