
饲料中玉米赤霉烯酮的生物降解
2014-12-20 07:37计成
动物营养学报 2014年10期
计 成
(中国农业大学动物科学技术学院,动物营养学国家重点实验室,北京 100193)
霉菌毒素(mycotoxin)是由曲霉菌属(Aspergillus spp.)、镰刀菌属(Fusarium spp.)和青霉菌属(Penicillium spp.)等真菌在生长过程中产生的有毒的次级代谢产物[1]。目前已经报道,大约有300多种霉菌代谢产物对人类和动物具有毒害作用,其中,黄曲霉毒素(aflatoxin,AFT)、单端孢霉烯族毒素[trichothecenes,如 T-2毒素和呕吐毒素(DON)]、玉米赤霉烯酮(zearalenone,ZEA)、赭曲霉毒素(ochratoxin A,OTA)及伏马毒素(fumonisins)等是对畜牧业威胁最大的几种霉菌毒素[2]。ZEA,又称F-2毒素,是一种非类固醇类的、具有类雌激素作用的真菌毒素,主要由禾谷镰刀菌、三线镰刀菌、粉红镰刀菌和雪腐镰刀菌等多种镰刀菌产生[3]。ZEA的化学名为6-(10-羟基 -6-氧基-十一碳烯基)-β-雷锁酸内酯,分子式是C18H22O5,相对分子质量为318。ZEA紫外线光谱的最大吸收波长有236、274和316 nm,红外线光谱的最大吸收波长为970 nm。ZEA的熔点为161~163℃,ZEA不溶于水、二氧化碳(CO2)和四氯甲烷(CCl4),溶于碱性水溶液和甲醇、乙腈、乙醚、苯、二氯甲烷、乙醇和乙酸乙酯等有机溶剂,微溶于石油醚。
1 ZEA污染现状
据联合国粮农组织(FAO)的统计结果可知,每年全世界有25%的谷物受各种各样的霉菌毒素污染,其中2%的粮食由于霉菌毒素污染而不能食用,仅畜禽食用霉菌毒素污染的饲料使美国畜牧业每年遭受近10亿美元的经济损失,而对全世界造成的经济损失,每年达数千亿美元,而且这种趋势有增无减[4]。而镰刀菌毒素是造成全世界谷物经济损失最大的一类污染性霉菌毒素。镰刀菌产生毒素的过程主要发生于谷物收获之前,但若在收仓存放前没有经过适当的干燥等方法处理,镰刀菌也会在谷物存放阶段大量产生毒素。ZEA广泛存在于霉变的玉米、小麦、大麦、燕麦、高粱等谷物以及奶产品中,是世界上污染范围最广的一种镰刀菌毒素。Elisabeth等[5]测定了2004—2011年世界范围内共计13 578个饲料及饲料原料样品中ZEA含量,结果表明,样品中ZEA检出率为36%,平均含量为101μg/kg,最高含量高达26 728μg/kg。王金勇等[6]对2012年从我国采集的841个饲料原料和饲料样品进行检测,检测结果表明,我国的饲料原料和饲料均受到不同程度的ZEA污染,其中,猪料、禽料、玉米和干酒糟及其可溶物(DDGS)的呈阳性的ZEA平均含量分别为446、977、658 和 659 μg/kg。雷元培等[7]2011年对北京地区131个饲料原料和饲料样品抽样检测发现,玉米、麸皮、DDGS和猪全价饲料中ZEA阳性检出率均达100.00%,豆粕中ZEA阳性检出率达54.45%。其中,DDGS污染情况最严重,ZEA最高含量达5 957.87μg/kg,平均含量达(882.68 ±146.40)μg/kg。由此可见,我国的ZEA污染情况严重。
2 动物ZEA中毒症
猪是对ZEA最敏感的动物,特别是青春期前的母猪,其对ZEA的敏感性更强。ZEA中毒可引起猪性早熟、母猪发情周期延长、假发情、卵巢萎缩、持久黄体、母猪假孕和流产、窝产仔数减少或产弱仔等症状[8-9]。Price等[10]报道,高剂量的ZEA(50~100 mg/kg)可对母猪排卵、受孕、胚胎定植、胎儿发育和新生儿的生活力造成显著的影响,并且会出现明显的外阴阴道炎、阴道和直肠垂脱,同时,母猪ZEA中毒出现连续动情会导致不育、假孕、卵巢畸形和流产等一系列的生殖障碍[11-12]。母猪或仔猪接触ZEA时的阶段不同、接触ZEA的持续时间和剂量不同,都将会导致母猪或仔猪出现不同程度的雌激素中毒症状(如轻微中毒时出现阴户和乳头红肿、子宫和阴道肿大,严重时还会出现阴道或直肠脱垂)[13-15]。Collins等[16]研究发现,在怀孕母鼠第6~19天的饲粮中添加8 mg/kg的ZEA可引起胎儿发育延迟、骨骼肌发育迟钝以及初生小鼠的成活率降低。同时,大量研究证明,ZEA和 α-玉米赤霉烯醇(αzearalenol,α-ZOL)对雄性动物的生殖发育也具有一定的负面影响。在雄性动物体内,ZEA和α-ZOL能够通过影响雄性动物睾酮的分泌,进而影响成年雄性动物精子的发生、成熟和性功能[17]。Young等[18]报道,公猪采食 ZEA含量为9 mg/kg的饲粮,公猪的精液质量明显下降(如精液的总量、总活精数以及凝胶自由体积)。Wilson等[19]报道,放牧公山羊耳埋36 mg ZEA 90 d后,其攻击性行为、睾丸生长和性欲活动都明显减少。同时,体外研究表明,于精液中添加ZEA和α-ZOL,精子生存力和生活力会显著降低,公猪精子的生育能力下降[20-21]。图1为ZEA引起的母猪外阴红肿和脱落症状,图2为ZEA中毒引起的子宫肿大症状。

图1 ZEA引起的外阴红肿、阴道和直肠脱垂Fig.1 Swell of the vulva and prolapse of the vagina and rectum caused by ZEA
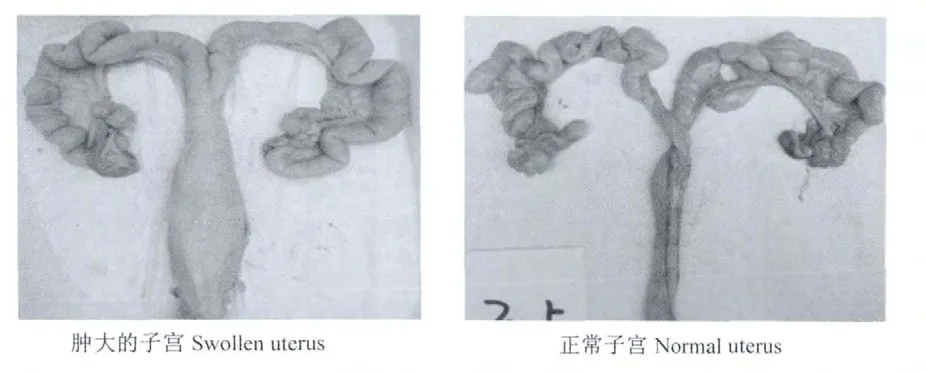
图2 ZEA对母猪子宫的影响Fig.2 Effects of ZEA on uterus of sows
3 ZEA毒理作用机制
ZEA和其代谢产物都具有雌激素活性作用,主要是因为它们能够与胞膜上的雌激素受体(estrogen receptor,ER)结合,使 ER发生构型改变,ZEA与ER形成的复合物被转移至核内将会和雌激素效应元件结合,从而对靶基因的转录与蛋白质合成进行调节[22]。Kuiper-Goodman 等[9]研究结果表明,在ZEA和其最主要的几种代谢产物之中,α-ZOL与ER的结合力最高,ZEA及其代谢产物与ER的相对结合力高低顺序依次是α-ZOL>α-玉米赤霉醇(α-ZAL)>β-玉米赤霉烯醇(β-ZOL)>ZEA>β-玉米赤霉醇(β-ZAL)。ER有1个具有反式激活功能的高度变异的N末端区,1个负责特定的DNA结合、二聚化及核定位的高度保守的中央区,以及1个参与配体结合及配体依赖的反式激活功能的C末端区[23]。ER与雌激素分子之间的亲和力非常高,能够通过受体特异性进行结合。Dees等[24]研究发现,ZEA在动物机体内与ER结合后,被转移到细胞核中,进而与细胞核内的染色质相结合,通过调节细胞内的基因转录和蛋白质合成,从而对细胞的分裂和生长产生影响,加速乳腺癌细胞MCF-7进入细胞周期,促进MCF-7细胞的增殖。
ZEA可导致细胞核内DNA和染色体的损伤,这可能是ZEA引起动物机体出现一系列中毒症状的原因之一。据 Abid-Essefi等[25]报道,ZEA 在1~15μg/mL细胞悬液浓度范围内可导致细胞DNA发生损伤,且具有明显的剂量反应关系。Ouanes等[26]研究发现,当 ZEA 的剂量为 2~20 mg/kg时,可诱导细胞核内染色体出现不同程度和不同类型的变异、畸形,该剂量达到半数致死量的0.4% ~4.0%。但相同剂量ZEA分次连续注射和一次性注射导致的染色体损伤程度不同,分次连续注射对染色体的损伤比一次性注射更加严重。
4 ZEA生物降解国内外研究进展
ZEA污染范围广泛,危害程度严重,因此,有效控制和解决其对粮食和饲料的污染,对改善动物生产性能和提高人类食品安全有着非常重要的意义。目前去除饲料中ZEA的方法主要包括物理法、化学法和生物降解法。传统的物理和化学方法虽然能部分地去除饲料和饲料原料中的霉菌毒素,但这些方法都有着效果不稳定、营养成分损失大、影响饲料适口性和难以规模化生产应用等不可避免的缺点,而且在饲料和饲料原料中添加化学试剂也会将一些不确定的危害因素引入动物生产环节中[27-28]。近年来ZEA生物降解方法越来越受到重视。ZEA的生物降解方法主要是指微生物、植物或其代谢时产生的酶与ZEA发生作用并使ZEA分子结构中毒性基团被破坏从而生成无毒代谢产物的过程[29]。由于微生物及其生物酶降解ZEA的方法能够彻底去除毒素,同时,该方法的专一性强、对饲料无污染、不会影响饲料营养价值,因此,该方法在动物霉变饲料和饲料原料中的应用具有广阔的发展前景。
4.1 降解ZEA的真菌
研究报道黏帚霉(Gliocladium sp.)属中的粉红黏帚霉(Gliocladium roseum)可通过一系列脱羧反应使大部分ZEA开环生成无毒化合物[30-32]。Varga等[33]报道,根霉菌属(Rhizopus sp.)中的米根霉(Rhizopus oryzae)、葡枝根霉(Rhizopus stolonifer)和小孢根霉(Rhizopus microsporus)能将ZEA完全降解。雅致枝霉(Thamnidium elegans)和贝勒被毛霉(Mucor bainieri)使ZEA成ZEA-4-O-β-葡糖苷[34],龟裂链霉菌(Streptomyces rimosus)和班尼小克银汉霉(Cunninghamella bainieri)将ZEA分别转化成8’-羟基-ZEA和2,4-二甲氧基-ZEA[35]。研究报道,少根根霉(Rhizopus arrhizus)[36]和 黑 曲 霉 (Aspergillus section Nigri)[37]能将ZEA上C-4位的羟基氧化为硫酸根,从而将ZEA转化为低毒的硫酸-ZEA。Molnar等[38]报道解毒毛孢酵母(Trichosporon mycotoxinivorans)可在24 h内将ZEA降解成二氧化碳或者无荧光和无紫外吸收的代谢物,其代谢物中没有检测到 α-ZOL、β-ZOL或ZEA的存在,并且认为该菌可用于去除动物饲粮中的霉菌毒素。Vekiru等[39]使用液相色谱串联质谱法(LC-MS/MS)、液相色谱二极管阵列(DAD)以及核磁共振(NMR)方法对解毒毛孢酵母生物降解ZEA的代谢产物进行分析,结果发现ZEA分子中的大环被打开,代谢生成了非雌激素化合物。
4.2 降解ZEA的细菌
目前,关于细菌降解ZEA的研究报道较少,黏质塞氏杆菌(Serratia marcescens)[40]、假单胞菌(Pseudomonas sp.)[41-42]、动球杆菌(Planococcus sp.)[43]、来源于土壤的细菌混合物、猪肠道微生物混合物[44]能够对 ZEA进行生物降解。同时,Yi等[45]研究发现,地衣芽孢杆菌(Bacillus licheniformis)与ZEA孵育36 h后能够降解95.8%的ZEA,同时该菌株还具有木聚糖酶、纤维素酶及蛋白酶活性,其在动物饲粮中具有一定的应用前景。Tinyiro等[46-47]分离到2株具有降解ZEA作用的芽孢杆菌,即枯草芽孢杆菌(Bacillus subtilis)168和纳豆芽孢杆菌(Bacillus natto)CICC 24640,30℃供氧条件下与20μg/L ZEA作用24 h后,能够分别降解81%和100%的 ZEA。除此之外,Cho等[48]从土壤中筛选得到1株枯草芽孢杆菌,其液态发酵24 h后能够降解99%的ZEA,固态发酵48 h后能够降解95%的ZEA。
中国农业大学动物营养学国家重点实验室对细菌生物降解ZEA及其降解机理进行了研究,并取得很大进展。首先,Lei等[49]从72个自然界样品及1 088株实验室保藏菌株中,筛选出1株能够高效降解ZEA的枯草芽孢杆菌ANSB01G,其在液体培养基中对ZEA的降解率为88.65%,并且该菌能够降解自然霉变玉米、DDGS和猪全价配合饲料中 84.58%、66.34% 以及 83.04% 的 ZEA。该菌对沙门氏菌、大肠杆菌以及金黄色葡萄球菌具有很好的抑菌活性,同时在高温、胃酸和胆盐条件下具有较高的存活率和活性。该菌株已经申请国家发明专利得到保护(专利号 ZL 201010620651.2)。其次,雷元培[50]通过对枯草芽孢杆菌ANSB01G降解ZEA特性的研究,发现该菌降解ZEA的活性物质为一种细菌所产的胞外酶。通过用硫酸铵沉淀,离子交换层析、疏水层析和分子筛层析方法从菌株ANSB01G发酵液中分离出降解ZEA的活性蛋白,该活性物质大致相对分子质量范围为 20.1×103~44.3×103。本实验室还通过高效液相色谱-二级质谱联用(HPLCMS)测定ZEA降解产物的分子结构,推测菌株ANSB01G降解ZEA的作用机制,即首先ZEA分子中酚羟基与谷氨酸的γ-羧基结合,然后分子结构中内酯环水解、脱羧、还原羰基,最后脱水,该解毒机制是一种新的ZEA代谢途径[50]。
4.3 降解ZEA的酶及酶制剂
2002 年,Takahashi-Ando 等[51]从粉红黏帚霉IFO7063中分离纯化得到能够生物降解ZEA的内酯水解酶(lactonohydrolase),该酶能够将ZEA转化成为无毒的代谢物排出体外。同时对编码降解ZEA的基因(zhd101)进行了克隆,并将该基因成功转入到大肠杆菌(Escherichia coli)和酿酒酵母(Saccharomyces cerevisae)进行了表达,外源表达该片段基因的大肠杆菌在24 h内可将培养基中的ZEA、α-ZOL和β-ZOL全部降解成无毒的代谢产物[52]。此外,Igawa等[53]将 zhd101 基因导入到玉米中得到转基因玉米,使用该转基因玉米可以在48 h内将溶液中90%的ZEA去除,每克转基因玉米种子可以去除16.9μg的ZEA。
5 小结
自从1966年发现ZEA以来,世界各国的科研工作者对ZEA的研究从未间断过,主要包括ZEA的合成、代谢、毒理、检测、降解和防治措施等方面。在ZEA脱毒方面,由于传统的物理和化学方法在应用上存在弊端,生物降解ZEA备受关注,但目前ZEA生物降解技术研究尚存在一些不足之处,还需进一步深入研究。目前虽然在降解ZEA的微生物筛选方面取得了一定的进展,但大部分研究报道没有探明其作用机制,不能确定ZEA浓度的降低是因为微生物的物理性吸附作用还是生物酶的生物降解作用。由于微生物的吸附作用是可逆的,毒素分子并没有消失,随着时间的延长,部分毒素会重新释放,因此,并不是很理想的解毒方法;其次,被证明能生物降解ZEA的微生物菌株数量和种类相对较少,特别是能直接应用于饲料中降解ZEA的益生菌的报道更少。因此,需要进一步筛选高效降解ZEA的益生菌,并对其生物降解机理和代谢途径进行深入研究和探讨。
筛选降解饲料ZEA的益生菌是饲料工业未来研究发展的一个方向。益生菌可以直接应用于饲料中,降解饲料ZEA的同时还具有益生性,不但生产工艺简单,而且可降低生产者和养殖者的成本,易于规模化推广和应用。另外,运用生物化学、分子生物学及基因工程等手段,将ZEA降解酶基因转化到表达宿主细胞中,大量表达和生产ZEA降解酶,开发高纯度、高活性的新型降解酶饲料添加剂,也是饲料工业研究发展的方向。
[1] HUSSEIN H S,BRASEL J M.Toxicity,metabolism,and impact of mycotoxins on humans and animals[J].Toxicology,2001,167(2):101-134.
[2] GLENN A E.Mycotoxigenic Fusarium species in animal feed[J].Animal Feed Science and Technology,2007,137(3/4):213-240.
[3] RICHARD J L,PAYNE G A,DESJARDINS A E,et al.Mycotoxins:risks in plant,animal and human systems[J].CAST Task Force Report,2003,139:101-103.
[4] ZINEDINE A,SORIANO J M,MOLTÓ J C,et al.Review on the toxicity,occurrence,metabolism,detoxification,regulations and intake of zearalenone:an oestrogenic mycotoxin[J].Food and Chemical Toxicology,2007,45(1):1-18.
[5] ELISABETH S,KARIN N,INÊS R,et al.Mycotoxin occurrence in feed and feed raw materials worldwide:long-term analysis with special focus on Europe and Asia[J].Journal of the Science of Food and Agriculture,2013,93(12):2892-2899.
[6] 王金勇,刘颖莉,关舒.2012年中国饲料和原料霉菌毒素检测报告[J].中国畜牧杂志,2013,49(4):29-34.
[7] 雷元培,马秋刚,谢实勇,等.抽样调查北京地区猪场饲料及饲料原料玉米赤霉烯酮污染状况[J].动物营养学报,2012,24(5):905-910.
[8] D’MELLO J P F,PLACINTA C M,MACDONALD A M C.Fusarium mycotoxins:a review of global implications for animal health,welfare and productivity[J].Animal Feed Science and Technology,1999,80(3/4):183-205.
[9] KUIPER-GOODMAN T,SCOTT P M,WATANABE H.Risk assessment of the mycotoxin zearalenone[J].Regulatory Toxicology and Pharmacology,1987,7(3):253-306.
[10] PRICE W D,LOVELL R A,MCCHESNEY D G.Naturally occurring toxins in feedstuffs:Center for Veterinary Medicine Perspective[J].Journal of Animal Science,1993,71(9):2556-2562.
[11] EDWARDS S,CANTLEY T C,ROTTINGHAUS G E,et al.The effects of zearalenone on reproduction in swine.Ⅰ.The relationship between ingested zearalenone dose and anestrus in non-pregnant,sexually mature gilts[J].Theriogenology,1987,28(1):43-49.
[12] OSWEILER G D.Occurrence and clinical manifestations of trichothecene toxicoses and zearalenone toxicoses[J].Diagnosis of Mycotoxicoses,1986,33:31-42.
[13] BIEHL M L,PRELUSKY D B,KORITZ G D,et al.Biliary excretion and enterohepatic cycling of zearalenone in immature pigs[J].Toxicology and Applied Pharmacology,1993,121(1):152-159.
[14] JIANG SZ,YANG Z B,YANG W R,et al.Effects of purified zearalenone on growth performance,organ size,serum metabolites,and oxidative stress in postweaning gilts[J].Journal of Animal Science,2011,89(10):3008-3015.
[15] KORDIC'B,PRIBIC'EVIC'S,MUNTANOLA-CVETKOVIC'M,et al.Experimental study of the effects of known quantities of zearalenone on swine reproduction[J].Journal of Environmental Pathology,Toxicology and Oncology:Official Organ of the International Society for Environmental Toxicology and Cancer,1992,11(2):53-55.
[16] COLLINS T F X,SPRANDO R L,BLACK T N,et al.Effects of zearalenone on in utero development in rats[J].Food and Chemical Toxicology,2006,44(9):1455-1465.
[17] RUZSÁS C,BIRÓ-GOSZTONYI M,WÖLLER L,et al.Effect of the fungal toxin(zearalenone)on the reproductive system and fertility of male and female rats[J].Acta Biologica Academiae Scientiarum Hungaricae,1979,30(4):335-345.
[18] YOUNG L G,KING G J.Low concentrations of zearalenone in diets of boars for a prolonged period of time[J].Journal of Animal Science,1986,63(4):1197-1200.
[19] WILSON T W,NEUENDORFF D A,LEWISA W,et al.Effect of zeranol or melengestrol acetate(MGA)on testicular and antler development and aggression in farmed fallow bucks[J].Journal of Animal Science,2002,80(6):1433-1441.
[20] TSAKMAKIDIS I A,LYMBEROPOULOS A G,ALEXOPOULOS C,et al.In vitro effect of zearalenone andα-zearalenol on boar sperm characteristics and acrosome reaction[J].Reproduction in Domestic Animals,2006,41(5):394-401.
[21] RAJKOVIC A,UYTTENDAELE M,DEBEVERE J.Computer aided boar semen motility analysis for cereulide detection in different food matrices[J].International Journal of Food Microbiology,2007,114(1):92-99.
[22] RILEY R T.Mechanistic interactions of mycotoxins:theoretical considerations[M]//SINHA K H,BHAT-NAGAR D.Mycotoxins in Agriculture and Food Safety.New York:Marcel Dekker,Inc.,1998:227-253.
[23] CLARK J H,SCHRADER W T,O’MALLEY B W.Mechanisms of action of steroid hormones[M]//WILSON J D,FOSTER D W.Williams Textbook of Endocrinology.8th ed.Philadelphia:Saunders,1992:35-90.
[24] DEES C,FOSTER JS,AHAMED S,et al.Dietary estrogens stimulate human breast cells to enter the cell cycle[J].Environmental Health Perspectives,1997,105(Suppl.3):633-636.
[25] ABID-ESSEFI S,BAUDRIMONT I,HASSEN W,et al.DNA fragmentation,apoptosis and cell cycle arrest induced by zearalenone in cultured DOK,Vero and Caco-2 cells:prevention by vitamin E[J].Toxicology,2003,192(2/3):237-248.
[26] OUANES Z,AYED-BOUSSEMA I,BAATI T,et al.Zearalenone induces chromosome aberrations in mouse bone marrow:preventive effect of 17β-estradiol,progesterone and vitamin E[J].Mutation Research/Genetic Toxicology and Environmental Mutagenesis,2005,565(2):139-149.
[27] AVANTAGGIATO G,HAVENAAR R,VISCONTI A.Evaluation of the intestinal absorption of deoxynivalenol and nivalenol by an in vitro gastrointestinal model,and the binding efficacy of activated carbon and other adsorbent materials[J].Food and Chemical Toxicology,2004,42(5):817-824.
[28] HUWIG A,FREIMUND S,KÄPPELI O,et al.Mycotoxin detoxication of animal feed by different adsorbents[J].Toxicology Letters,2001,122(2):179-188.
[29] BATA Á,LÁSZTITY R.Detoxification of mycotoxincontaminated food and feed by microorganisms[J].Trends in Food Science & Technology,1999,10(6/7):223-228.
[30] EL-SHARKAWY S,ABUL-HAJJY J.Microbial cleavage of zearalenone[J].Xenobiotica,1988,18(4):365-371.
[31] KAKEYA H,TAKAHASHI-ANDO N,KIMURA M,et al.Biotransformation of the mycotoxin,zearalenone,to a non-estrogenic compound by a fungal strain of Clonostachys sp.[J].Bioscience,Biotechnology,and Biochemistry,2002,66(12):2723-2726.
[32] UTERMARK J,KARLOVSKY P.Role of zearalenone lactonase in protection of Gliocladium roseum from fungitoxic effects of the mycotoxin zearalenone[J].Applied and Environmental Microbiology,2007,73(2):637-642.
[33] VARGA J,PÉTERI Z,TÁBORI K,et al.Degradation of ochratoxin A and other mycotoxins by Rhizopus isolates[J].International Journal of Food Microbiology,2005,99(3):321-328.
[34] EL-SHARKAWY S,ABUL-HAJJ Y.Microbial transformation of zearalenone,Ⅰ.Formation of zearalenone-4-O-β-glucoside[J].Journal of Natural Products,1987,50(3):520-521.
[35] EL-SHARKAWY S H,ABUL-HAJJ Y J.Microbial transformation of zearalenone.2.Reduction,hydroxylation,and methylation products[J].The Journal of Organic Chemistry,1988,53(3):515-519.
[36] EL-SHARKAWY SH,SELIM M I,AFIFI M S,et al.Microbial transformation of zearalenone to a zearalenone sulfate[J].Applied and Environmental Microbiology,1991,57(2):549-552.
[37] JARD G,LIBOZ T,MATHIEU F,et al.Transformation of zearalenone to zearalenone-sulfate by Aspergillus spp.[J].World Mycotoxin Journal,2010,3(2):183-191.
[38] MOLNAR O,SCHATZMAYR G,FUCHS E,et al.Trichosporon mycotoxinivorans sp.nov.,a new yeast species useful in biological detoxification of various mycotoxins[J].Systematic and Applied Microbiology,2004,27(6):661-671.
[39] VEKIRU E,HAMETNER C,MITTERBAUER R,et al.Cleavage of zearalenone by Trichosporon mycotoxinivorans to a novel nonestrogenic metabolite[J].Applied and Environmental Microbiology,2010,76(7):2353-2359.
[40] AZIZ N H.The role of chitinese of Serratia marcescens in controlling the production of zearalenone by Fusarium graminearum[J].Acta Microbiologica Polonica,2002,51(2):131-137.
[41] ABDULLAH D A.Plasmid-mediated detoxification of mycotoxin zearalenone in Pseudomonas Sp.ZEA-1[J].American Journal of Biochemistry and Biotechnology,2007,3(3):150-158.
[42] ALTALHI A D,EL-DEEB B.Localization of zearalenone detoxification gene(s)in pZEA-1 plasmid of Pseudomonas putida ZEA-1 and expressed in Escherichia coli[J].Journal of Hazardous Materials,2009,161(2/3):1166-1172.
[43] LU Q J,LIANG X C,CHEN F.Detoxification of Zearalenone by viable and inactivated cells of Plano-coccus sp[J].Food Control,2010,22(2):191-195.
[44] KOLLARCZIK B,GAREIS M,HANELT M.In vitro transformation of the Fusarium mycotoxins deoxynivalenol and zearalenone by the normal gut microflora of pigs[J].Natural Toxins,1994,2(3):105-110.
[45] YI P J,PAI C K,LIU J R.Isolation and characterization of a Bacillus licheniformis strain capable of degrading zearalenone[J].World Journal of Microbiology and Biotechnology,2011,27(5):1035-1043.
[46] TINYIRO S E,YAO W R,SUN X L,et al.Scavenging of zearalenone by Bacillus strains-in vitro[J].Research Journal of Microbiology,2011,6(3):304-309.
[47] TINYIRO S E,WOKADALA C,XU D,et al.Adsorption and degradation of zearalenone by Bacillus strains[J].Folia Microbiologica,2011,56(4):321-327.
[48] CHO K J,KANG JS,CHO W T,et al.In vitro degradation of zearalenone by Bacillus subtilis[J].Biotechnology Letters,2010,32(12):1921-1924.
[49] LEI Y P,ZHAO L H,MA Q G,et al.Degradation of zearalenone in swine feed and feed ingredients by Bacillus subtilis ANSB01G[J].World Mycotoxin Journal,2014,7(2):143-151.
[50] 雷元培.ANSB01G菌对玉米赤霉烯酮的降解机制及其动物试验效果研究[D].博士学位论文.北京:中国农业大学,2014.
[51] TAKAHASHI-ANDO N,KIMURA M,KAKEYA H,et al.A novel lactonohydrolase responsible for the detoxification of zearalenone:enzyme purification and gene cloning[J].The Biochemical Journal,2002,365(Pt.1):1-6.
[52] TAKAHASHI-ANDO N,OHSATO S,SHIBATA T,et al.Metabolism of zearalenone by genetically modified organisms expressing the detoxification gene from Clonostachys rosea[J].Applied and Environmental Microbiology,2004,70(6):3239-3245.
[53] IGAWA T,TAKAHASHI-ANDO N,OCHIAI N,et al.Reduced contamination by the Fusarium mycotoxin zearalenone in maize kernels through genetic modification with a detoxification gene[J].Applied and Environmental Microbiology,2007,73(5):1622-1629.
猜你喜欢
当代水产(2022年1期)2022-04-26
少儿科学周刊·少年版(2015年3期)2015-07-07
少儿科学周刊·少年版(2015年3期)2015-07-07
食品工业科技(2014年15期)2014-03-11
养殖与饲料(2014年10期)2014-02-28
鸭绿江(2013年12期)2013-03-11
养禽与禽病防治(2012年3期)2012-08-01
养禽与禽病防治(2012年4期)2012-02-03
中国火炬(2011年7期)2011-07-25
养禽与禽病防治(2011年6期)2011-02-05
