
广东廉江野生荔枝种群结构和分布格局
2014-11-03 04:23韦阳连欧阳勤森钟卫东卓书斌王雪兵黄小凤
亚热带植物科学 2014年1期
韦阳连,欧阳勤森,钟卫东,卓书斌,王雪兵,黄小凤
(东莞植物园,广东 东莞 523086)
作物野生近缘种或与之遗传关系较近的野生种,由于长期在自然逆境中生存,多演化为携带抗病、抗虫、抗逆性基因的重要载体,其丰富多样的遗传基因是作物品种改良及抗性品种选育的重要基础,因此,作物野生种质资源多样性的保护和利用日益受到关注[1]。荔枝Litchi chinensis原产于中国和越南,是我国广东、广西、海南和福建四省(区)种植业的支柱种类。荔枝的演化发展是由野生荔枝→半野生荔枝→栽培荔枝的过程,荔枝品种的改良创新最终需回归到最基础的野生荔枝种质资源研究及其保护与利用。野生荔枝是现有荔枝栽培品种的野生种,具有多种优良的园艺学性状、丰富的植物学性状和遗传多样性、很好的抵御生物和非生物胁迫的基因源以及土壤适应性[2],是进行荔枝品种改良、种质创新的重要基因库。我国海南、广西、广东等地野生荔枝林的发现不仅为证实荔枝起源于中国提供了强有力的证据,也为我国荔枝种质的改良创新注入新的血液[3]。
种群是构成群落的基本单位,研究种群结构可以了解种群现在的状态,有助于分析种群未来的数量动态;种群分布格局能反映种群空间变化规律,体现种群动态及其群落的演替趋势,对于揭示群落的多样性、稳定性及物种共存也具有重要意义[4]。我国现已开展的野生荔枝研究多集中在野生荔枝果实表型多样性、等位酶多样性及DNA水平遗传多样性研究[5-8],而关于野生荔枝群落、种群的研究还不多,仅有刘冰浩等[9]对广西玉林市博白县野生荔枝种群进行了研究。本文拟通过对广东廉江市谢鞋山野生荔枝种群结构和分布格局进行分析,为该省仅存的野生荔枝资源的保护提供依据。
1 研究地区和研究方法
1.1 研究地自然环境概况
调查样地位于广东省廉江市以东6 km的石城镇谢鞋山,地理位置110°20′E,21°36′N,最高海拔107.8 m,最低31.0 m,面积约60 hm2。成土母质为火山岩,林地土壤为砖红壤性红壤,有机质含量丰富,pH 4.5~5.5。廉江市属南亚热带季风气候,夏长冬暖,雨热同季,降水空间分布和季节分配不均,冬季寒潮入侵偶有严寒,夏秋期间台风、暴雨频繁,年均日照1745 h,年均温23.5 ℃,年均降水量1758.8 mm,4~9月降雨量占全年的83%。
谢鞋山现在所存留的植物群落为上世纪五十年代伐后的次生林。群落乔木层郁闭度70%~90%。优势树种是野生荔枝、山杜英、橄榄、乌榄、山乌桕等。灌木层由乔木种的幼树苗和灌木组成,灌木以九节、罗伞树、鸡屎树、毛果算盘子、柞木等占优势。草本层主要是露兜、淡竹叶、隐穗苔草、铁线蕨、肾蕨、剑叶凤尾蕨、芒萁等植物。
1.2 研究方法
1.2.1 样地调查 2011年6月,在谢鞋山野生荔枝分布相对集中的平缓区域(21°36′01″ N,110°20′43″E)共设置16个连续的10 m×10 m样方。对每个样方内的所有高于1.5 m木本植物进行每木调查,记录植物种类、数量、胸径等情况。
1.2.2 静态生命表编制 采用空间序列代替时间变化的方法,根据野生荔枝植株胸径大小分级,以立木级结构代替种群年龄结构,将野生荔枝种群按胸径(DBH)划分为9个径级,每级间隔5 cm,即第Ⅰ龄级为0<DBH≤5 cm,第Ⅱ龄级为5 cm<DBH≤10 cm,以此类推。根据特定时间生命表的编制原理和方法[4,10-11],通过各龄级的实际存活数ax和生命表各项参数之间的相互关系,编制野生荔枝的特定时间生命表,各参数计算方法为:lx= ax/a0×1000;dx= lx-lx+1;qx= dx/lx×100%;Lx= ( lx+lx+1) /2;;ex= Tx/lx。上述各式中,x为龄级;a0指标准化存活数为1000的龄级的实际存活数,一般指第xⅠ龄级的存活数;ax为x龄级存活的实际个体数;lx为从x龄级开始时的标准化存活数;dx为x龄级间隔期(x~x+1)的标准化死亡数;qx为各龄级的个体死亡率;Lx为x至x+1龄级期间存活的个体数;Tx为x龄级及以上龄级的个体总数;ex为进入x龄级开始时的平均生命期望或平均余年。
1.2.3 种群分布格局的测定 种群分布格局采用龚滨等[4]研究银缕梅种群的方法进行:根据采集的数据,采用方差(S2)/均值()比的t检验法进行分布格局判定。扩散指数C = S2/,其统计学的基础是在泊松分布中方差与均值相等。在泊松分布中,S2/值等于或接近于1,为随机分布;S2/值大于1,为集群分布;S2/值小于1,为均匀分布。采用t检验法来确定C实测值与数值“1”差异的显著性。t值通过公式t =计算,然后与t0.05(n-1)进行比较以确定其差异显著性。
2 结果与分析
2.1 野生荔枝种群静态生命表、龄级结构和存活曲线
根据16个10 m×10 m样方内各龄级野生荔枝的存活数,编制谢鞋山野生荔枝种群静态生命表(表1)。根据野生荔枝的径级大小,谢鞋山野生荔枝可分为9个龄级,其中低龄级和高龄级个体偏少,中龄级个体偏多。第Ⅰ龄级至第Ⅳ龄级的个体死亡率qx等于0或为负值,表明该阶段野生荔枝个体间的竞争压力不大,个体死亡率低;自第Ⅴ龄级开始,除第Ⅷ龄级外,qx均大于0,表明随着龄级的增大,林内光照、水分、养分和空间等因子已不能充分满足林内所有植株的要求,植株间竞争激烈,个体死亡率增加。期望寿命ex反应的是x龄级内个体的平均生存能力,由表1可知,ex在第Ⅰ、Ⅱ龄级较高,表明此阶段野生荔枝的生存质量较高,第Ⅲ龄级至第Ⅸ龄级普遍降低,这可能是在野生荔枝从低龄级向高龄级发展的过程中,野生荔枝个体间的种内竞争和群落不同物种间的竞争激烈,导致较高龄级野生荔枝的生存质量下降。
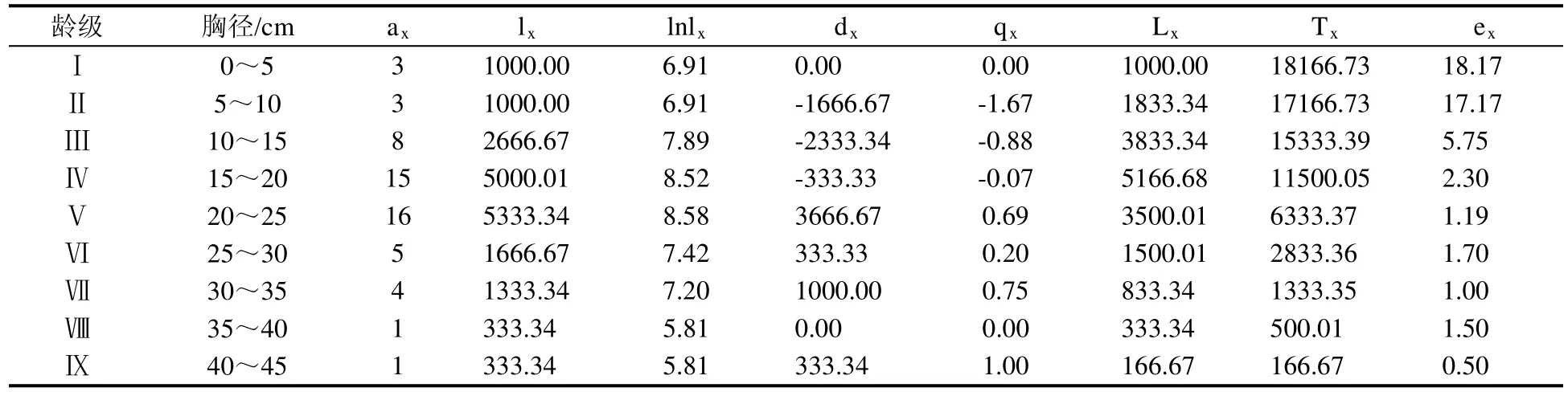
表1 谢鞋山野生荔枝种群静态生命表Table1 Static life table of wild Litchi chinensis population in Mt.Xiexie
以龄级为x轴,分别以各龄级实际存活数ax和各龄级标准化存活量的自然对数lnlx为y轴,绘制谢鞋山野生荔枝种群的龄级结构图和存活曲线图(图1、图2)。由图1可以看出,谢鞋山野生荔枝种群的龄级结构呈纺锤形,低龄级和高龄级个体偏少,中龄级个体偏多,表现为种群更新能力较差。此外,调查过程中还发现,乔木层内所有荔枝植株高≥5 m,灌木层和草本层很难看到野生荔枝幼苗,随着时间的推移、高龄级植株的死亡,幼苗更新较少势必导致种群变小。
由图2可知,野生荔枝种群的存活曲线在第Ⅰ龄级至第Ⅴ龄级呈现上升趋势,自第Ⅴ龄级后呈下降趋势,这可能是因为幼龄野生荔枝植株对光照、水分、空间等资源需求量不是很大,个体间生存竞争不激烈,种群内植株存活率高,使得小树有所积累,在第Ⅴ龄级,植株数量积累到最大值。自第Ⅵ龄级之后,此阶段的植株多为大树,对光照、水分、空间等资源的需求增加,个体间的生存竞争不断增大,植株存活率下降,使进入下一龄级的植株数量减少。综合而言,野生荔枝种群的存活曲线接近Deevey-Ⅰ型(即凸型),荔枝种群为衰退种群。
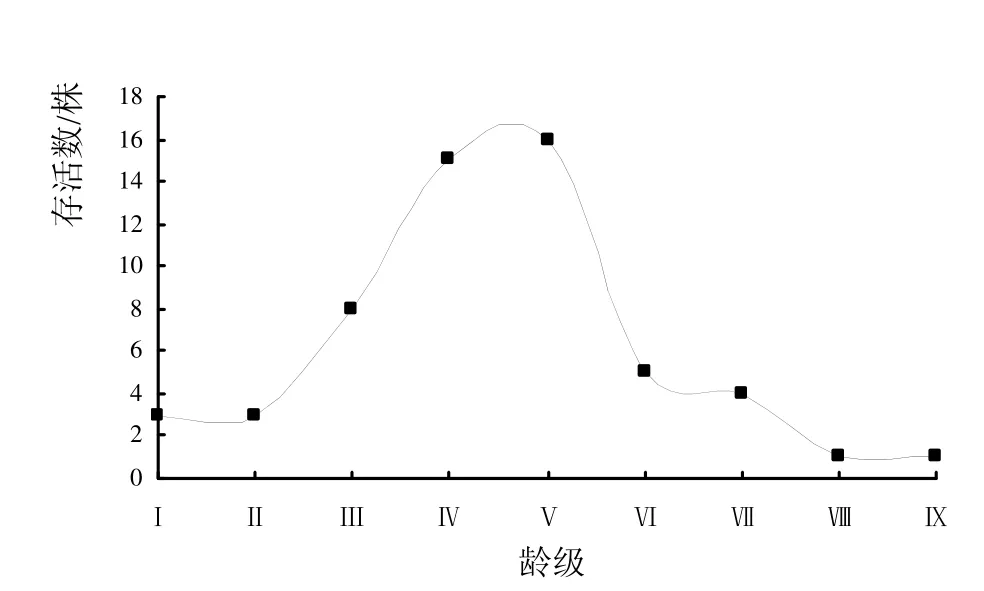
图1 谢鞋山野生荔枝种群的龄级结构Fig.1 The age structure of wild Litchi chinensis population in Mt.Xiexie
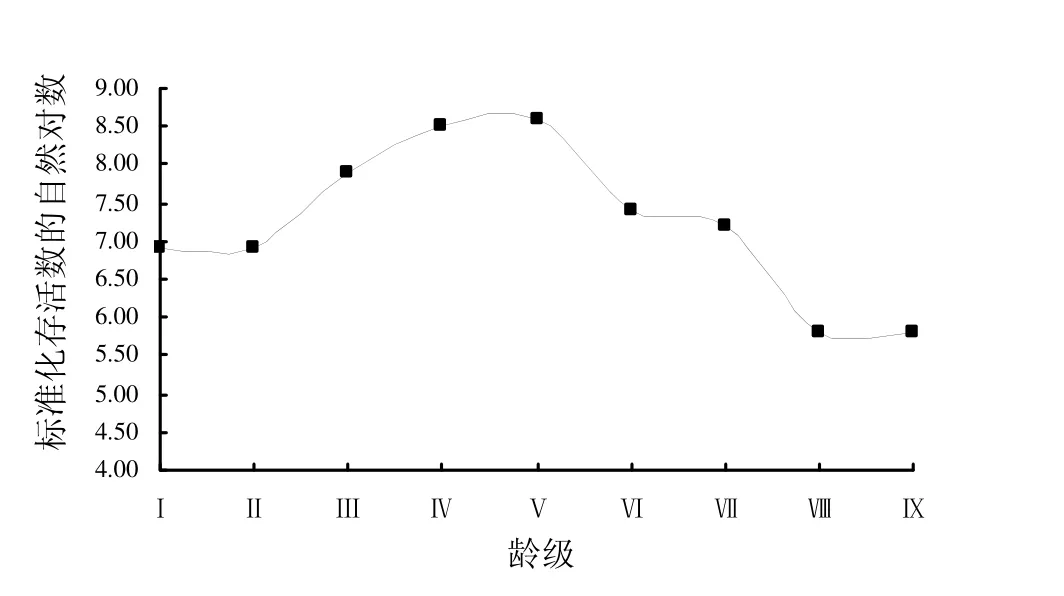
图2 谢鞋山野生荔枝种群的存活曲线Fig.2 Survival curve of wild Litchi chinensis population in Mt.Xiexie
2.2 野生荔枝种群空间分布格局
谢鞋山野生荔枝16个样方(n = 16)在泊松分布中的方差S = 2.062、均值= 3.733,扩散指数C = S2/= 0.552,其t检验实测值为t == -1.231,t0.05(15) = 2.131,∣t∣<t0.05,表明C的实测值与数值“1”的差异不显著,即C值接近1,谢鞋山野生荔枝种群属于随机分布。这是因为谢鞋山次生林闭郁度高,林下光线不足,导致灌草层植物个体生存竞争压力大,林下稀见荔枝树种的幼苗,荔枝树呈随机分布。
3 结论与讨论
上世纪五十年代大跃进时,广东廉江谢鞋山上生长数百年的野生荔枝树遭砍伐。现存野生荔枝林是在原来老树桩上重新发芽生长的。虽然是次生荔枝林,但其数量多,品种齐全。据当地居民介绍有些品种曾是贡品,亟待保护和利用。本研究显示,谢鞋山野生荔枝种群低龄级和高龄级个体偏少,中龄级个体偏多,呈随机分布,种群更新能力不足,种群的存活曲线接近Deevey-Ⅰ型,这与广西博白县野生荔枝种群的存活曲线一致[9],属衰退型种群,这是首次对谢鞋山野生荔枝资源的种群结构和分布格局作较为系统的报道,可为广东仅存的野生荔枝资源的合理保护和科学繁育提供理论依据。
随着资源保护意识的提高,廉江当地居民已开始自觉保护谢鞋山的野生荔枝资源,但野生荔枝林中还是很少见到荔枝幼苗,导致荔枝种群更新能力不足,呈衰退型,其原因可能与荔枝在自然状态下繁殖能力低及林内郁闭度高有关。首先,荔枝花芽分化对环境条件的要求较为苛刻,冬季日平均气温在11~14 ℃有利于花芽分化和花穗形成;日均温在14 ℃以上,温度越高、时间越长、营养梢小叶生长越迅速,消耗有机养分越多,花穗发育不良;温度l8~19 ℃可形成带叶花穗,20~25 ℃时花的原始体消失[9],而廉江冬季日平均气温是16~23 ℃,不利于花穗发育。其次,廉江每年4月份降雨量较多,此时恰逢荔枝开花,影响荔枝花药开裂和传粉昆虫的活动,使花药腐烂并导致霜疫霉病发生;雨水多还导致荔枝花粉和柱头粘液下降,不利于花粉管萌发,萌芽力剧减,甚至失去活性;此外,廉江夏季台风、暴雨频繁,造成荔枝落果严重、坐果率低,这些都影响了野生荔枝种群的繁育更新[9]。最后,林内郁闭度高,林内灌草层光照不足,野生荔枝种子在弱光下即使能萌发成幼苗,也会因光照不足无法进行光合作用而变得纤弱,最终失绿死亡。
针对野生荔枝种群的生存现状,建议采取如下保护措施:(1)开展谢鞋山野生荔枝资源普查,进行品种评价和归类;(2)收集野生荔枝种子,在林缘光线充足区域播种繁殖,进行就地保护;(3)收集野生荔枝品种资源,在周边气候条件相似的地区建立种质资源圃,对各类型的野生资源进行迁地保护;(4)开展野生荔枝资源的开发利用研究,使优良种质资源在推广应用中得到有效保护和发展。
[1] 侯向阳,高卫东.作物野生近缘种的保护与利用[J].生物多样性, 1999, 7(4): 327-331.
[2] 王艳琼,陈业渊.荔枝种质资源研究进展[J].江西农业学报, 2008, 20(7): 54-57.
[3] 吴淑娴.中国果树志( 荔枝卷)[ M] .北京: 中国林业出版社,1998:1-30.
[4] 龚滨,夏洋洁,张光富,陆芸,孙国.中国特有珍稀濒危物种银缕梅种群结构和空间格局[J].生态与农村环境学报,2012,28(6): 638-646.
[5] 傅玲娟,袁沛元.广东海南野生荔枝多种类型的发现[J].中国果树, 1983 (4) :17 -19.
[6] 姜成东,蔡胜忠,肖翔,肖小军,卢业凌.海南两个自然保护区野生荔枝遗传多样性研究[J].中国农学通报, 2009,25(9): 282-286.
[7] 姚庆荣.用SSR标记对中国荔枝(Litchi chinensis Sonn)野生种质资源的遗传多样性分析[D].兰州: 甘肃农业大学硕士学位论文, 2004: 30-39.
[8] 李明芳.荔枝 SSR标记的研究及其对部分荔枝种质的遗传多样性分析[D].儋州: 华南热带农业大学博士学位论文,2003: 50-51.
[9] 刘冰浩,朱建华,潘丽梅,彭宏祥.广西野生荔枝博白种群生命表分析[J].果树学报, 2010,27(3): 445-448.
[10] 李先琨,向悟生,唐润琴.濒危植物元宝山冷杉种群生命表分析[J].热带亚热带植物学报, 2002,10(1): 9-14.
[11] 陈少瑜,付玉嫔,吴涛,司马永康,郝佳波,韩燕.濒危植物大果木莲种群格局及濒危原因分析[J].植物资源与环境学报,2012,21(2): 102-106.
猜你喜欢
科学大众(2022年23期)2023-01-30
湖南生态科学学报(2022年4期)2022-11-11
农业科技与信息(2022年12期)2022-08-09
河南农业大学学报(2020年2期)2020-05-22
中国(俄文)(2019年8期)2019-08-24
山西林业科技(2019年1期)2019-05-27
意林·全彩Color(2019年4期)2019-05-11
运动(2017年1期)2017-09-03
湖北农业科学(2017年11期)2017-07-13
岭南音乐(2017年2期)2017-05-17
