
温度和pH对驼背鲈消化酶活力的影响*
2014-11-24 08:35刘江华区又君李加儿吴水清
中山大学学报(自然科学版)(中英文) 2014年3期
刘江华,区又君,李加儿,吴水清
(1.中国水产科学研究院南海水产研究所∥农业部南海渔业资源开发利用重点实验室,广东广州 510300;2.上海海洋大学水产与生命学院,上海 201306)
消化酶是由消化系统分泌的起营养和消化作用的酶类,因反应底物不同而分为蛋白酶、脂肪酶和淀粉酶等几种。消化酶活力是评价动物生理机能的重要指标之一,不同消化酶在动物机体中的分布随消化器官不同而不同;同一种消化酶在不同消化器官中的分布也存在明显差异;甚至同一消化器官中同一种消化酶,当外界环境因子改变时,其活力大小及分布也会发生改变。近年来,国内外对鱼类消化酶的研究已有较多报道[1-4],但关于外界环境因子对消化酶活力的影响的研究较少;温度和pH是消化酶活力两个重要环境因子,关于其对鱼类消化酶活力影响的报道,国内已对卵形鲳鲹 (Trachinotus ovatus)[5-6]、黄鳍鲷 (Sparus latus)[7-8]、洞庭鲶鱼 (Silnrus asotus)[9]、黑鲷 (Sparus macrocephalus)[10]、杂色鲍 (Haliotis diversicolor Reeve)等鱼类进行了大量研究[11]。
驼背鲈 (Cromileptes altivelis)俗名老鼠斑,隶属鮨科、石斑鱼亚科、驼背鲈属[12]。驼背鲈具有较高的食用和观赏价值,分布在西太平洋和印度洋的热带地区,我国见于南海[13]。至今国外对驼背鲈的研究处于起步阶段,国内的研究主要包括形态和生物学性状[14]、引种驯养及人工诱导性腺发育和繁殖[15]、胚胎发育[16]、年龄与生长特征[12]、染色体核型分析[17]、肾脏、鳃丝、嗅囊、肝脏及脑垂体的组织学结构等[13,18-21],关于驼背鲈消化系统的研究只见于对组织学与组织化学的观察[22],而对消化酶的研究尚未报道,因此本文通过温度和pH对驼背鲈消化酶活力的影响,为其营养生理和消化吸收机能的深入研究提供理论依据。
1 材料与方法
1.1 实验用鱼及驯养
实验用鱼共9尾,购自水产市场,体质量320.97~386.45 g,全长28.50~30.00 cm。实验前在水温24~25℃,盐度27‰~28‰,pH 7.8~8.2的条件下驯养2 d;每天换水一次,换水量为80%,换水温差≤1℃;连续充气并保持溶解氧≥5 mg/L。
1.2 温度和pH梯度实验设计
1.2.1 温度梯度实验设计 设7个温度梯度,在20~50℃范围内以5℃为一个梯度,在pH 7.2的条件下在恒温水浴锅内分别测定每一个温度下的酶活力。
1.2.2 pH梯度实验设计 在pH为2.2~9.2的范围内,以一个pH为幅度,设8个梯度,以37℃为反应温度,分别测定每一个pH下的酶活力。
1.3 粗酶液制备
将停食24 h后的实验鱼处死并迅速在冰盘上进行解剖,取胃、肠、幽门盲囊、肝等消化器官,剔除附着的脂肪,用预冷 (4℃)的双蒸水洗去内容物,用滤纸吸取表面的水分,称量,剪碎并加入10倍体积的预冷 (4℃)双蒸水,在冰浴的条件下用匀浆机高速匀浆,匀浆液放置于4℃的冰箱中静置1 h,然后用高速冷冻离心机离心 (0~4℃,10000 r/min)30 min,离心后的上清液为粗酶液,将粗酶液置于4℃的冰箱中保存备用。
1.4 酶活力测定
参照区又君等[5]和范春燕等[23]的方法,测定蛋白酶、脂肪酶、淀粉酶及蛋白浓度的试剂盒购自南京建成生物工程研究所。
1.4.1 蛋白酶 每毫克组织蛋白在37℃的条件下1 min内水解酪蛋白质产生1 μg酪氨酸为一个酶活力单位 (μg/min)。
1.4.2 脂肪酶 组织脂肪酶活力单位定义:在37℃条件下,每克组织蛋白在本反应体系中与底物反应1 min每消化 μmol底物为一个酶活力单位(μmol/min)。
1.4.3 淀粉酶 淀粉酶活力单位是组织中每毫克蛋白在37℃与底物作用30 min,水解10 mg淀粉定义为一个淀粉酶活力单位 (mg/min)。
1.4.4 蛋白浓度 采用考马斯亮兰蛋白测定试剂盒测定蛋白浓度。
1.5 数据的处理与分析
实验所得数据用统计分析软件SPSS PASW Statistics进行处理,用软件中的ANOVA和LSD分别进行差异显著性比较和多重比较,(P>0.05)不存在显著性差异。 (P<0.05)具有显著性差异,(P<0.01)具有极显著性差异。
2 结果
2.1 温度对消化酶活力的影响
2.1.1 温度对蛋白酶活力的影响 图1所示,在设定的温度20~50℃范围内,驼背鲈各消化器官的蛋白酶活力均随着温度的升高呈先升后降的趋势。胃、肠、幽门盲囊和肝都在40℃时酶活力达到最高值,20℃时酶活力最低。胃蛋白酶在40℃和45℃不存在显著性差异 (P>0.05),因此胃蛋白酶的最适温度为40~45℃;其他部位蛋白酶的最适温度均为40℃;各实验温度组的酶活力与该消化器官最适温度的酶活力存在显著性差异 (P<0.05)和极显著性差异 (P<0.01)。

图1 温度对驼背鲈蛋白酶活力影响Fig.1 Effects of temperature on protease activity in C.altivelis
2.1.2 温度对脂肪酶活力的影响 图2所示,驼背鲈各消化器官在20~50℃温度范围内的脂肪酶活力均随着温度的升高呈先升后降的趋势。胃脂肪酶的最适温度为35℃,50℃时酶活力最低;其他部位在20℃时酶活力最低,肠和肝的脂肪酶最适温度为40℃,幽门盲囊在40℃与45℃时的脂肪酶酶活力不存在显著性差异 (P>0.05),因此40~45℃为幽门盲囊脂肪酶的最适温度;各消化器官其他温度的脂肪酶活力与最适温度脂肪酶活力有显著性差异 (P<0.05)和极显著性差异 (P<0.01)。
2.1.3 温度对淀粉酶活力的影响 图3所示,驼背鲈胃淀粉酶的最适温度为40℃,其他部位均为35℃;胃和肝淀粉酶在20℃时最低,与最适温度之间存在显著差异 (P<0.05);肠和幽门盲囊淀粉酶在50℃时最低,与最适温度之间存在极显著性差异 (P<0.01)。在设定的温度范围内,各消化器官的淀粉酶活力均随着温度的升高呈先升后降的趋势。
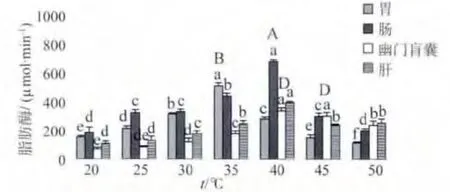
图2 温度对驼背鲈脂肪酶活力影响Fig.2 Effects of temperature on lipase activity in C.altivelis

图3 温度对驼背鲈淀粉酶活力影响Fig.3 Effects of temperature on amylase activity in C.altivelis
2.2 pH对消化酶活力的影响
2.2.1 pH对蛋白酶活力的影响 图4所示,在设定的pH 2.2~9.2范围内,驼背鲈各消化器官蛋白酶活力均随着pH的升高呈先升后降的趋势。胃蛋白酶的最适pH为3.2,肠、幽门盲囊和肝的最适pH均为8.2;pH 9.2时胃蛋白酶最低,其他部位的最低值均在pH 2.2;各部位的最适pH的酶活力与其他pH组的酶活力之间存在显著性差异 (P<0.05)和极显著性差异 (P<0.01)。
2.2.2 pH对脂肪酶活力的影响 图5所示,胃和肝脂肪酶的最适pH分别为7.2、8.2,肠和幽门盲囊的最适pH均为6.2,各部位在pH 2.2时酶活力最低;驼背鲈各消化器官脂肪酶活力在设定的pH范围内均随着pH的升高呈先升后降的趋势;最适pH酶活力与其他各pH组间存在显著性差异 (P<0.05)和极显著性差异 (P<0.01)。

图4 pH对驼背鲈蛋白酶活力影响Fig.4 Effects of pH on protease activity in C.altivelis

图5 pH对驼背鲈脂肪酶活力影响Fig.5 Effects of pH on lipase activity in C.altivelis
2.2.3 pH对淀粉酶活力的影响 图6所示,驼背鲈各消化器官淀粉酶活力在设定的pH 2.2~9.2范围内均随着pH的升高呈先升后降的趋势。肠、幽门盲囊和肝淀粉酶的最适pH为7.2;胃淀粉酶在pH 7.2和8.2时酶活力不存在显著性差异 (P>0.05),所以7.2~8.2为胃淀粉酶的最适pH;胃、肠和幽门盲囊最适pH酶活力与其他各组pH酶活力存在显著性差异 (P<0.05),肝的最适pH酶活力与最低酶活力pH2.2时之间存在极显著性差异(P<0.01)
2.3 最适温度下驼背鲈不同消化器官消化酶活力的分布比较
如图1-3所示,在最适温度条件下,各消化器官蛋白酶活力顺序依次为肠>胃>肝>幽门盲囊;肠最适温度酶活力显著高于胃 (P<0.05),极显著高于肝和幽门盲囊 (P<0.01);肝和幽门盲囊不存在显著性差异 (P>0.05)。脂肪酶最适温度酶活力顺序为肠>胃>肝>幽门盲囊,各部位之间存在显著性差异 (P<0.05)。最适温度下的淀粉酶活力高低顺序为肠>幽门盲囊>胃>肝,肠与幽门盲囊的最适温度淀粉酶活力不存在显著性差异 (P>0.05),肠、幽门盲囊与胃之间存在显著性差异 (P<0.05),胃与肝之间存在显著性差异(P <0.05)。

图6 pH对驼背鲈淀粉酶活力影响Fig.6 Effects of pH on amylase activity in C.altivelis
2.4 最适pH下驼背鲈不同消化器官消化酶活力的分布比较
图4所示,最适pH条件下各消化器官的蛋白酶活力顺序为肠>胃>幽门盲囊>肝;胃与幽门盲囊的最适酶活力不存在显著性差异 (P>0.05);肠的最适酶活力显著高于胃和幽门盲囊 (P<0.05),极显著高于肝 (P<0.01)。图5所示,各部位最适pH脂肪酶活力顺序为肠>胃>肝>幽门盲囊,各器官之间存在显著性差异 (P<0.05)。图6所示,在最适pH酶活力下的淀粉酶活力顺序为肠>幽门盲囊>胃>肝;肠与幽门盲囊之间不存在显著性差异 (P>0.05);肠和幽门盲囊与胃存在显著性差异 (P<0.05),与肝存在极显著性差异 (P<0.01)。
3 讨论
3.1 温度与驼背鲈消化酶活力的关系
本研究在离体条件下,测定驼背鲈消化酶的最适反应温度,在一定程度上能反映消化酶的热稳定性和温度对酶活力的影响规律。John[24]研究发现,鱼类消化酶的最适温度一般在30~50℃之间。黄鳍鲷胃、肠和肝在35~45℃时,3种主要消化酶都有较高活性[8]。卵形鲳鲹蛋白酶、脂肪酶和淀粉酶分别在40,40和35℃达到最高值[25]。韩庆等[9]报道了洞庭鲶鱼胃蛋白酶、脂肪酶和淀粉酶的最适温度分别为40,35,35℃,肠和肝胰脏这3种主要消化酶的最适温度均分别为45,35,30℃。本文对驼背鲈的研究结果显示,各消化器官消化酶的最适温度在35~45℃之间,与上述研究结果基本相同。酶与底物的反应随着温度的升高而加快,但超过一定温度范围后,酶受热变性占主要优势,反应速度反而随温度上升而减慢。本研究也表明,在20~50℃的温度范围内,驼背鲈消化酶的活力均随着温度的升高呈先升后降的趋势,其最适反应温度在35~45℃之间,并且高于生活的水体温度,而鱼类在最适反应温度下其消化酶活性达到最大值,此时机体会加快对营养物质的消化吸收,从而大大提高鱼类的生长速率。因此在实践生产过程中,要以驼背鲈最适消化酶温度为基础,在养殖水体温度范围内,应随着水温的升高时,相应的增加投饵量,加快驼背鲈的生长速度,降低养殖成本。
3.2 pH与驼背鲈消化酶活力的关系
pH也是影响酶活力的重要因子,改变酶分子的结构和改变酶与底物的结合是pH影响酶活力的两种主要途径。pH过酸或过碱都会改变酶分子的结构,甚至使酶变性失活,本研究结果显示,驼背鲈消化酶活力在设定的pH范围内均随着pH上升呈先升高后降低的趋势。大眼鰤鲈 (Stizostedion vireum)胃、肠、幽门盲囊和肝胰脏蛋白酶的最适pH 分别为3.0,8.0,8.0,7.0[26]。杂色鲍胃蛋白酶最适pH为2.6~3.4[11]。卵形鲳鲹胃蛋白酶的最适pH为3.2,肠、幽门盲囊和肝的最适pH为8.0[5]。本研究结果表明,驼背鲈胃蛋白酶的最适pH为3.2,肠、幽门盲囊和肝蛋白酶均为8.2,呈弱碱性,与上述结果一致。驼背鲈胃、肝脂肪酶的最适pH分别为7.2和8.2,肠、幽门盲囊均为6.2。而黄鳍鲷[7]和洞庭鲶鱼[9]的最适 pH 都为弱碱性,卵形鲳鲹[5]消化器官脂肪酶的最适pH均为6.2,呈弱酸性。本研究结果与上述研究结论相符,说明驼背鲈胃和肝脂肪酶在弱碱性条件下活力最强,而肠道和幽门盲囊脂肪酶在弱酸性条件下活力最强。目前对鱼类淀粉酶最适pH的研究已有不少。姜永华等[11]研究报道,杂色鲍消化道淀粉酶的最适pH为7.2。李俊辉等[27]报道了马氏珠母贝(Pinctada martensii)肝胰脏淀粉酶的最适pH为7.0。区又君等[5]研究表明,卵形鲳鲹胃、肠、幽门盲囊和肝淀粉酶的最适pH均呈弱碱性。本研究结果与上述结论一致。黄鳍鲷[7]的淀粉酶最适pH偏酸性,与本研究结果不同,这可能是因为淀粉酶的适宜pH因鱼类种间差异而有所不同,导致生活习性和摄食对象不同而对相应的酶的性质会有所改变。
3.3 驼背鲈不同消化器官消化酶活性的分布比较
不同酶在同一器官的温度和pH的敏感性不同,同一种酶在不同器官的温度和pH的敏感性也不同,所以不同器官中的消化酶活力肯定有所不同,这种不同对于控制机体内复杂的代谢途径具有重要的意义。驼背鲈蛋白酶在最适温度和pH下的酶活力顺序分别为肠>胃>肝>幽门盲囊、肠>胃>幽门盲囊>肝。鱼类的蛋白酶分为酸性和碱性,酸性蛋白酶主要位于胃内,肠和肝内以碱性蛋白酶为主;驼背鲈的肠蛋白酶活性明显高于胃蛋白酶,可见驼背鲈碱性蛋白酶占主要地位。Dask等[28]研究鱼类消化酶的分泌机制,发现肝胰脏是分泌蛋白酶原主要场所,但其蛋白酶活力微弱或甚至没有,并对肠道消化吸收蛋白酶起促进作用,说明了肠道的蛋白酶活力要高于肝胰脏,本实验结果与此理论相符。驼背鲈在最适温度和pH条件下,脂肪酶活力顺序为肠>胃>肝>幽门盲囊;李国希等[29]对黄鳍鲷消化酶分布的研究表明,脂肪酶活力的大小顺序为肠>胃>肝脏。本研究结果表明,肠脂肪酶活力在两种条件下均最大,其原因可能是氨基酸是脂肪酶的基本组成单位,而肠道是吸收营养物质的主要部位,从而说明肠道是吸收脂肪酶的重要场所。驼背鲈淀粉酶在最适温度和pH条件下酶活力顺序均为肠>幽门盲囊>胃>肝,鱼种类不同淀粉酶分泌的器官也不同,有的鱼类肝胰脏为唯一分泌器官,有些可能有辅助分泌器官;国外学者对鲤(Cyprinus carpio)[30]的研究也表明,大多数鱼类的肝脏是分泌淀粉酶原的主要场所,其淀粉酶活性却很低。本研究也得出驼背鲈肝脏的淀粉酶活性最低,原因可能是肝脏分泌淀粉酶后,淀粉酶吸附于肠道,因而检测到肠道的淀粉酶活性要远高于肝脏。本研究结果与卵形鲳鲹[31]和中国龙虾 (Panulirus stimpsoni Holthuis)[32]的结果有所差异,其主要原因还是不同种和不同食性鱼之间的消化酶活力存在差异,至于其他原因尚待做进一步研究。
[1]BABAEI S S,KENARI A A,NAZARI R,et al.Developmental changes of digestive enzymes in Persian sturgeon(Acipenser persicus)during larval ontogeny[J].Aquaculture,2011,318(1):138-144.
[2]UDDIN M,AHN H,KISHIMURA H,et al.Comparative study of digestive enzymes of squid(Todarodes pacificus)viscera after supercritical carbon dioxide and organic solvent extraction[J].Biotechnology and Bioprocess Engineering,2009,14(3):307-315.
[3]GAO Fei,YANG Hongsheng,XU Qiang,et al.Effect of water temperature on digestive enzyme activity and gut mass in sea cucumber Apostichopus japonicus(Selenka),with special reference to aestivation[J].Chinese Journal of Oceanology and Limnology,2009,27(4):714-722.
[4]吴莉芳,秦贵信,孙泽威,等.饲料中去皮豆粕替代鱼粉对埃及胡子鲇消化酶活力和肠道组织的影响[J].中山大学学报:自然科学版,2010,49(4):99-105.
[5]区又君,罗奇,李加儿.卵形鲳鲹消化酶活性的研究ⅢpH对幼鱼和成鱼消化酶活性的影响[J].海洋渔业,2010,32(4):417-421.
[6]区又君,罗奇,李加儿,等.卵形鲳鲹消化酶活性的研究Ⅳ养殖水温和酶反应温度对幼鱼酶活性的影响[J].海洋渔业,2011,33(1):28-32.
[7]李希国,李加儿,区又君.pH值对黄鳍鲷主要消化酶活性的影响[J].南方水产,2005,1(6):18-22.
[8]李希国,李加儿,区又君.温度对黄鳍鲷主要消化酶活性的影响[J].南方水产,2006,2(1):43-48.
[9]韩庆,刘良国,张建平,等.温度和pH对洞庭鲇鱼消化酶活性的影响[J].水生生物学报,2011,35(1):22-29.
[10]梅景良,马燕梅.温度和pH对黑鲷主要消化酶活性的影响[J].集美大学学报:自然科学版,2004,9(3):226-230.
[11]姜永华,颜素芬,严正凛.温度和pH对杂色鲍消化酶活力的影响[J].海洋科学,2012,36(2):11-18.
[12]区又君,廖锐,李加儿,等.驼背鲈的年龄与生长特征[J].水产学报,2007,31(5):624-632.
[13]蔡文超,区又君.驼背鲈肾脏显微和超微结构观察[J].中国水产科学,2008,15(5):773-778.
[14]区又君,李加儿,陈福华.驼背鲈的形态和生物学性状[J].中国水产科学,1999(1):25-27.
[15]区又君,李加儿,陈福华.驼背鲈引种驯养及人工诱导性腺发育和繁殖[J].湛江海洋大学学报,1999(3):20-23.
[16]区又君.驼背鲈的胚胎发育[J].海洋科学,2006,30(8):17-19.
[17]区又君,谢菁.驼背鲈的染色体核型分析[J].南方水产,2007,3(5):49-53.
[18]何永亮,区又君,蔡文超,等.驼背鲈鳃丝的光镜、扫描和透射电镜观察[J].华南农业大学学报:自然科学版,2009,30(2):86-89.
[19]区又君,蔡文超,李加儿.驼背鲈的嗅囊及嗅板细胞的形态学和组织学[J].海洋湖沼通报,2010(2):61-66.
[20]王静香,李加儿,区又君,等.驼背鲈肝脏结构的光镜和透射电镜观察[J].海洋渔业,2010,32(4):388-394.
[21]王静香,李加儿,区又君,等.驼背鲈脑垂体显微和超微结构的观察[J].中国水产科学,2011,18(5):1061-1068.
[22]区又君,勾效伟,李加儿.驼背鲈消化系统组织学与组织化学研究[J].海洋渔业,2011,33(3):289-296.
[23]范春燕,区又君,李加儿,等.卵形鲳鲹消化酶活性的研究Ⅴ大规格幼鱼消化酶活性在不同消化器官中的分布及盐度对酶活性的影响[J].海洋渔业,2011,33(4):423 -428.
[24]JOHN E H.Fish nutrition[M].California:Academic Press Inc,1987:332 -432.
[25]区又君,罗奇,李加儿,等.卵形鲳鲹消化酶活性的研究Ⅳ养殖水温和酶反应温度对幼鱼酶活性的影响[J].海洋渔业,2011,33(1):28-32.
[26]周景祥,张东鸣,黄权,等.大眼鰤鲈蛋白酶活性的研究[J].吉林农业大学学报,2000,22(3):107-110.
[27]李俊辉,王庆恒,杜晓东,等.温度和pH对马氏珠母贝肝胰脏消化酶活力的影响[J].水产科学,2011,30(2):115 -117.
[28]DAS K M,TRIPATHI S D.Studies on the digestive enzymes of grass carp,Ctenopharyngodon idella(Val.)[J].Aquaculture,1991,92:21-32.
[29]李希国,李加儿,区又君.黄鳍鲷主要消化酶活性在消化道不同部位的比较研究[J].海洋水产研究,2005,26(5):34 -38.
[30]UGOLEV A M,EGOROVA V V,KUZMINA V V,et al.Comparativemolecular characterization ofmembrane digestion in fish and mammals[J].Comp Biochem Physio l,1983,3(76B):627 -635.
[31]区又君,罗奇,李加儿,等.卵形鲳鲹消化酶活性的研究Ⅰ.成鱼和幼鱼消化酶活性在不同消化器官中的分布及其比较[J].南方水产科学,2011,7(1):50-55.
[32]姜永华,颜素芬.pH值对中国龙虾消化酶活力的影响[J].动物学报,2008,54(2):317-322.
猜你喜欢
中华胰腺病杂志(2022年4期)2022-08-23
小读者·阅世界(2022年3期)2022-05-05
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
医学前沿(2021年18期)2021-04-14
广西科技大学学报(2018年2期)2018-09-10
中国粮油学报(2017年6期)2017-07-19
意林·少年版(2017年7期)2017-05-08
中国洗涤用品工业(2017年2期)2017-04-16
故事作文·低年级(2016年10期)2016-09-10
