
P53siRNA对肺腺癌细胞A549中HMGN2表达的影响*
2014-12-05 06:36李洪敏冷言冰
成都医学院学报 2014年6期
李洪敏,王 晶,韩 琴△,刘 新,冷言冰,黄 宁
1.成都医学院 公共卫生系(成都 610083);2.四川大学华西医学中心 感染与免疫实验室(成都 610041)
HMGN2属于高迁移率非组蛋白家族成员,是动物细胞核中含量最丰富的非组蛋白家族,具有抗病毒、细菌和肿瘤等多种微生物活性。近年来发现,它在哺乳动物的发育包括胚胎发育方面起着重要作用,故HMGN2是一类具有多功能的分子[1]。P53蛋白是一种抑癌基因,目前对其在肿瘤方面的研究比较深入,而早在上个世纪末就发现P53与胚胎发育有着紧密联系,P53基因被敲除后的胚胎小鼠在出生后会因多器官衰竭而很快死亡。有报道[2]显示,与HMGN2同一家族成员的HMNB-1可作为一个独特的活化剂调节P53基因在信号通路中的变化,但P53基因能否调节其同源蛋白HMGN2的变化尚无报道。为了观察P53和HMGN2的关系,本研究利用RNA干扰(RNAi)技术靶向稳定沉默A549细胞中的突变型P53基因,观察其对细胞中HMGN2蛋白调节的变化,以探讨P53基因和HMGN2蛋白之间的关系,为今后开展小鼠生长过程中HMGN2参与发育机制的体内研究提供科学依据。
1 材料和方法
1.1 人肺腺癌细胞株及细胞培养
人肺腺癌细胞株A549由四川大学生物治疗重点实验室惠赠。P53-siRNA试剂盒和小鼠抗人P53抗体购至美国Santa cruz公司,胎牛血清(FBS)购自美国 HyClone公司。Trizol试剂盒、脂质体(lipofectamineTM-2000)购自美国Invitrogen公司,G418购至美国sallgon公司,小鼠抗人β-actin一抗和兔抗鼠IgG/HRP二抗购自北京中杉金桥生物技术有限公司。P53-siRNA真核细胞转染的人肺腺癌细胞株A549作为实验组P53-si,转染空质粒的A549细胞为空白对照组con-si和未转染任何载体的A549细胞为正常对照组con。各组细胞于含10%FBS的细胞培养液DMEM中、37℃、含5%CO2温箱中培养,每组细胞的培养基加入G418(终浓度为200μg/mL)处理。
1.2 小分子干扰引物序列
P53基因的寡核苷酸的P53-siRNA序列设计并合列。实验序列为靶向P53基因的寡核苷酸,其正 义 序 列 为:5′-CUACUUCCUGAAAACAAC GdTd T-3′,反义序列为:5′-CGUUGUUUUCAGG AAGUAGdTdT-3′;对照的β-actin的正义链为:5′-GCG CGCTTTGTAGGATTCG-3′,反 义 链:5′-GGTAC CACCATGTACCCAG-3′。由上海吉玛公司合成,经测序完全正确。
1.3 细胞转染和蛋白提取
转染方法按照lipofectamineTM-2000使用说明书进行。转染24h后在倒置荧光显微镜下观察绿色荧光表达情况,分别提取各个克隆细胞的总RNA和蛋白质进行鉴定,选出干扰效果最好的克隆进行保种。分别提取稳定转染PSG-P53i和PSG-P53c的细胞总蛋白。步骤如下:用PBS缓冲液洗涤培养瓶中的细胞两遍;用细胞刮棒刮细胞,收集细胞到离心管中,1 000r/min、4℃离心5min,去上清;加入适量蛋白裂解液(预冷),或者加三去污裂解缓冲液(50mol/L pH=8.0Tris-HCl,150mmol/L NaCl,0.1%SDS,100μg/mL 苯 甲 基 磺 酰 氟,1μg/mL Aprotinin,1%NP-40,0.5%去氧胆酸钠)40μL,超声粉碎3~4次,3~4s/次,在振荡器上强烈振荡;将裂解物在15 000r/min、4℃的条件下离心15min;将上清转移到另一个微量离心管中,-80℃冰箱保存。上述操作均于冰上进行,以避免蛋白降解。
1.4 Western blot检测P53及HMGN2蛋白的表达
提取的总蛋白经BAC法进行蛋白定量,取100 g蛋白100℃变性5min,10%SDS-PAGE电泳,将目的条带电转至PVDF膜上,5%脱脂奶粉室温封闭1h,-抗4℃孵育过夜,HRP标记的二抗室温孵育2h。用LAS-4000化学发光捕获系统显色,以βactin为内参,用ImageJ2X软件扫描灰度值进行半定量分析。
1.5 统计学方法
采用SPSS 16.0软件进行统计分析,计量资料以均数±标准差(±s)表示,多组均数比较采用单因素方差分析,两组间比较采用q检验,以P<0.05为差异有统计学意义。
2 结果
2.1 细胞转染结果
实验组和空白对照组细胞转染24h后,在蓝色荧光激发下,A为实验组带荧光的siRNA细胞中发出明亮的绿色荧光,荧光强度较强,B为空白对照组无荧光,提示质粒转染效率较好,转染效率达到90%以上(图1)。

图1 荧光显微镜检测转染的A549细胞效果图
2.2 P53及HMGN2蛋白的表达
Western blot检测结果显示,实验组P53-si中P53基因表达降低,正常对照con和空白对照组con-si中的P53蛋白表达无明显变化,表明P53-siRNA对人肺腺癌A549细胞P53蛋白含量下调(图2)。沉默A549细胞中的P53基因后,实验组中P53的表达率为62%,空白对照组为98%,达到了干扰效果。Western blot检测HMGN2的结果表明,实验组P53-si中HMGN2蛋白含量较正常对照组明显下调,空白对照组con-si中HMGN2的表达有所降低。HMGN2的表达率为:实验组56%,空白对照组88%(P<0.01)(图3)。
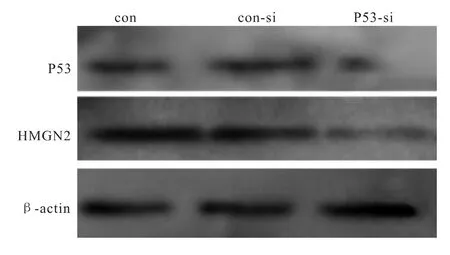
图2 各组细胞中P53和HMGN2蛋白的表达

图3 半定量P53和HMGN2蛋白在各组细胞中的表达含量
3 讨论
HMGN在胚胎发育的早期表达量极为丰富,对胚胎发育以及相关基因有整体调节作用[3]。Mohamed等[4]检测出HMGN2与胚胎发育的时相性有关,发现HMGN2的mRNA在大鼠胚胎和分化组织中高表达,而在成年大鼠的各组织器官中表达水平较低;将HMGN2的反义寡核苷酸注入大鼠单细胞受精卵中,检测到HMGN2蛋白在双细胞及四细胞阶段缺失,以及单细胞分裂成双细胞、双细胞分裂成四细胞胚胎会延迟。HMGN2可以作为组织或细胞在器官发生分化的标志,而对于这种分化的调节作用,具体表现为HMGN2对胚胎发育信号通路的调节作用和导致信号通路末端基因的变化[5]。有报道[2]称,HMGB-1作为其信号通路的调节因子是P53一个独特的活化剂。
P53蛋白是一种抑癌基因,定位于人类染色体17p13.1,被称为P53蛋白。目前对P53研究比较深入的是肿瘤方面[6],而早在上个世纪末就发现P53与胚胎发育有着紧密的联系,甚至包括P63/P73的整个P53家族在人体的发育过程起着重要的作用[7-8]。P53是细胞生长周期中的负调节因子,与细胞周期的调控、DNA修复、细胞分化和细胞凋亡等生物学功能有关,它可以和HMGN2同一家族成员HMGB-1中的C端结合,从而调节HMGB-1在胚胎发育中的作用。本实验显示,P53可下调A549细胞中的HMGN2表达,P53家族参与胚胎发生发育过程的功能已被肯定,HMGB-1蛋白在结构上分为两个HMG区域即A域和B域。Sourav等[2]和Rowell等[9]利用瞬态转染检测发现,C终端酸性区域和HMGB-1是促使体内P53激活的关键,并证明酸性C末端和P53能介导转录激活HMGB-1蛋白的A盒,并进一步影响下游的作用,可见P53在发育过程中的重要性,它能够作为调节因子促使HMGB-1蛋白在发育过程中发挥作用。
本研究通过小分子RNAi技术,利用siRNA沉默靶向基因P53,Western blot检测结果发现:A549细胞中HMGN2表达下调,表明P53基因可对HMGN2mRNA表达水平产生明显的抑制作用,HMGN2水平与P53呈正相关,这意味着P53基因的调控和HMGN2参与发生发育功能活性有着密切联系。根据实验结果推测,这种作用可能是P53作为HMGN2参与调节胚胎发育的激活因子,从而参与HMGN家族蛋白变化的调控途径之一。实验只通过基因沉默技术干扰了细胞中的P53基因,为下一步的动物实验研究提供了理论基础,对于动物体内P53基因是否也能参与调节HMGN2蛋白的变化有待进一步研究。
[1]Rattner BP,Yusufzai T,Kadonaga JT.HMGN proteins act in opposition to ATP-dependent chromatin remodeling factors to restrict nucleosome mobility [J].Mol Cell,2009,34(5):620-626.
[2]Sourav B,Tapas TK.The acidic C-terminal domain and A-box of HMGB-1regulates p53-mediated transcription [J].Nucleic Acids Research,2007,31(12):3236-3247.
[3]Deng LX,Wu GX,Cao Y,etal.The chromosomal protein HMGN2mediates the LPS-induced expression of betadefensins in mice[J].Inflammation,2012,35(2):456-473.
[4]Mohamed OA,Bustin M,Clarke HJ.High-mobility group proteins 14and 17maintain the timing of early embryonic development in the mouse [J].Dev Biol,2001,229(1):237-249.
[5]Cao Y,Wu G,Huang N.High mobility group nucleosomal binding domain 2protein protects bladder epithelial cells from Klebsiella pneumoniae invasion[J].Biol Pharm Bull,2011,34(7):1065-1071.
[6]Lane DP,Cheok CF,Lain S.p53-based cancer therapy[J].Cold Spring Harb Perspect Biol,2010,2(9):1222.
[7]Danilova N,Sakamoto KM,Lin S.p53family in development[J].Mech Dev,2008,125(11-12):919-931.
[8]罗兰,张闻,张彦,等.P53基因在小鼠睾丸发育过程中的表达规律[J].昆明理工大学学报,2011,36(2):56-61.
[9]Rowell JP,Simpson KL,Stott K.HMGB1-facilitated p53DNA binding occurs via HMG-Box/p53transactivation domain interaction,regulated by the acidic tail[J].Structure,2012,20(12):2014-2024.
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07
中日友好医院学报(2022年4期)2022-10-15
汽车维修与保养(2021年8期)2021-02-16
中国生殖健康(2020年5期)2021-01-18
看世界·学术下半月(2020年7期)2020-09-10
中国生殖健康(2018年5期)2018-11-06
广东农业科学(2017年10期)2018-01-25
中国医学创新(2017年22期)2017-11-15
工业设计(2016年4期)2016-05-04
新疆医科大学学报(2015年10期)2015-12-26
