
猪抗病营养研究进展
2014-09-20 02:19陈代文毛湘冰德张克英
动物营养学报 2014年10期
陈代文 毛湘冰 余 冰 何 军 吴 德张克英 郑 萍 虞 洁
(四川农业大学动物营养研究所,教育部动物抗病营养重点实验室,成都 611130)
猪的健康水平是影响养猪生产水平和效益的重要因素,在规模化养殖条件下猪健康的重要性尤为突出。近几年来,我国养猪业一直面临疫病的威胁和困扰,不但使其生产潜力不能充分发挥,而且因防病治病大量使用药物导致猪肉安全质量得不到保障,严重影响养猪业的可持续发展。解决猪健康问题必须依靠综合措施,在继续加强和规范猪病防治的疫苗和药物管理的同时,寻求新思路,研究和应用新理论、新技术、新产品十分重要和必要。抗病营养理念和技术则属此范畴。
现代医学和生物学研究表明,营养是决定健康的关键因素。四川农业大学于2005年提出“抗病营养”(disease-resistant nutrition)的概念,这是一个研究动物营养与健康之间关系的新兴交叉领域。通过研究,揭示动物健康的营养调控规律与机制,建立抗病营养原理和技术,进而提高动物对应激和疾病抵抗力,确保动物健康,减少疾病,降低用药,取消药物饲料添加剂,最终实现畜产品的安全高效生产。
1 抗病营养概念与研究内容
营养物质是一切生命活动的物质基础,既影响动物生产潜力和效率,也决定了动物健康状况。
关于营养与健康的关系研究已有很长的历史。早期的研究至少可追溯到19世纪中叶,研究内容集中在营养缺乏和过量中毒的危害方面,逐步认识营养与健康的表观关系。从20世纪中后期开始,随着免疫学的发展,动物营养学开始探索营养与免疫的关系,并逐步拓展到应激—免疫—营养的互作方面。但直至今天,尽管有不少文章和著作发表[尤其是Adams[1]在2001年提出了动物营养与免疫系统、胃肠道、非感染性疾病之间的关系,并认为全方位营养对畜禽养殖的重要性],但该领域的研究很不系统和深入。从广度上看,营养不仅可以通过调节免疫功能而影响健康,而且可以通过调节动物抗病基因的表达、调控肠道微生态环境、影响特异性疾病的发生发展过程、干预霉菌毒素和自由基对健康的危害等多种途径影响动物健康,但其定性定量作用规律尚未建立;从深度上看,营养可调节动物内分泌、生长相关因子、免疫相关因子的表达和肠道微生物菌群结构及代谢产物而影响动物健康,但其分子机制及其信号转导途径尚不清楚。同时,由于营养素在动物体内代谢具有交互作用,这不仅增加了研究的难度,也影响营养素在动物机体中抗病效应的发挥。因此,营养与健康问题仍然是一个科学难题,传统的营养与免疫的研究不能涵盖全部范畴。研究并揭示营养对动物健康的影响规律及其机制是动物科学的重大理论课题。开展这一难题和课题的研究不仅有助于深入认识生命的本质,而且可为动物健康养殖和高效安全生产提供理论基础。
四川农业大学在深入分析和总结相关学科发展并结合自身多年研究与实践的基础上,于2005年提出“抗病营养”的概念,用此术语来统领营养与健康关系研究领域,同时也显示营养对健康的特殊作用和营养学朝着保健和抗病方向分支发展的必然性和必要性。目前,抗病营养的理念逐渐得到广大学者和行业的认可,抗病营养理论和技术体系的雏形已经凸显。
抗病营养学是动物营养学与免疫学、生理学、病理学、分子和细胞生物学的交叉领域,其目的是要回答动物吃什么、吃多少和怎么吃才能确保动物最大抗病力。抗病营养研究内容及目标见图1,具体内容包括以下几个方面:1)营养与免疫。本方向重点研究营养对免疫系统发育和免疫功能发挥的影响,建立提高动物一般免疫力的营养原理和技术。2)营养与肠道健康。本方向重点研究营养对肠道发育、功能和微生态环境的影响,建立确保肠道健康的营养原理和技术。3)营养与抗病基因。本方向重点研究营养与抗病基因表达的互作规律,探索抗病的表观遗传规律,建立抗病遗传的营养调控原理和技术。4)营养与应激。本方向重点研究各类应激对营养代谢和需要的影响,建立抗应激营养原理和技术。5)营养与疾病。本方向重点研究营养对疾病发生发展过程的影响及其机制,建立提高特异疾病抵抗力的营养原理和技术。6)营养与抗营养因子。本方向重点研究抗营养因子(重点是霉菌毒素)对免疫应答和营养代谢的影响,建立缓解其危害的营养原理和技术。
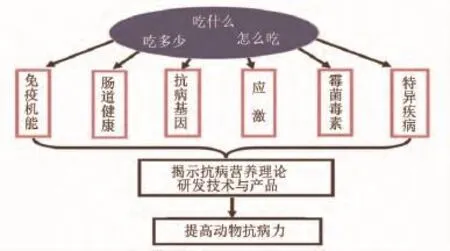
图1 抗病营养研究内容及目标Fig.1 The content and aim of disease-resistant nutrition
开展并完成上述研究,需要营养学家、生化学家、生理学家、免疫学家、病理学家等多学科专家的共同参与,需要将传统营养学和医学手段与现代分子生物学技术、基因工程技术以及组学技术等先进技术与方法相结合,需要长期坚持和稳定投入,方能取得进展和突破。其中,最难的难点当属科学合理的研究模型的建立,包括建模方法、评价标识、模型可靠性、稳定性和适用性等。
2 猪抗病营养研究进展
四川农业大学近几年在猪抗病营养方面开展了系统研究,涉及内容包括:营养与猪的免疫调控;营养与肠道微生态环境及健康的关系;猪抗病力差异的分子机制及抗病基因表达的营养调控;氧化应激及抗氧化营养;营养与病原微生物关系;霉菌毒素及其防霉抗霉营养措施。
2.1 抗病营养研究模型
成功构建抗病营养研究模型是开展深入研究的基础。本团队探索了猪活体和细胞层面的免疫应激模型、氧化应激模型、病原微生物诱导的疾病模型、霉菌毒素攻击模型,构建了适合一定条件的研究模型[2-11]。但由于每种模型均涉及由神经和内分泌系统参与的复杂的生理病理反应,要选择最佳应激源、确定适宜引入方式和剂量、筛选敏感监测标识、明确应激持续时间都十分困难,有待深入研究。
2.2 营养与非特异性免疫
本团队就蛋白质、氨基酸、维生素、矿物元素等养分和植物提取物、微生态制剂、益生素等非营养性物质与仔猪和母猪一般免疫力(非特异性免疫力)的关系进行了较系统的研究,以淋巴细胞数量、T细胞亚群组成、免疫球蛋白浓度、细胞因子浓度和基因表达量等为评价标识,提出了正常状况下保证猪最佳免疫力的养分需要量。
以仔猪为对象,研究了饲粮蛋白质水平和氧化锌添加量对体液免疫、细胞免疫及肠道黏膜免疫的影响,弄清了锌与蛋白质水平互作影响免疫功能的作用机制,确定了仔猪饲粮适宜的氧化锌与蛋白质水平[12-13]。
以生长猪为对象,探讨了部分氨基酸和维生素对机体免疫力的影响及机制。研究发现,饲粮中添加色氨酸和苏氨酸促进免疫球蛋白的合成与分泌,提高猪的生长性能[9]。
以妊娠母猪、哺乳母猪和断奶仔猪为对象,研究了营养水平、硒源和硒水平对母猪繁殖性能和仔猪生长性能及免疫功能的调节效应,确定了母猪和仔猪获得最佳免疫保护效应的硒源和硒水平及母猪获得最佳繁殖性能的营养水平[14-16]。
妊娠鼠模型研究发现,适度提高维生素A、维生素E和色氨酸的供给量可改善体液免疫功能,提高Th2型细胞因子的表达,改善Th1/Th2免疫平衡,从而给胚胎发育提供免疫保护,提高胚胎存活率[17-19]。
此外,本团队的研究还发现,在母猪和仔猪饲粮中添加寡糖、植物提取物和微生态制剂等也可提高母猪繁殖性能和仔猪免疫力[14,20-21]。
通过研究,初步构建了营养调控非特异性免疫力的途径与机制,见图2。
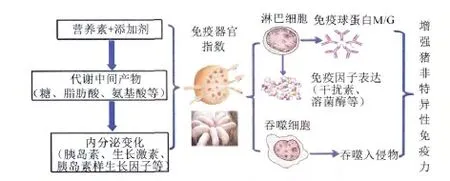
图2 营养调控猪非特异性免疫力的途径与机制Fig.2 The way and mechanism of nutrients regulating non-specific immunity in swine
2.3 营养与肠道健康
有学者应用猪全基因组表达谱芯片技术研究了饲粮及断奶应激对仔猪小肠发育的分子机制,从一定程度上揭示了哺乳仔猪肠道发育过程的分子基础,以及不同蛋白质源饲粮对断奶仔猪肠道发育影响的分子差异[22]。同时发现,出生后3日龄和断奶(21日龄)后3 d是仔猪肠道发育的关键转折点,蛋白质源不同影响肠道发育[22-23](图3)。
宫内发育迟缓(IUGR)显著影响仔猪肠道发育和健康,肠道形态结构、肠细胞代谢能力、抗氧化能力、线粒体功能等均存在明显缺陷,改变营养模式可能缓解IUGR的缺陷;与肠外营养比,采用全肠营养更有利于肠道健康,降低坏死性肠炎(NEC)发生率;初生仔猪饲喂初乳较配方乳更能促进仔猪消化道发育,增强消化酶的活性,阻止微生物黏附,降低NEC发生率[24]。
营养源不同对仔猪肠道健康的影响也不同[25-28]。用21日龄断奶仔猪进行的系列研究发现,蛋白质、淀粉和脂肪来源不同,仔猪生产性能、肠道形态结构、消化酶活性、大肠菌群结构、微生物代谢产物均存在显著差异。酪蛋白优于大豆蛋白,玉米蛋白最差,添加氨基酸平衡氨基酸模式后不能完全消除这种差异;豌豆淀粉优于玉米或小麦淀粉,木薯淀粉最差;椰子油优于玉米油或鱼油,棕榈油最差。同时发现,就肠道健康而言,纯合饲粮不如自然配合饲粮。营养源差异的机制与宿主内分泌、微生物结构和代谢产物有关,营养源、宿主、微生物相互作用决定了肠道健康(图4)。

图3 仔猪肠道发育全基因表达谱及营养调控关键时间节点与营养要点Fig.3 The whole-profile gene expression in the gut of piglets,and the key timepoint of nutrients that were regulated by nutrition

图4 营养源对猪肠道健康的影响及其机制Fig.4 The effect and mechanism of nutrient sources on gut health of swine
合理使用低聚糖、活菌制剂、酸化剂、酶制剂、丁酸钠、高剂量铜均可改善肠道微生态平衡,保障肠道健康,而长期或大量使用抗生素可破坏肠道微生态平衡[20-21,29-32]。
饲料形态(颗粒或粉料)或状态(固体或液体)影响仔猪肠道健康[33]。
在机制方面,初步探索了营养因素对肠道形态结构、分泌功能、免疫功能和微生物菌群的影响,构 建 了 肠 道 保 健 营 养 原 理[13,20-22,29-31,34-35](图5)。
2.4 营养与抗病基因
对藏猪、梅山猪和杜长大(DLY)猪的比较研究发现,Toll样受体(TLR)、β -防御素(pBD)、维甲酸诱导基因(RIG-1)、线粒体抗病毒信号蛋白(MAVS)等抗病基因的表达量具有显著的品种差异和组织特异性[36-39]。藏猪的表达量显著高于梅山猪,后者又高于DLY猪(图6)。
以猪小肠细胞系IPEC-J2为研究对象,研究了维生素A、维生素D、维生素E、微量元素锌、精氨酸和异亮氨酸对pBD表达的调控效应[36-37]。结果表明,所研究的营养物质都能够不同程度地诱导猪小肠上皮细胞pBD基因的表达和蛋白分泌。核转录因子-κB(NF-κB)通路和丝裂原激活/细胞外信号调节激酶-细胞外信号调节蛋白激酶(MEK-ERK)通路可能分别是维生素A影响pBD-2基因表达和影响pBD-1、pBD-3基因表达的信号通路之一。
用DLY仔猪或藏猪进行的活体试验发现,在正常和病原微生物攻毒2种状态下,饲粮添加维生素 A、维生素 D可提高组织中 pBD-1、pBD-3、TLR、RIG-1 和 MAVS 基因的表达量[36,39];添加精氨酸有缓解应激导致的采食量下降的效应,增加各组织的pBD-1、pBD-2、pBD-3基因表达量,显著下调因免疫应激导致的仔猪Toll样受体4(TLR4)和Toll样受体5(TLR5)及其下游信号分子髓样分化因子88(MyD88)、NF-κB/p65 mRNA 等的过度表达,降低炎症因子白细胞介素-6(IL-6)、干扰素 -γ(IFN-γ)和肿瘤坏死因子 - α(TNF-α)等的产生,进而缓解免疫应激[5,37];一氧化氮(NO)是介导精氨酸(Arg)调节免疫应激状态下TLR4信号途径的关键信号分子[37](图6)。

图5 动物肠道保健营养原理Fig.5 The nutritional mechanism of health protection in animal gut

图6 不同品种猪抗病力差异分子机制及抗病基因表达的营养调控Fig.6 The molecular mechanism of disease-resistant difference among the different breeds of swine,and the nutritional regulation of the expression of disease-resistant genes
2.5 营养与应激
2.5.1 免疫应激
以仔猪为对象,以注射脂多糖(LPS)为手段,建立了仔猪免疫应激模型。结果表明,免疫应激后短期内可导致仔猪处于高度免疫活化状态,采食量急剧下降,通过抑制胰高血糖素样肽-2(GLP-2)分泌而影响仔猪肠道形态学(绒毛高度/隐窝深度)和肠黏膜乳糖酶、蔗糖酶及碱性磷酸酶的活性;通过神经内分泌途径改变养分代谢方向,仔猪整体蛋白质代谢朝着分解代谢加强、合成代谢降低的方向进行;营养需要模型发生改变,应激初期饲粮整体蛋白质需要量降低,理想氨基酸模式发生变化,苏氨酸和色氨酸与赖氨酸之比大大提高,应激后期的恢复期,蛋白质和赖氨酸需要量增加[7,10,40-41]。对锌的研究发现,NRC(1998)推荐的仔猪锌需要量足以满足正常条件下仔猪生长和适宜免疫力的需要,但在免疫应激条件下需要大大提高锌的添加量[35]。营养缓解免疫应激的途径与机制如图7。
2.5.2 氧化应激
饲喂氧化鱼油和腹腔注射diquat都可诱导仔猪产生氧化应激[3,15,42]。表现为生产性能、养分利用率及组织抗氧化能力下降,血相改变,组织细胞过氧化损伤加剧,肠黏膜损伤,养分转运载体的表达量和养分的转运能力显著降低,机体免疫应答能力下降。

图7 营养缓解免疫应激的途径与机制Fig.7 The way and mechanism of nutrients alleviating immune stress
在氧化应激状况下,仔猪硒的代谢和需要量发生变化,0.2 mg/kg的硒添加量不能满足机体抗氧化的需要;硒抗氧化应激的机制可能与影响三碘甲状腺原氨酸(T3)、甲状腺素(T4)的分泌、调节细胞因子白细胞介素-1β(IL-1β)和白细胞介素-6(IL-6)的生成以及NF-κB活性有关;同时,氧化应激导致仔猪体内色氨酸和精氨酸的代谢改变[15]。氧化应激通过增强肝脏色氨酸2,3-加双氧酶活性来提高断奶仔猪色氨酸分解代谢,通过提高肠道氨基酸转运载体(CAT-1)mRNA表达、增加精氨酸内源合成关键酶鸟氨酸转氨酶(OAT)的活性、降低肝脏和肺的诱导型一氧化氮合酶(iNOS)和内皮型一氧化氮合酶(eNOS)的mRNA的表达来改变精氨酸的吸收和代谢[43]。在应激条件下,提高精氨酸添加量可以通过调控精氨酸的代谢、维持精氨酸内源稳定性、增加精氨酸的有效性、调控内分泌、降低炎症因子的表达等综合途径缓解氧化应激;增加色氨酸添加量则通过影响胰岛素样生长因子-1(IGF-1)系统基因、促食欲相关基因及炎症相关基因表达,调控仔猪生长、食欲及炎症过程,发挥其抗应激作用[44](图8)。
2.6 营养与疾病
在构建模型基础上,系统研究了营养对病毒[猪圆环病毒(PCV2)、轮状病毒、伪狂犬病毒(PRV)]、弱毒苗(猪瘟、伪狂犬病弱毒、猪繁殖和呼吸综合征)和细菌(大肠杆菌、沙门氏菌杆菌)攻击下仔猪的保护效应,揭示了营养调节猪特异性免疫力的机理和可能途径[4-6,9,11,19,25,39,45-46](图 9)。
生物素和叶酸促进IFN-γ合成,缓解PCV2对猪淋巴组织的破坏,促进免疫应答反应和PCV2抗体生成,发病率降低 10%[4,6]。维生素 D 通过促进仔猪轮状病毒(RV)抗体和肠道黏膜分泌型免疫球蛋白A(SIgA)生成,调节RIG-1信号通路关键分子表达,降低RV攻击下仔猪肠道黏膜炎症反应,显著降低腹泻率[39](图 10)。
精氨酸通过提高母猪高致病性猪蓝耳病病毒(PRRSV)抗体水平,降低死胎率和弱仔率;血浆蛋白粉作为蛋白质来源能显著提高接种猪瘟弱毒苗仔猪抗体效价,且显著提高其血清IFN-γ及免疫球蛋白水平[11,45]。色氨酸可促进生长猪IGF-1分泌及特异性抗体合成,缓解因接种PRRSV和PRV弱毒苗引起的生长抑制[9,19]。苏氨酸通过增强淋巴细胞功能、促进特异和非特异性抗体产生、下调TLR表达、减少炎性因子产生以及促进蛋白质合成等途径缓解PRV对仔猪的损伤[9,46](图 11)。
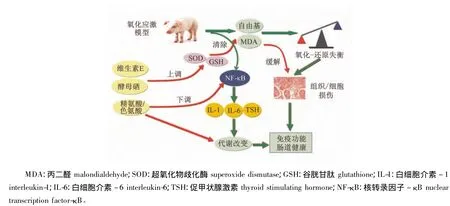
图8 营养缓解氧化应激的途径与机制Fig.8 The way and mechanism of nutrients alleviating oxidative stress
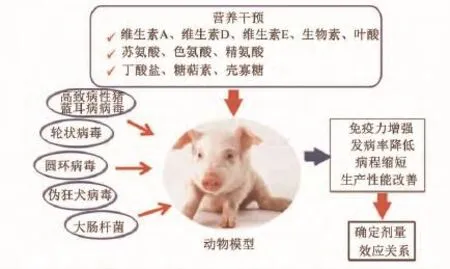
图9 营养调控特异性疾病的可能途径和机制Fig.9 The possible way and mechanism of nutrients regulating specific diseases
椰子油和鱼油可通过改善仔猪糖脂代谢、肠道发育及免疫相关分子的表达,缓解大肠杆菌(O8∶K87/K88ab)攻击对仔猪生产性能和健康的负面影响[25]。精氨酸可下调仔猪TLR4和TLR5及其信号途径关键分子表达,缓解沙门氏菌杆菌(S.C500)对免疫系统的激活[5]。
2.7 营养与霉菌毒素
本团队重点研究了霉菌毒素对健康的危害及防霉抗霉营养技术。在调查分析饲料霉菌毒素污染现状[47]的基础上,以仔猪和生长育肥猪为对象,研究揭示了自然霉变饲粮(以含镰刀菌毒素或黄曲霉毒素为主)在导致采食量下降、小肠黏膜结构损伤方面的毒性效应,发现自然霉变玉米导致仔猪肝脏和空肠黏膜氧化损伤,改变空肠形态结构和养分转运载体基因表达量,降低养分消化率和转运能力,改变肠道微生物平衡,进而抑制仔猪生长[2,48]。仔猪采食量下降的原因是血清中与采食调节相关的激素水平发生了明显变化[48](图12)。
通过检测机体氧化还原状态发现,霉菌毒素的危害可能与毒素导致的机体氧化与抗氧化系统失衡有关[49]。进一步的系列研究证实了这一推论。体内外试验揭示,硒和维生素E能够缓解霉菌毒素对大鼠和仔猪肠细胞的氧化损伤,降低霉菌毒素的危害;且硒是通过增强核因子E2相关因子2/抗氧化反应元件(Nrf2/ARE)途径的防御作用而发挥其降低毒素危害效应[49-50](图12)。
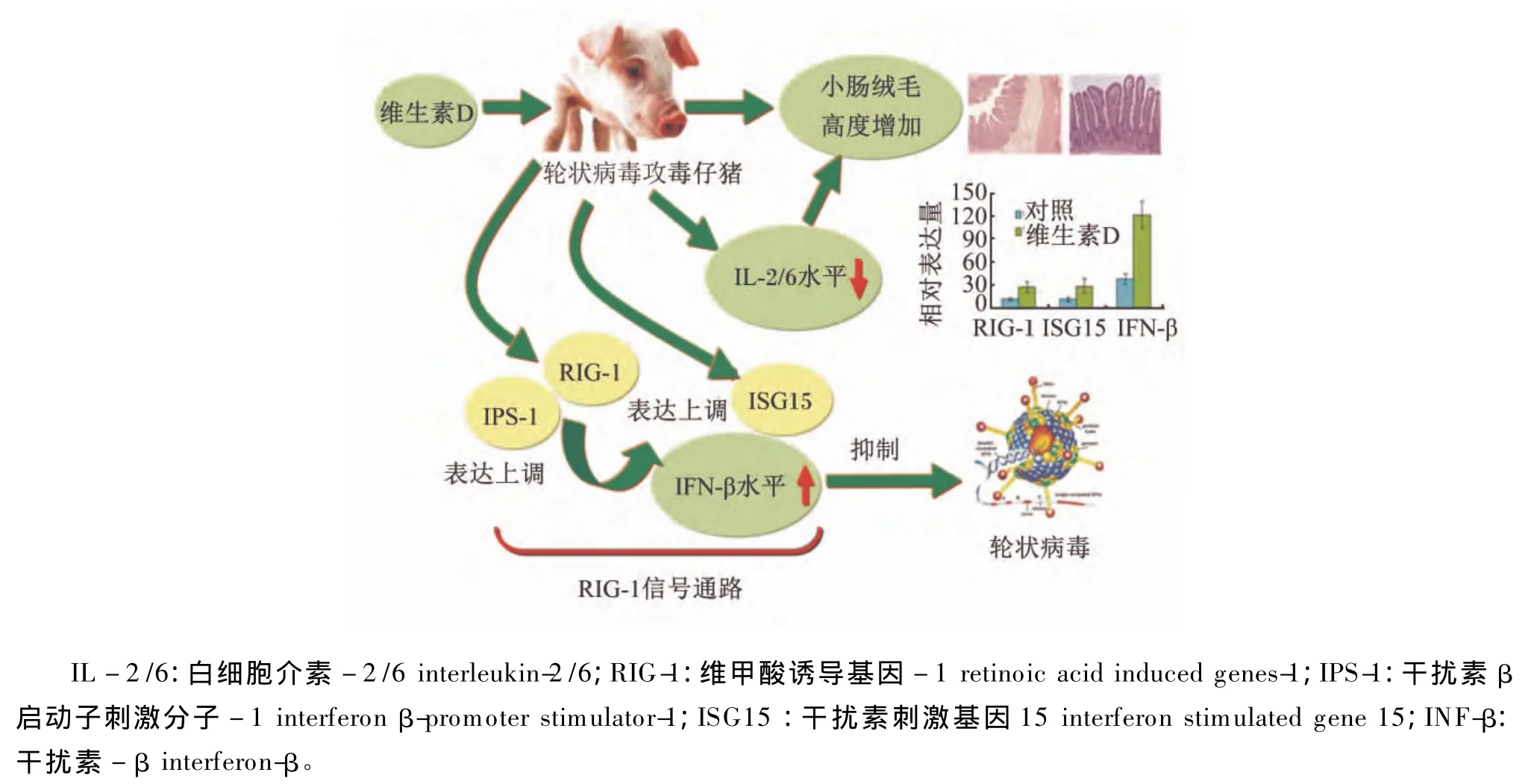
图10 维生素D对轮状病毒攻毒的保护机制Fig.10 The protective mechanism of vitamin D on rotavirus challenge in pigs
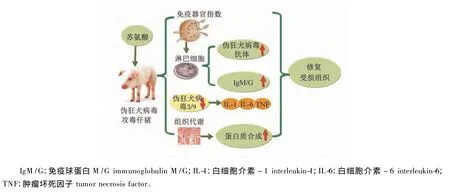
图11 苏氨酸伪狂犬攻毒的保护机制Fig.11 The protective mechanism of threonine on pseudorabies virus challenge in pigs
3 抗病营养的应用
抗病营养的应用技术包括:第一,应用抗病营养需要参数配制抗病饲料;第二,开发应用抗病饲料新产品,包括生物蛋白质饲料、生物能量饲料、生物饲料添加剂、功能性配合饲料等。
通过近几年的研究,初步探明了主要养分的抗病营养原理和功效,建立了10多种养分的抗病营养需要量,研制了10多种具有抗病功效的生物饲料及饲料添加剂,一些产品已在企业生产使用。应用这些成果,规模猪场的发病率可以降低20%~30%,生产性能和效益显著改善,药物使用量显著下降,显示抗病营养具有明显的功效和潜力。

图12 霉菌毒素的危害及营养防霉抗霉的机制Fig.12 The harm of mycotoxins and the mechanism of nutrients resisting mycotoxin
4 小结
抗病营养的研究与应用符合养猪业可持续发展的高效、安全、优质和环保要求,具有广阔的应用前景。由于此领域的研究时间短,很多问题还有待深入研究,理论与技术体系尚需进一步完善。继续开展抗病营养理论与技术体系及其机制研究对于深入认识健康的本质、确保猪的健康和猪肉安全具有重大的理论意义和实践价值。
[1]ADAMS C A.Total nutrition:feeding animals for health and growth[M].England:Nottingham University Press,2001.
[2]雷晓娅.自然霉变玉米及甘露寡糖对仔猪生长性能和肠道健康的影响[D].硕士学位论文.雅安:四川农业大学,2011.
[3]徐静,余冰,陈代文.Diquat诱导的生长猪氧化应激持续时间及适宜的应激标识[J].中国农业科学,2008,41(12):4359-4364.
[4]陈宏.生物素对断奶仔猪生产性能及免疫功能影响的研究[D].博士学位论文.雅安:四川农业大学,2008.
[5]陈渝,陈代文,毛湘冰,等.精氨酸对免疫应激仔猪肠道组织Toll样受体基因表达的影响[J].动物营养学报,2011,23(9):1527-1535.
[6]高庆.饲粮添加叶酸对断奶仔猪生产性能和免疫功能的影响研究[D].博士学位论文.雅安:四川农业大学,2011.
[7]李建文,陈代文,张克英,等.免疫应激对仔猪理想氨基酸平衡模式影响的研究[J].畜牧兽医学报,2006,37(1):34-37.
[8]廖波.25-OH-D3对免疫应激断奶仔猪的生产性能、肠道免疫功能和机体免疫应答的影响[D].博士学位论文.雅安:四川农业大学,2011.
[9]沈杰.添加苏氨酸和色氨酸对接种猪繁殖与呼吸综合症(PRRS)弱毒苗生长猪生产性能和血液参数的影响[D].硕士学位论文.雅安:四川农业大学,2009.
[10]吴春燕.免疫应激对断奶仔猪整体蛋白质周转代谢的影响[D].硕士学位论文.雅安:四川农业大学,2002.
[11]詹黎明.不同蛋白来源对接种猪瘟弱毒苗仔猪生性能、肠道形态和免疫功能影响的研究[D].硕士学位论文.雅安:四川农业大学,2010.
[12]刘军.饲粮不同蛋白质和锌水平对早期断奶仔猪血液和淋巴器官中细胞因子的影响[D].硕士学位论文.雅安:四川农业大学,2011.
[13]岳双明.不同蛋白水平日粮添加高锌对早期断奶仔猪生产性能、抗氧化作用和肠道粘膜免疫的影响[D].硕士学位论文.雅安:四川农业大学,2008.
[14]黎文彬.酵母硒对脂多糖(LPS)诱导的免疫应激早期断奶仔猪的影响[D].硕士学位论文.雅安:四川农业大学,2009.
[15]袁施彬.仔猪氧化应激及硒的抗应激效应和机理的研究[D].博士学位论文.雅安:四川农业大学,2007.
[16]林燕.母体纤维营养对胎儿和生后生长发育及抗氧化能力的影响[D].博士学位论文.雅安:四川农业大学,2011.
[17]罗晓容.维生素A对感染伪狂犬病毒妊娠雌鼠免疫功能及繁殖性能的影响[D].硕士学位论文.雅安:四川农业大学,2009.
[18]罗小林.维生素E对病毒感染妊娠雌鼠繁殖性能及免疫功能的保护效应[D].硕士学位论文.雅安:四川农业大学,2009.
[19]邱时秀.日粮色氨酸水平对感染伪狂犬病毒妊娠雌鼠繁殖性能和免疫功能的影响[D].硕士学位论文.雅安:四川农业大学,2009.
[20]陈代文,张克英,王万祥,等.酸化剂、益生素和寡糖对断奶仔猪粪中微生物茵群和免疫功能的影响及其互作效应研究[J].动物营养学报,2006,18(3):172-178.
[21]邹健.银耳多糖对断奶仔猪生产性能和肠道菌群及免疫功能的影响[D].硕士学位论文.雅安:四川农业大学,2006.
[22]孙云子.哺乳期及断奶后饲喂不同日粮仔猪小肠发育全基因组表达谱研究[D].博士学位论文.雅安:四川农业大学,2009.
[23]孙云子,余冰,陈代文.不同蛋白源日粮对断奶仔猪小肠形态的影响[J].中国饲料,2011(12):34-37.
[24]车炼强.宫内发育迟缓和营养对新生仔猪消化道生长发育及坏死性肠炎发生机理的研究[D].博士学位论文.雅安:四川农业大学,2010.
[25]刘忠臣.不同来源脂肪对仔猪的营养效应及对E.coli攻毒的保护作用研究[D].博士学位论文.雅安:四川农业大学,2011.
[26]亓宏伟.不同来源蛋白对断奶仔猪肠道微生态环境及肠道健康的影响[D].博士学位论文.雅安:四川农业大学,2011.
[27]相振田.饲粮不同来源淀粉对断奶仔猪肠道功能和健康的影响及机理研究[D].博士学位论文.雅安:四川农业大学,2011.
[28]邹芳.不同种类脂肪对大鼠生长性能及微生态效应的影响研究[D].硕士学位论文,雅安:四川农业大学,2009.
[29]陈代文,张克英,余冰,等.仔猪饲粮添加酸化剂及黄霉素对生产性能、消化道pH和微生物数量的影响[J],中国畜牧杂志,2004,40(4):16-19.
[30]毛倩,陈代文,余冰,等.复合益生素对生长育肥猪生产性能、盲肠菌群及代谢产物的影响[J].中国畜牧杂志,2010(17):34-39.
[31]梅绍锋,余冰,鞠翠芳,等.高锌和高铜对断奶仔猪生产性能、消化生理和盲肠微生物数量的影响[J].动物营养学报,2009,21(6):903-909.
[32]王纯刚,张克英,丁雪梅.丁酸钠对轮状病毒攻毒和未攻毒断奶仔猪生长性能和肠道发育的影响[J].动物营养学报,2009,21(5):711-718.
[33]唐仁勇.饲料不同形态、状态对早期断奶仔猪生产性能及消化生理的影响[D].硕士学位论文.雅安:四川农业大学,2004.
[34]贾刚,蒋荣川,晏家友,等.胰高血糖素样肽 -2对28日龄断奶仔猪肠上皮细胞的保护及修复效应研究[J].畜牧兽医学报,2009,40(10):1478-1486.
[35]孙国君.锌对仔猪及小肠上皮细胞免疫功能的调节作用[D].博士学位论文.雅安:四川农业大学,2009.
[36]陈金永.猪β-防御素基因表达特点及维生素A的调节作用[D].博士学位论文.雅安:四川农业大学,2010.
[37]齐莎日娜.猪β-防御素基因表达特点及精氨酸的调节作用[D].博士学位论文.雅安:四川农业大学,2010.
[38]朱浩妮.不同品种仔猪TLRs表达规律及VA对仔猪TLRs表达量影响的研究[D].硕士学位论文.雅安:四川农业大学,2010.
[39]赵叶.不同猪种抗病毒相关模式识别受体基因表达差异及维生素D的抗病毒作用与机制[D].博士学位论文.雅安:四川农业大学,2013.
[40]车炼强,张克英,丁雪梅,等.免疫应激对仔猪肠道发育及胰高血糖素样肽-2分泌的影响[J].畜牧兽医学报,2009,40(5):676-682.
[41]吕继蓉.免疫应激对仔猪生产性能和蛋白质需求规律的影响[D.]硕士学位论文.雅安:四川农业大学,2002.
[42]李丽娟,陈代文,余冰,等.氧化应激对断奶仔猪肌体氧化还原状态的影响[J].动物营养学报,2007,19(3):199-203.
[43]吕美.氧化应激对仔猪色氨酸代谢和需求特点的影响及机制研究[D].博士学位论文.雅安:四川农业大学,2009.
[44]郑萍.氧化应激对仔猪精氨酸代谢和需求特点的影响及机制研究[D].博士学位论文.雅安:四川农业大学,2010.
[45]杨平.饲粮添加L-精氨酸或N-氨甲酰谷氨酸对感染PRRSV妊娠母猪繁殖性能及免疫功能的影响[D].硕士学位论文.雅安:四川农业大学,2011.
[46]赖翔.饲粮添加苏氨酸对PRV诱导的免疫应激仔猪生长性能和免疫功能的影响[D].硕士学位论文.雅安:四川农业大学,2012.
[47]杨晓飞.四川地区主要饲料霉菌毒素分布规律的研究[D].硕士学位论文.雅安:四川农业大学,2007.
[48]苏军.镰刀菌毒素对猪的抗营养效应及其机制研究[D].博士论学位文.雅安:四川农业大学,2008.
[49]虞洁.自然霉变玉米对大鼠抗氧化能力的影响及抗氧化剂的保护作用研究[D].硕士学位论文.雅安:四川农业大学,2008.
[50]陈平.赭曲霉毒素A对IPEC-J2细胞Nrf2抗氧化系统的影响及硒的保护效应研究[D].硕士学位论文.雅安:四川农业大学,2010.
猜你喜欢
云南农业大学学报(自然科学)(2022年5期)2022-09-22
中国地市报人(2022年6期)2022-07-01
今日农业(2022年4期)2022-06-01
湖南农业大学学报(社会科学版)(2022年1期)2022-02-26
韩国语教学与研究(2017年2期)2017-10-20
浙江农业学报(2017年1期)2017-05-17
上海农业学报(2017年3期)2017-04-10
知识经济·中国直销(2016年5期)2016-11-07
航天器工程(2014年5期)2014-03-11
延河(2013年5期)2013-11-07
