
生物节律基因Timeless的生物学功能研究进展
2014-10-27 09:04宋何煜张耀江红
生物技术通讯 2014年3期
宋何煜,张耀,江红
北京交通大学 理学院生命科学与生物工程研究院,北京 100044
生物节律是生物体普遍存在的生命现象,参与多种生命活动的调控。目前在哺乳动物中发现有10种节律基因,分别是Per1、Per2、Per3、Cry1、Cry2、Clock、Bmal1、Timeless、NPAS2和CK1ε。生物节律是由几种核心节律基因通过转录-翻译反馈环控制和维持的,反馈环由正激活因子(Clock和Bmal1)、负效应因子(Per1、Per2、Per3、Cry1、Cry2和Timeless)和调节因子(CK1ε)组成[1]。Timeless是生物节律和细胞周期的直接连接者,同时参与ATR-Chk1和ATMChk2两大DNA损伤修复通路[2],在肺和肾的胚胎发育过程中发挥重要作用[3]。并且,在多种癌组织中检测到Timeless表达水平与癌旁非癌组织相比明显不同[4-10],提示它可能和癌症的发生和预后密切相关。我们简要总结了Timeless在生物节律、细胞周期、胚胎发育、癌症发生发展和诊疗中的生物学功能研究进展。
1 Timeless基因的结构
Timeless是Michael等在筛选影响果蝇生物节律的突变体时发现的,位于果蝇第2多线染色体的左臂,dpp和dp基因之间[11]。不同物种中都有Timeless同源基因的存在(表1)。人类的Timeless基因位于第12号染色体,全基因长约43 kb,编码由1208个氨基酸残基组成的蛋白TIMELESS,该蛋白在细胞质中形成后,与PER家族成员形成二聚体入核[12]。
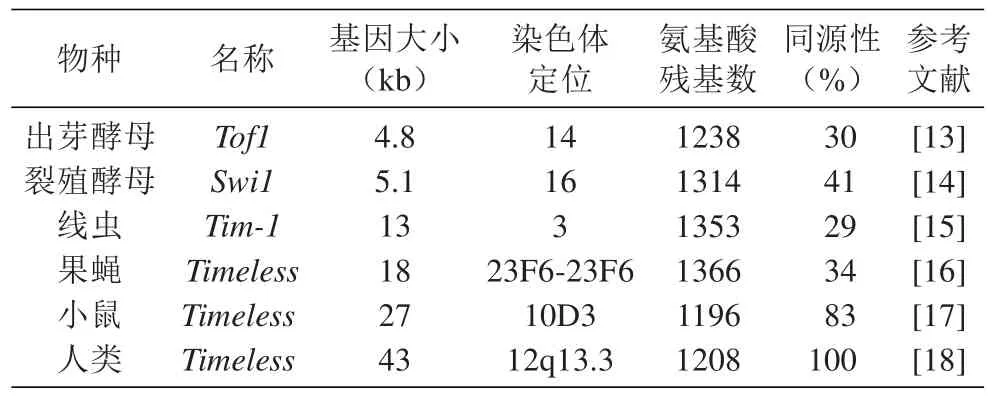
表1 不同生物Timeless基因的信息
2 Timeless在生物节律中的功能
生物节律是生物的一种生理和行为的周期性振荡[19],大部分生物的节律周期为24 h;在不同物种的生物节律活动中,Timeless的作用各不相同。
在果蝇中,当CLOCK-CYCLE二聚体激活Per和Timeless的表达时,新的节律周期开始。在黑暗条件下,PER-TIMELESS二聚体入核后,抑制CLOCKCYCLE的活性,降低其表达水平,从而终止节律周期[20]。光照可以激活光受体CRY,它和Timeless结合后引起TIMELESS的泛素化降解,节律周期发生转换[21]。蛋白激酶CK2可以对TIMELESS进行磷酸化修饰,增加细胞质中Timeless的表达水平,从而缩短节律周期[21]。在小鼠视交叉上核,下调Timeless表达水平会使Per1、Per2和Per3表达水平降低,而Cry1和Cry2表达水平升高[22],Per1和Per2表达水平降低可使节律周期的振幅降低[23],Cry是Per的负调控因子,缺失Cry1和Cry2的小鼠会出现节律紊乱[24]。细胞周期激酶CHEK1可以和Timeless及CK2发生相互作用,CHEK1表达水平降低可使小鼠的节律周期至少缩短1 h[25],而下调DNA解旋酶结合蛋白CDH4的表达水平可以延长节律周期。
3 Timeless在细胞周期中的功能
细胞周期的顺利进行依赖于细胞周期检测点机制的存在,其中的DNA损伤检测点能够延迟和阻止细胞分裂,主要有ATM(ataxia telangiectasia mutat⁃ed)依赖的 Chk2和 ATR(ataxia telangiectasia and Rad3-related)依赖的Chk1等2条DNA损伤信号通路[2]。出芽酵母Tof1基因和裂殖酵母Swi1基因可以激活DNA损伤检测点,同时,Swi1可以阻止复制叉解体[19]。Tim-1是线虫染色体凝聚的必需因子,在DNA复制中扮演着重要的角色,它的缺失会引起线虫胚胎死亡。基于对线虫、果蝇和小鼠的TIMELESS同源物的比较,Barbara等[26]认为TIMELESS样蛋白保守的功能不仅仅是生物节律调节,同时调控染色体的凝聚。在小鼠和人类细胞中,Timeless和复制叉共同移动,它是复制子复合体的成员之一,它和Tip⁃in的相互作用可以稳定复制叉和促进姊妹染色单体凝聚[27]。Timeless或一些蛋白(如Hus1、claspin、Chk1和Tipin)的失活可以逆转紫外线(UV)对DNA合成的抑制作用[28]。当DNA损伤发生时,Tim-Tipin复合体启动UV介导的S期细胞周期检测点,下调Time⁃less会抑制S期细胞周期检测点的激活,这些正好说明在小鼠Timeless缺失会引起胚胎致死现象[29]。虽然端粒长度的维护依赖于端粒酶,但Timeless和Shelterin家族成员TRF1和TRF2相互作用,在端粒的DNA重复区域阻止复制叉降解,促进DNA复制,进而维护端粒长度,保证染色体完整性的维持[30]。
当DNA复制被UV或羟基脲(HU)干扰后,抑制ATR-Chk1通路中蛋白的表达,促进早熟染色质凝聚(prematurechromosomecondensation,PCC)[19]。小鼠[31]和人类[2]的Timeless和Chk1相互作用,介导复制检测点信号从ATR向Chk1传递,而Timeless的缺失也会引起PCC,提示Timeless参与了ATR-Chk1依赖的复制检测点调控。阿霉素(doxorubicin,Dox)是一种治疗癌症的常用药物,能够激活ATM-Chk2反应通路,使细胞周期阻滞在G2/M期。人类Time⁃less不能激活ATM,但能激活ATM依赖的Chk2,下调Timeless使肿瘤细胞对Dox的细胞毒性更敏感[2],提示降低Timeless表达水平的药物可以作为提高Dox抗瘤效果的增敏剂。
4 Timeless在小鼠组织胚胎的发育中的作用
Timeless对小鼠胚胎发育至关重要,它在器官发生的初期广泛表达于大脑、肺、心脏、肝、胃和尿生殖嵴等[3],如果缺失可导致胚胎死亡率升高[32]。在胚胎的肺、肝、肾及神经上皮的发育过程中,Timeless表达水平较高,对上皮形成具有重要作用[33]。在9.5 d的胚胎,肺开始形成,Timeless存在于上皮细胞和间充质细胞中;在15 d胚胎的呼吸道上皮中Timeless仍维持较高的表达水平,但在间充质中的表达水平降低。Northern印迹结果也提示与18 d的小鼠胚胎和成体小鼠相比,Timeless在15 d的小鼠胚胎中表达水平最高。在肺发育过程中,Timeless表达水平的动态变化对肺的发育是至关重要的[3]。
在分支形成的器官中,Timeless基因的功能高度保守[3]。在肾发育过程中,输尿管芽分支形成时,Timeless是一种分化表达序列,在胚胎中的表达方式与其在成体中的表达不同。Timeless表达水平降低会抑制肾的胚胎发育,提示在肾发生和其他上皮器官形成中有重要作用[33]。
5 Timeless在肿瘤发生、发展和诊疗中作用的研究进展
5.1 肿瘤的发生和发展
恶性肿瘤的形成是一个涉及多因素多步骤的复杂过程。多项研究表明,大鼠和人类肿瘤可能是一种节律失调相关疾病。已经在多种肿瘤(如乳腺癌、卵巢癌、子宫内膜癌、前列腺癌和血液系统肿瘤等)中发现生物节律分子的遗传或功能性异常,这种异常的表现可能与癌症或其他疾病的发生有重要的相关性。与对应的癌旁非癌组织相比,Timeless在不同癌组织中的表达水平明显不同。Timeless在肺癌中mRNA和蛋白表达水平[7]以及结直肠癌中的mRNA表达水平[9]均升高,而肾癌和胰腺癌中mRNA表达水平降低,肝癌中的mRNA和蛋白水平表达也同样下降[4,6,10](表2)。不同文献对乳腺癌中Timeless表达水平的报道不一致,Yong等的研究提示乳腺癌组织中Timeless的mRNA表达水平升高[8],而Jan-Gowth等则认为乳腺癌组织中Timeless蛋白表达水平降低[5],结果不一致的原因可能与样品来源和检测方法不同有关,同时基因的表达受不同水平的诸多因素调控。
Timeless在癌组织和癌旁非癌组织中的差异表达和肿瘤的预后具有相关性。肝癌中Timeless表达水平的降低和肿瘤的分级相关[4]。肺癌中Timeless高表达的患者整体生存时间明显缩短,但与肿瘤分期和组织学分型无关,Yoshida认为Timeless可以作为肺癌诊断和预后的标记物,对肺癌的治疗效果有提示价值[7]。在结直肠癌中Timeless的mRNA表达水平和TNM分期关联密切,在TNM的Ⅲ~Ⅳ阶段,Time⁃less的mRNA表达水平达到最高,这种升高可能和结直肠癌的发生有关,也可能与对辅助化疗反应的减弱有关[9]。Yong等在80例乳腺癌组织和80例癌旁非癌组织中发现Timless启动子发生甲基化,并且甲基化程度在乳腺癌的Ⅱ、Ⅲ、Ⅳ期显著降低,这和Timeless在乳腺癌组织中高表达是一致的[8];而Jan-Gowth等在乳腺癌组织中未发现Timeless的启动子甲基化现象[5],同样,在子宫内膜癌[34]、肝癌[4]中也得到相似的结果。在肺癌细胞NCI-H157和NCIH460中,如果下调Timeless的表达水平,可以抑制细胞增殖和克隆形成,同时诱导细胞凋亡[7],提示它可以作为肺癌诊断和预防的标志物,同时也是具有较高治疗价值的靶分子。
5.2 肿瘤的诊疗
鉴于Timeless连接了生物节律和细胞周期两大调控系统,一些关于Timeless与癌症易感性的遗传学和表观遗传学研究成为当下热门课题。单核苷酸多态性(single nucleotide polymorphism,SNP)在人类基因组中广泛存在,且分布具有高密度性和高保守性。研究发现,一些基因的SNP与肿瘤发生、预后和药物敏感性相关,是一种便于利用的生物标志物。在441例乳腺癌组织和479例癌旁非癌组织中,Timeless的2个SNP位点(rs2291738和rs7302060)与乳腺癌的发生密切相关[8]。而Kristan[35]和Yong等[1]分别发现,在乳腺癌组织和前列腺癌组织中所检测的Timeless SNP位点与肿瘤的易感性和恶性程度无关。

表2 Timeless在各种癌组织中的差异表达及其预后相关性
节律调节是人类抵制癌症的重要先决条件。在小鼠和大鼠的实验模型中,用药时间的变化会给30多种抗癌药物的毒性和功效带来约百分之五十的差异[36]。雌激素受体α(ERα)的表达水平可用来筛选对激素类药物(如它莫西芬,tamoxifen)反应的乳腺癌患者,在体外实验中,Timeless的表达水平受17β雌二醇调控,并且可以在耐它莫西芬细胞株中过表达。Timeless有望成为筛选对它莫西芬耐药的ERα阳性乳腺癌患者的标记物[37]。通过检测肿瘤中Timeless基因的突变,可以预测药物敏感性的变化[9];同时,减低Timeless的表达水平可以增强一些化疗药物的活性,这些药物能够激活肿瘤细胞的DNA损伤通路,提示Timeless可能是一种新的治疗癌症的药物靶分子[2]。
6 结语
Timeless是重要的生物节律基因,在生物节律、细胞周期、胚胎发育、癌症发生发展和诊疗中发挥重要的生物学作用,如何将这些生物学功能用于临床尚有许多问题需要解决。Timeless在癌组织和癌旁非癌组织中的差异表达特点是否与癌症的组织类型和发病地区有关?是否在体外和体内降低Timeless的表达水平都能够抑制肿瘤细胞增殖、并诱导其凋亡?Timeless的SNP位点对于肿瘤易感性和治疗效果的预测是否具有实用性?随着研究数据的不断累积和丰富,期望能够阐明Timeless表达水平与肿瘤发生发展和预后的联系,明确其中的作用机制,充分发挥Timeless作为肿瘤诊断和治疗的靶分子的作用。
[1]Zhu Y,Stevens R G,Hoffman A E,et al.Testing the circadi⁃an genehypothesis in prostatecancer:apopulation-based case-control study[J].Cancer Res,2009,69(24):9315-9322.
[2]Yang X,Wood P A,Hrushesky W J.Mammalian TIMELESS is required for ATM-dependent CHK2 activation and G2/M checkpoint control[J].J Biol Chem,2010,285(5):3030-3034.
[3]Xiao J,Li C,Zhu N L,et al.Timeless in lung morphogenesis[J].Dev Dyn,2003,228(1):82-94.
[4]Lin Y M,Chang J H,Yeh K T,et al.Disturbance of circadi⁃an gene expression in hepatocellular carcinoma[J].Mol Car⁃cinog,2008,47(12):925-933.
[5]Kuo S J,Chen S T,Yeh K T,et al.Disturbance of circadi⁃an gene expression in breast cancer[J].Virchows Arch,2009,454(4):467-474.
[6]Mazzoccoli G,Piepoli A,Carella M,et al.Altered expression ofthe clock genemachinery in kidney cancerpatients[J].Biomed Pharmacother,2012,66(3):175-179.
[7]Yoshida K,Sato M,Hase T,et al.TIMELESS is overex⁃pressed in lung cancerand itsexpression correlateswith poor patient survival[J].Cancer Sci,2013,104(2):171-177.
[8]Fu A,Leaderer D,Zheng T,et al.Genetic and epigenetic as⁃sociations of circadian gene TIMELESS and breast cancer risk[J].Mol Carcinog,2012,51(12):923-929.
[9]Mazzoccoli G,Panza A,Valvano M R,et al.Clock gene ex⁃pression levels and relationship with clinical and pathological features in colorectal cancer patients[J].Chronobiol Int,2011,28(10):841-851.
[10]Relles D,Sendecki J,Chipitsyna G,et al.Circadian gene ex⁃pression and clinicopathologic correlates in pancreatic cancer[J].J Gastrointest Surg,2013,17(3):443-450.
[11]刘仲敏,张亚平.Timeless与生物钟基因[J].动物学研究,2001,22(6):497-501.
[12]Panda S,Hogenesch J B,Kay S A.Circadian rhythms from flies to human[J].Nature,2002,417(6886):329-335.
[13]Philippsen P,Kleine K,Pöhlmann R,et al.The nucleotide se⁃quence of Saccharomyces cerevisiae chromosome XIV and its evolutionary implications[J].Nature,1997,387(6632 Suppl):93-98.
[14]Bussey H,Storms R K,Ahmed A.The nucleotide sequence of Saccharomyces cerevisiae chromosome XVI[J].Nature,1997,387(6632 Suppl):103-105.
[15]C.eleganssequencing consortium.Genome sequence ofthe nematode C.elegans:a platform for investigating biology[J].Sci⁃ence,1998,282(5396):2012-2018.
[16]Hoskins R A,Carlson J W,Kennedy C,et al.Sequence fin⁃ishing and mapping of Drosophila melanogaster heterochromatin[J].Science,2007,316(5831):1625-1628.
[17]O'Reilly L P,Watkins S C,Smithgall T E.An unexpected role for the clock protein timeless in developmental apoptosis[J].PLoS One,2011,6(2):e17157.
[18]Dheekollu J,Chen H S,Kaye K M,et al.Timeless-depen⁃dent DNA replication-coupled recombination promotes Kapo⁃si's Sarcoma-associated herpesvirus episome maintenance and terminal repeat stability[J].J Virol,2013,87(7):3699-3709.
[19]Unsal-Kaçmaz K,Mullen T E,Kaufmann W K,et al.Cou⁃pling of human circadian and cell cycles by the timeless pro⁃tein[J].Mol Cell Biol,2005,25(8):3109-3116.
[20]Kaushik R,Nawathean P,Busza A,et al.PER-TIM interac⁃tions with the photoreceptor cryptochrome mediate circadian temperature responses in Drosophila[J].PLoS Biol,2007,5(6):e146.
[21]Meissner R A,Kilman V L,Lin J M,et al.TIMELESS is an important mediator of CK2 effects on circadian clock func⁃tion in vivo[J].J Neurosci,2008,28(39):9732-9740.
[22]Barnes J W,Tischkau S A,Barnes J A,et al.Requirement of mammalian Timeless for circadian rhythmicity[J].Science,2003,302(5644):439-442.
[23]Zheng B,Larkin D W,Albrecht U,et al.The mPer2 gene en⁃codesa functionalcomponentofthemammalian circadian clock[J].Nature,1999,400(6740):169-173.
[24]Gauger M A,Sancar A.Cryptochrome,circadian cycle,cell cycle checkpoints,and cancer[J].CancerRes,2005,65(15):6828-6834.
[25]Wallach T,Schellenberg K,Maier B,et al.Dynamic circadi⁃an protein-protein interaction networks predict temporal organi⁃zation of cellular functions[J]. PLoS Genet, 2013,9(3):e1003398.
[26]Chan R C,Chan A,Jeon M,et al.Chromosome cohesion is regulated by a clock gene paralogue TIM-1[J].Nature,2003,423(6943):1002-1009.
[27]Leman A R,Noguchi C,Lee C Y,et al.Human Timeless and Tipin stabilize replication forks and facilitate sister-chro⁃matid cohesion[J].J Cell Sci,2010,123(Pt 5):660-670.
[28]Kaufmann W K.Initiating the uninitiated:replication of dam⁃aged DNA and carcinogenesis[J].Cell Cycle,2007,6(12):1460-1467.
[29]Unsal-Kaçmaz K,Chastain P D,Qu P P,et al.The human Tim/Tipin complex coordinates an Intra-S checkpointre⁃sponse to UV that slows replication fork displacement[J].Mol Cell Biol,2007,27(8):3131-3142.
[30]Leman A R,Dheekollu J,Deng Z,et al.Timeless preserves telomere length by promoting efficient DNA replication through human telomeres[J]. CellCycle,2012,11(12):2337-2347.
[31]Smith K D,Fu M A,Brown E J.Tim-Tipin dysfunction cre⁃ates an indispensible reliance on the ATR-Chk1 pathway for continued DNA synthesis[J].J Cell Biol,2009,187(1):15-23.
[32]Gotter A L,Manganaro T,Weaver D R,et al.A time-less function for mouse timeless[J].Nat Neurosci,2000,3(8):755-756.
[33]Li Z,Stuart R O,Qiao J,Pavlova A,et al.A role for Time⁃less in epithelial morphogenesis during kidney development[J].Proc Natl Acad Sci USA,2000,97(18):10038-10043.
[34]Shih M C,Yeh K T,Tang K P,et al.Promoter methylation in circadian genes of endometrial cancers detected by methyla⁃tion-specific PCR[J].Mol Carcinog,2006,45(10):732-740.
[35]Grundy A,Schuetz J M,Lai A S,et al.Shift work,circadian gene variants and risk of breast cancer[J].Cancer Epidemiol,2013,37(5):606-612.
[36]Mormont M C,Levi F.Cancer chronotherapy:principles,appli⁃cations,and perspectives[J].Cancer,2003,97(1):155-169.
[37]Tozlu-Kara S,Roux V,Andrieu C,et al.Oligonucleotide mi⁃croarray analysis of estrogen receptor alpha-positive postmeno⁃pausal breast carcinomas:identification of HRPAP20 and TIME⁃LESS asoutstanding candidate markersto predictthe re⁃sponse to tamoxifen[J].J Mol Endocrinol,2007,39(4):305-318.
猜你喜欢
现代苏州(2022年9期)2022-05-26
文萃报·周五版(2022年9期)2022-03-11
国际呼吸杂志(2019年4期)2019-03-12
计算机测量与控制(2017年6期)2017-07-01
中华老年多器官疾病杂志(2016年7期)2016-04-28
癌症进展(2016年10期)2016-03-20
广西林业科学(2016年3期)2016-03-16
医学研究杂志(2015年5期)2015-06-10
医学研究杂志(2015年5期)2015-06-10
中国交通信息化(2015年12期)2015-06-06
