
豇豆叶绿体微卫星标记的开发及其在近缘种的通用性研究
2014-12-07 05:14潘磊李依郭瑞张凤银曾长立胡志辉吴华陈禅友
长江蔬菜 2014年6期
潘磊 ,李依 ,郭瑞 ,张凤银 ,曾长立 ,胡志辉 ,吴华 ,陈禅友
(1.湖北省豆类(蔬菜)植物工程技术研究中心,武汉,430056;2.江汉大学生命科学学院)
豇豆(Vigna unguiculata)(2n=2x=22),又名豆角、长豆角、带豆等,属豆科豇豆属一年生草本植物。豇豆起源于非洲,中国是其次生起源中心,它在我国栽培历史悠久,栽培面积大,南北各地均有分布。当前,豇豆的DNA分子标记研究在国内外越来越受到关注[1]。特别是在豇豆的遗传多样性及品种(系)间的遗传关系等研究方面,开展了多种分子标记技术的研究,如RFLP[2],AFLP[3],RAPD(Random amplified polymorphic DNA,随机扩增多态性 DNA)[4,5]、ISSR(Inter-simple sequence repeat,简单序列重复区间)[6]。 近年来,借助比较基因组学的技术手段,研究者们将豇豆近缘种——普通豇豆SNP(Single nucleotide polymorphism,单核苷酸多态性)[7]和普通豇豆SSR(Simple sequence repeat,简单序列重复)[8,9]等分子标记,成功用于长豇豆基因组的研究。
然而,豇豆核外基因组微卫星标记的研究,特别是叶绿体微卫星(Chloroplast simple sequence repeat,cpSSR)标记的发掘及其应用研究尚未见报道。Powell等[10]首次提出叶绿体cpSSR标记技术。与核基因组SSR标记相比,叶绿体基因组DNA(Chloroplast genome DNA,cpDNA) 呈单亲遗传模式、结构简单、多拷贝、分子量小等特点,此外,cpSSR标记还兼有核基因组SSR标记的共显性、高度变异和多态性等特点,可以用于种质资源分类[11]、亲缘关系鉴定[12]、遗传多样性[13,14]、遗传结构[15]、系统发生学等研究[16,17]。
开发cpSSR标记的传统策略是构建基因组文库和测序,然后设计引物进行筛选,以获得cpSSR引物,但是从植物组织中提取高纯度cpDNA十分困难,而且费时费力。当前,采用生物信息学技术方法,充分发掘公共数据库资源中的叶绿体基因组序列,根据cpSSR侧翼序列的保守性,设计筛选cpSSR引物是一条经济且便捷的途径。
虽然豇豆全基因组测序尚未完成,但是豇豆叶绿体全基因组已经测序完成并公布。本研究通过下载公共数据库中的豇豆叶绿体基因组全序列,分析豇豆叶绿体基因组的结构和组分特征,揭示豇豆cpSSR组成与分布特点,并检验豇豆cpSSR引物在豆科物种间的通用性,以便为豇豆cpSSR标记在其近缘种的群体分化、遗传多样性和系统发育等研究奠定基础。
1 材料与方法
1.1 叶绿体微卫星筛选
从 GenBank公共数据库中(http://www.ncbi.nlm.nih.gov/)下载豇豆(V.unguiculata)叶绿体基因组全序列(NC_018051),对豇豆叶绿体基因组全序列进行分析,筛选微卫星序列。单碱基微卫星采用软件Tandem Repeats Finder 4.04查找[18];多碱基微卫星采用SSRhunter 1.3软件进行分析[19],搜索条件按设置为单碱基微卫星最小重复次数为10、二和三碱基的完全重复微卫星最小重复次数分别为5和4、四碱基及四碱基以上的微卫星最小重复次数均为3。在每个SSR位点的上下游侧翼序列各为150 bp。
此外,采用DOGMA软件 (Dual Organellar Genome Annotator)(http://dogma.ccbb.utexas.edu/)对叶绿体基因组结构进行分析。
1.2 cpSSR引物设计
基于所获得的叶绿体微卫星序列,利用在线引物设计软件 Primer 3.0(http://frodo.wi.mit.edu/)分析微卫星的侧翼序列并进行引物设计。主要参数设置为引物长度18~22 bp,以20 bp为最佳;引物退火温度50~60℃,以55℃为最佳,上游和下游的引物退火温度之差在5℃以内;GC含量40%~70%,最适为50%;预期PCR产物长度为100~400 bp。引物由生工生物工程(上海)股份有限公司合成。
1.3 供试的豆类样本材料
本研究在对引物进行扩增检测时,采用了19份豆类种质资源,包括10份豇豆材料,5份菜豆材料,3份豌豆材料和1份利马豆材料(表1)。
1.4 PCR扩增和电泳分析
PCR反应体系总体积为20 μL,含有1×PCR buffer,20 ng DNA 模 板 ,4 nmol dNTP,30 nmol MgCl2,引物 10 pmol,0.5 U Taq DNA 聚合酶[天根生化科技(北京)有限公司]。在Mastercycler gradient型PCR仪(德国,Eppendorf公司)上进行扩增。PCR扩增反应程序为94℃预变性5 min;然后进行35个循环,每个循环包括 94℃变性 30 s,50~60℃退火 30 s,72℃延伸 40 s;最后 72℃延伸 5 min。
采用8%变性聚丙烯酰胺凝胶(Acrylamide∶Bis=19∶1)检测扩增产物。将PCR产物与上样缓冲液(98%甲酰胺,10 mmol/L EDTA,0.025%溴酚蓝和0.025%二甲苯青蓝)等体积混合,在95°C变性5 min后,立即冰浴3 min,每次点样量2.5 μL,恒功率55 W预电泳40 min,然后恒功率50 W电泳1.5 h。电泳完毕,经过固定、银染、显色并用数码相机拍照。

表1 供试的豆类样品信息

表2 52个cpSSR在豇豆叶绿体基因组上的分布
电泳后,统计每一样品扩增的DNA条带数及其多态性,在同一电泳迁移位置上,有DNA扩增条带记为“1”,无条带记为“0”。
2 结果与分析
2.1 豇豆叶绿体基因组cpSSR的分布
豇豆叶绿体基因组全序列分析表明,其基因组大小为152 415 bp,总GC含量为35.2%。豇豆叶绿体基因组的结构包括1个长单拷贝序列(Long single copy sequence,LSC)、1个短单拷贝序列 (Short singlecopysequence,SSC)及 2 个反向重复序列(Inverted repeat,IR)。其中,LSC约81 kb分布区域为1~81 468 bp,IRa约 26 kb分布区域为 81 469~107 351 bp,SSC 约 19 kb分 布 区 为 107 352~126 250 bp,IRb大约26 kb分布区域为126 251~152 415 bp。
从豇豆叶绿体基因组上共搜索出52个微卫星(表2),分布频率为每2 931 bp有1个 cpSSR;微卫星核心重复区及侧翼序列总长度为16 213 bp,占基因组全长的10.63%。豇豆cpSSR的分布范围较大,从7 245 bp到152 001 bp都有分布,但是其分布并不均匀,主要集中在LSC区,少量的在SSC区 ,而在IR区没有。在LSC区和SSC区分布的cpSSR分别为48个(92.3%)和 4个(7.7%)。

表3 52个豇豆叶绿体cpSSR的重复基序特点
2.2 豇豆叶绿体基因组cpSSR的重复基序特点
根据微卫星核心重复基序的结构特点[20],可将微卫星可分为3种类型,即完美型、非完美型和复合型。完美型指的是微卫星序列没有中断或附近没有其他种类基序的重复序列;非完美型指的是同种重复基序被3个碱基以下的非重复序列所间隔;复合型指一种重复基序与其他种类的重复基序被3个以下的非重复序列所间隔。在所获得的52个豇豆cpSSR中,完美型为48个,非完美型为4个,无复合型(表3)。这表明,在豇豆叶绿体基因组cpSSR中以完美型和非完美型微卫星为主。此外,对核心区的重复次数分析发现,重复次数的变化范围为4~19次,其中核心重复次数为10次的微卫星有14个(26.9%),而核心重复次数为11次的微卫星有11个(21.2%),两者累计所占比例为48.1%,几乎占全部微卫星数量的1/2。
依据重复基序的碱基数目差别,在这52个豇豆cpSSR中,单碱基重复基序微卫星共有33个(完美型29个和非完美型4个),占63.5%,而 A和T是单碱基重复基序的主要类型,未见C或者G的单碱基基序(表3)。二碱基重复基序的微卫星有14个,占26.9%,以AT和TA为主;三碱基重复基序有5个,占9.6%,碱基类型包括AAT (2个)、TTA(1个)、TAA(1 个)、ATA(1 个)。 由此可见,在豇豆叶绿体微卫星中,重复基序的碱基主要以A和T为主。
2.3 cpSSR引物的设计与筛选
利用在线引物设计软件Primer 3.0分析上述52个豇豆叶绿体微卫星序列,从中设计并合成10对cpSSR引物,通过对10份豇豆样品的PCR检测,发现其中8对cpSSR引物具有多态性(图1),且PCR产物片段位于100~400 bp(表4)。
检测上述8对多态性豇豆cpSSR在其近缘种中的可转移性(图2)。选取豇豆的3个近缘物种豌豆、利马豆和菜豆(共9份样品)进行扩增,结果表明,这8对豇豆cpSSR引物均可以在这三3个近缘种中成功扩增(表4)。

图1 引物CPM7在10份豇豆样品中扩增产物的电泳图谱(8%PAGE)
在豌豆中具有多态性的引物有CPM1、CPM3和CPM6,均检测到2个等位基因,其余5对cpSSR引物为单态性;在菜豆中具有多态性的引物有CPM1、CPM2、CPM4、CPM5 和 CPM7,也均检测到 2对等位基因,而其余3对cpSSR引物为单态性。
3 讨论与结论
叶绿体基因组是单倍体,保守性较强,具有细胞质遗传的特点,除少数植物例外,一般为单亲遗传,不发生重组[21]。由于在进化过程中,亲代到子代的遗传物质传递不涉及基因组重组,不会受到选择压力,而且位点发生的突变也容易保留,不会受到重组、缺失以及假基因等的干扰[22]。因此,具有叶绿体基因组特征的cpSSR分子标记技术广泛应用于在植物种质资源鉴定、群体分化、系统分类和物种进化等研究领域[23,24]。
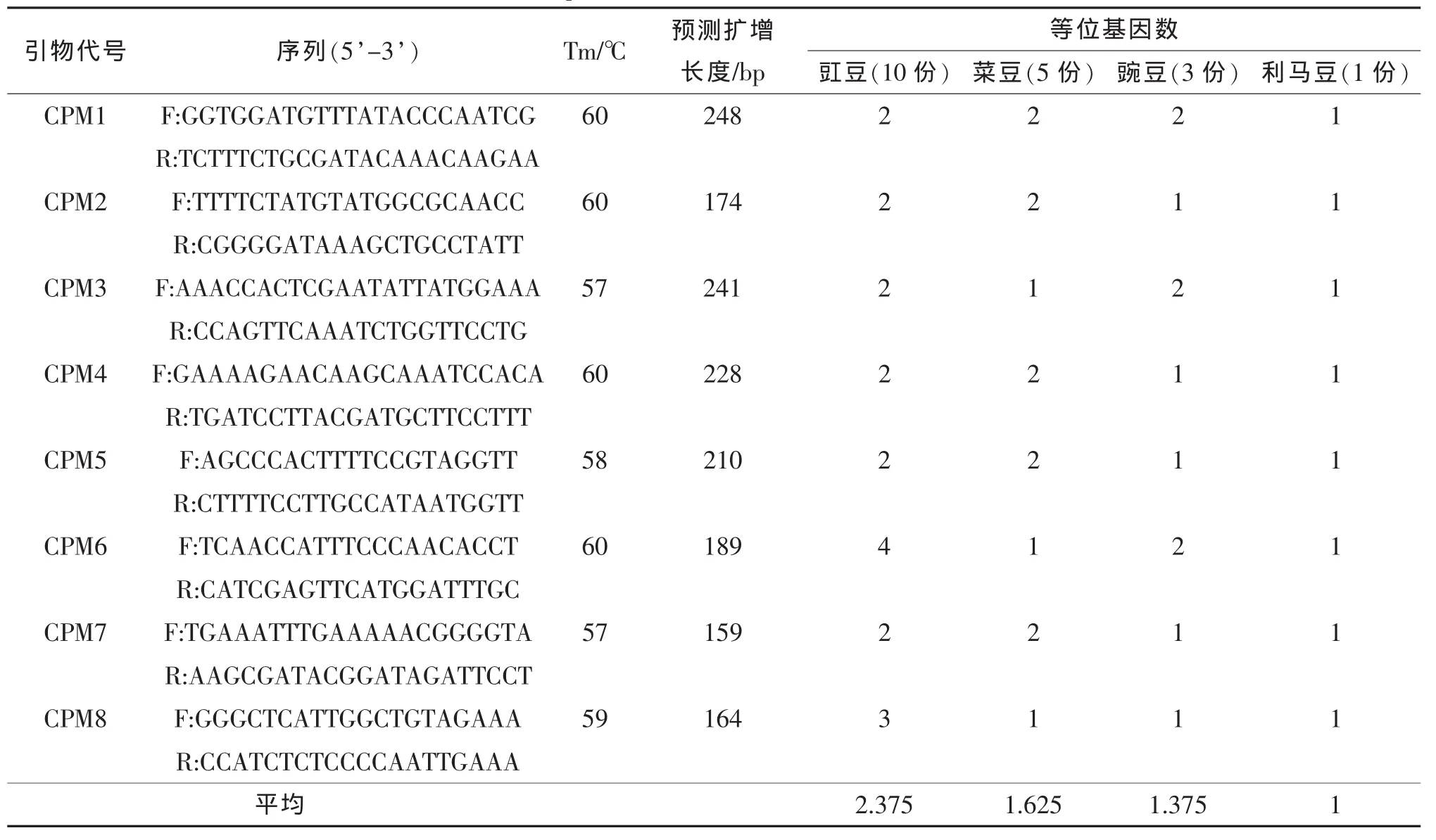
表4 8个豇豆cpSSR引物信息及在其近缘种中的可转移性
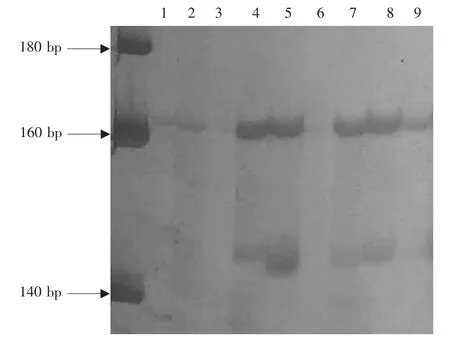
图2 引物CPM7在豌豆、利马豆和菜豆样品中扩增产物的电泳图谱(8%PAGE)
在cpSSR分子标记的应用上,其瓶颈和难点是引物的设计与筛选。与核基因组SSR引物的开发类似,cpSSR引物开发的前提是获取微卫星侧翼的序列信息。随着现代生物技术和基因组学的不断进步,当前cpSSR分子标记的开发主要有两种策略,一是直接进行叶绿体基因组测序,再设计筛选cpSSR引物;二是基于生物信息学手段,或者从公共数据库的叶绿体基因组中发掘cpSSR引物,或者利用近缘物种的cpSSR进行种间的可转移性检测,以获取cpSSR引物。对于前者而言,由于叶绿体基因组DNA的分离和纯化难度较大,增加了测序难度和成本,因此,后者利用公共数据库资源进行叶绿体cpSSR标记的开发,更加简便和经济。特别是随着开展全基因组测序的植物物种越来越多,许多植物的叶绿体基因组如拟南芥[25,26]、黄瓜[22]、蝴蝶兰[27]、柑橘[28,29]、菜豆[30]等的全序列或者部分序列已经释放到公共数据库中,极大促进了cpSSR分子标记的开发和应用。
豇豆的叶绿体基因组全序列刚公布不久,这为揭示豇豆叶绿体基因组特征,并开发豇豆cpSSR分子标记奠定了基础。豇豆叶绿体基因组比较大(约153 kb),而一般植物的叶绿体基因组大小在120~170 kb[7,31]。
本研究通过对豇豆叶绿体全基因进行微卫星分布特征分析,发现在豇豆叶绿体基因组上分布有52个cpSSR,其序列总长约占叶绿体基因组全长的1/10。这些微卫星在豇豆叶绿体基因组上分布不均,分布的密集区域为LSC区,只有少数几个微卫星分布在SSC区,而在IR区无微卫星分布。
这52个豇豆cpSSR在微卫星的序列特征上,以A/T为单碱基重复基序的微卫星为主 (63.5%),也包括少量的二碱基重复基序类型AT/TA和三碱基重复基序类型AAT/TTA/TAA/ATA;重复次数的变化范围为4次到19次不等。因此,在豇豆cpSSR中,无论单碱基、二碱基或者三碱基重复基序,它们的碱基组成均以A/T为主要类型,未见C/G的碱基基序,而且重复次数变异范围有限。cpSSR的这种碱基组成和重复特征可能与叶绿体的结构组成特点及其进化有关联。
由于叶绿体基因组序列在物种间具有较高的保守性,cpSSR引物可以在不同种属间进行跨种扩增,具有通用性。Ishii等[32]研究表明,水稻cpSSR标记在禾本科植物中具有较为广泛的通用性。Diekmann等[33]从多年生黑麦草中发掘出9个cpSSR标记并检验其在黑麦草属物种中的通用性。在园艺作物研究中,梁芳芳等[34]发现黄瓜cpSSR引物在其近缘种甜瓜、南瓜和西瓜中的通用性较好;Xue等[35]研究了中国莲cpSSR在莲属中的通用性及多态性;Jiang等[36]从中国芒中开发出了在芒属植物中具有通用性的cpSSR引物。而在木本植物研究中,韩键等[37]研究揭示了果梅cpSSR引物在李属的桃、李、梅、杏、樱桃等5个树种中均具有通用性。
关于豆科植物的cpSSR分子标记研究鲜见报道。Angioi等[38]报道了所开发的菜豆属cpSSR标记的通用性,并分析了其在豆科的羽扇豆、花生、蒺藜苜蓿、鹰嘴豆、大豆和豌豆中的通用性。当前,对豇豆叶绿体基因组cpSSR分子标记的研究尚未见正式报道。
因此,本研究初步尝试对豇豆cpSSR分子标记进行研究,设计筛选了8对多态性豇豆cpSSR引物,并在豇豆的近缘种豌豆、利马豆和菜豆中成功扩增,表明了这些cpSSR引物在不同豆类物种之间具有较为广泛的通用性。总之,本研究分析了豇豆基因组微卫星的序列组成和分布特点,并成功开发出具有种间通用性的豇豆cpSSR引物,这将为豇豆cpSSR分子标记应用于豆科物种开展群体遗传学、系统发育与进化等研究奠定一定的基础。
[1]张静,彭海,陈禅友.分子标记在长豇豆遗传分析中的应用进展[J].长江蔬菜,2010(16):1-5.
[2]Fatokun C A,Danesh D,Young N D.Molecular taxonomic relationships in the genusVignabased on RFLP analysis[J].Theoretical and Applied Genetics,1993,86:97-104.
[3]Coulibaly S,Pasquet R S,Papa R,et al.AFLP analysis of the phenetic organization and genetic diversity ofVigna unguiculataL.Walp.reveals extensive gene flow between wild and domesticated types[J].Theoretical and Applied Genetics,2002,104:358-366.
[4]陈禅友,潘磊,胡志辉,等.长豇豆品种资源的RAPD分析[J].江汉大学学报:自然科学版,2008,36(4):77-83.
[5]Fana S B,Pasquet R S,Gepts P.Genetic diversity in cowpea[Vigna unguiculata (L.)Walp.]as revealed by RAPD markers[J].Genetic Resources and Crop Evolution,2004,51:539-550.
[6]Chen C Y,Peng H.Differentiation and phylogenetic relationship among different cultibars of asparagus bean(Vigna unguiculata (L.)ssp.sesquipedalis)assessed using ISSR markers[J].Nordic Journal of Botany,2010,28:251-256.
[7]Shaw J,Lickey E B,Schilling E E,et al.Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms:the tortoise and the hare III[J].American Journal of Botany,2007,94(3):275-288.
[8]Kongjaimun A,Kaga A,Tomooka N,et al.An SSR-based linkage map of yardlong bean (Vigna unguiculata (L.)Walp.subsp.unguiculataSesquipedalis Group)and QTL analysis of pod length[J].Genome,2012,55(2):81-92.
[9]Xu P,Wu X,Wang B,et al.Development and polymorphism ofVigna unguiculatassp.unguiculatamicrosatellite markers used for phylogenetic analysis in asparagus bean (Vigna unguiculatassp.sesquipedialis (L.)Verdc.)[J].Molecular Breeding,2010,25:675-684.
[10]Powell W,Morgante M,McDevitt R,et al.Polymorphic simple sequence repeat regions in chloroplast genomes:applications to the population genetics of pines[J].Proceedings of the National Academy of Sciences of the U-nited States of America,1995,99:7 759-7 763.
[11]李博,冯慧敏,王静毅,等.基于cpSSR分子标记的香蕉种质资源分类[J].果树学报,2011,28(6):1 012-1 018.
[12]赵丹,隋心,孙晓艳,等.基于cpSSR标记的红松天然群体自由授粉子代的父本分析 [J].经济林研究,2011,29(4):23-27.
[13]程丽莉,封海东,饶群,等.湖北省十堰地区野生板栗cpSSR 遗传多样性研究[J].果树学报,2012,29(3):382-386.
[14]邵丹,裴赢,张恒庆.凉水国家自然保护区天然红松种群遗传多样性在时间尺度上变化的cpSSR分析[J].植物研究,2007,27(4):473-474.
[15]张田,李作洲,刘亚令,等.猕猴桃属植物的cpSSR遗传多样性及其同域分布物种的杂交渐渗与同塑[J].生物多样性,2007,15(1):1-22.
[16]Bucci G,González-Martínez S C,Le Provost G,et al.Range-wide phylogeography and gene zones inPinus pinasterAit.revealed by chloroplast microsatellite markers[J].Molecular Ecology,2007,16(10):2 137-2 153.
[17]Yu H,Nason J D.Nuclear and chloroplast DNA phylogeography ofFicus hirta:obligate pollination mutualism and constraints on range expansion in response to climate change[J].New Phytologist,2013,197(1):276-289.
[18]Benson G.Tandem repeats finder:a program to analyze DNA sequences[J].Nucleic Acids Research,1999,27(2):573-580.
[19]李强,万建民.SSRHunter,一个本地化的SSR位点搜索软件的开发[J].遗传,2005,27(5):808-810.
[20]Weber J L.Informativeness of human (dC-dA)n,(dG-dT)n polymorphisms[J].Genomics,1990,7(3):524-530.
[21]Birky C W.Uniparental inheritance of mitochondrial and chloroplast genes:mechanisms and evolution[J].Proceedings of the National Academy of Sciences of the United States of America,1995,92:11 331-11 338.
[22]胡建斌,李建吾,梁芳芳,等.黄瓜叶绿体基因组全序列微卫星分布特征与标记开发[J].细胞生物学杂志,2009,31(1):69-74.
[23]李博,刘合霞.叶绿体基因组微卫星标记(cpSSR)研究进展[J].安徽农业科学,2012,40(13):7 638-7 639.
[24]Provan J,Powell W,Hollingsworth P M.Chloroplast microsatellites:new tools for studies in plant ecology and evolution[J].Trends in Ecology and Evolution,2001,16:142-147.
[25]王化坤,乔玉山,娄晓鸣,等.拟南芥叶绿体DNA全序列微卫星分布规律的分析[J].中国生物化学与分子生物学报,2006,22(10):845-850.
[26]张扬勇,方智远,王庆彪,等.拟南芥叶绿体SSR引物在甘蓝上的应用[J].园艺学报,2011,38(3):549-555.
[27]张君毅.蝴蝶兰叶绿体DNA微卫星分析与标记开发[J].江西农业学报,2011,23(12):31-33.
[28]Bausher M G,Singh N D,Lee S B,et al.The complete chloroplast genome sequence ofCitrus sinensis (L.)Osbeck var'Ridge Pineapple':organization and phylogenetic relationships to other angiosperms[J].BMC Plant Biology,2006,6:21.
[29]龚桂芝,洪棋斌,彭祝春,等.枳属种质遗传多样性及其与近缘属植物亲缘关系的SSR和cpSSR分析[J].园艺学报,2008,35(12):1 742-1 750.
[30]Desiderio F,Bitocchi E,Bellucci E,et al.Chloroplast microsatellite diversity inPhaseolus vulgaris[J].Frontiers in Plant Science,2012(3):312.
[31]Clegg M T,Gaut B S,Learn G H,et al.Rates and patterns of chloroplast DNA evolution[J].Proceedings of the National Academy of Sciences of the United States of America,1994,91(15):6 795-6 801.
[32]Ishii T,McCouch S R.Microsatellites and microsynteny in the chloroplastgenomes ofOryza and eightother Gramineae species[J].Theoretical and Applied Genetics,2000,100:1 257-1 266.
[33]Diekmann K,Hodkinson T R,Barth S.New chloroplast microsatellite markers suitable for assessing genetic diversity ofLolium perenneand other related grass species[J].Annals of Botany,2012,110(6):1 327-1 339.
[34]梁芳芳,李琼,胡建斌,等.黄瓜cpSSR引物的多态性与通用性[J].江西农业学报,2010,22(3):1-4.
[35]Xue J,Wang S,Zhou S L.Polymorphic chloroplast microsatellite loci inNelumbo (Nelumbonaceae)[J].American Journal of Botany,2012,99(6):240-244.
[36]Jiang J X,Wang Z H,Tang B R,et al.Development of novel chloroplast microsatellite markers forMiscanthus species (Poaceae)[J].American Journal of Botany,2012,99(6):230-233.
[37]韩键,王化坤,练春兰,等.果梅nSSR、cpSSR引物在李属果树上的通用性分析[J].果树学报,2009,26(6):830-835.
[38]Angioi S A,Desiderio F,Rau D,et al.Development and use of chloroplast microsatellites inPhaseolusspp.and other legumes[J].Plant Biology,2009,11(4):598-612.
猜你喜欢
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
南方医科大学学报(2022年3期)2022-04-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
铁道学报(2018年5期)2018-06-21
四川动物(2017年6期)2017-12-12
四川动物(2017年4期)2017-07-31
中国铸造装备与技术(2017年3期)2017-06-21
安徽电气工程职业技术学院学报(2016年4期)2017-01-15
