
外泌体—电离辐射诱导旁效应的另一种机制
2014-09-28 01:51蒋友芹尹晓明田文倩王敬东杨红英
辐射研究与辐射工艺学报 2014年3期
陈 纤 蒋友芹 尹晓明 田文倩 王敬东 杨红英
(苏州大学医学部放射医学与防护学院 放射医学及交叉学科研究院 苏州 215123)
电离辐射诱导旁效应指受照射细胞周围没有受到照射的细胞表现出来的生物学效应。已有大量研究证明了旁效应的存在[1-4],但介导旁效应的分子机制却仍然不清楚。至今,已提出4种介导旁效应的分子机制,包括细胞间缝隙连接、信号细胞膜上配体与旁效应细胞膜上受体之间的相互作用、信号细胞释放的细胞因子或者生长因子与其在旁效应细胞膜上受体之间的相互作用以及细胞受照后分泌的各种可溶性分子[5]。最近,有报道指出细胞受照后分泌的外泌体可能介导旁效应[6]。外泌体是一类多种细胞均可分泌的膜结构囊泡[7-8],直径30-150nm,内含蛋白质、RNA和脂类,可介导细胞间通讯[9-11]。本研究以H460人非小细胞肺癌细胞为研究对象,检测细胞受照后分泌外泌体的特性,探讨外泌体是电离辐射诱导旁效应的一种可能的介导机制。
1 材料与方法
1.1 材料
人非小细胞肺癌细胞系H460购自中国科学院上海生命科学研究院细胞库;胎牛血清购自美国Hyclone公司;RPMI1640培养基、丙酮酸钠和4',6-二脒基-2-苯基吲哚(4',6-diamidino-2-phenylindole,DAPI)购自美国 Sigma-Aldrich公司;FM4-64荧光染料购自美国Invitrogen公司;青霉素-链霉素、抗HSP90小鼠单克隆抗体、抗辣根过氧化物酶标记山羊抗小鼠 IgG(H+L)和 DiO荧光染料购自中国碧云天生物技术研究所;HEPES缓冲液购自中国Robiot公司。
1.2 方法
1.2.1 细胞培养和辐照条件
H460细胞培养于含 10%胎牛血清、2.5g·L-1葡萄糖、1 mmol·L-1丙酮酸钠、100 U·mL-1青霉素-链霉素和1 mmol·L-1HEPES的RPMI1640培养基中,置于含5% CO2的培养箱中在37 ℃培养。提取外泌体实验中,为避免血清中外泌体的干扰,采用无血清培养基。
采用美国RAD SOURCE RS2000 X-射线机照射,剂量率为1.16 Gy·min-1。
1.2.2 收集条件培养液
本研究采用条件培养液转移法研究经培养液介导的电离辐射诱导旁效应[12]。细胞接种24 h后,更换新鲜培养液,接受 X-射线照射。在受照后 0到24 h内不同时间点收集条件培养液,并通过滤膜(Ф0.22 µm)过滤以除去细胞碎片。为方便起见,文中未受照射和受照射的条件培养液分别用 SCM(Conditioned medium from sham-irradiated cells)和RCM(Conditioned medium from irradiated cells)表示。另外,采用新鲜培养液作为空白对照。
1.2.3 微核形成实验
将对数生长期的3万个H460细胞接种在带无菌小圆玻片的六孔板中,培养24 h后,分别更换为新鲜培养液、条件培养液或者含外泌体的新鲜培养液继续培养24 h;弃去培养液,PBS快速洗涤2次,细胞用V甲醇: V乙酸=3:1固定10 min并风干;细胞水化后用 DAPI (10 µg·mL-1)染色 2 min,PBS 洗涤后用抗荧光淬灭液封片。小圆玻片置于倒置荧光显微镜(DM2000, Leica, Germany)下进行细胞观察、计数。含微核的细胞为阳性细胞,计数1000个细胞中阳性细胞的个数。
1.2.4 细胞克隆形成实验
将对数生长期的H460细胞接种于60mm培养皿中,100个细胞/皿,每组3皿,第二天分别更换为新鲜培养液、条件培养液或含外泌体的新鲜培养液在 37 ℃、5% CO2条件下静置培养14 d。弃去培养基,用甲醇固定,亚甲基蓝染色,镜下计数含50个细胞以上的细胞集落。计算克隆形成率(%)和细胞克隆存活率:
克隆形成率=(克隆数/接种细胞数)×100%;
细胞克隆存活率=处理组克隆形成率/对照组克隆形成率。
1.2.5 外泌体的分离与提纯
外泌体的分离和收集采用文献[13]报道的方法。具体说来,收集细胞培养液,20mL培养液培养1500万细胞,首先用孔径为0.22 μm的过滤器过滤,除去杂质和悬浮细胞;然后进行差速离心,300g离心10 min,2,000g离心10 min,10,000g离心30 min,以除去细胞碎片;上清再经100,000g离心70 min,获得细胞分泌的外泌体;外泌体经 PBS清洗,100,000g离心70 min并去除上清后重悬于PBS中,分装,-80 ℃保存备用。所有离心步骤均在4 ℃条件下进行。采用总蛋白定量方法定量外泌体:大约900万细胞的条件培养液能提取1 μg外泌体。
1.2.6 外泌体尺寸分析
用马尔文激光粒度仪 Zetasizer Nano ZS 90(Malvern, UK)测量提取的各组外泌体的尺寸分布。
1.2.7 外泌体的透射电镜鉴定
取100 μL(约5 μg)新鲜提取的外泌体溶液,用2%戊二醛固定30 min。将外泌体滴于230目碳支持膜上,2%乙酸双氧铀染色5 min,风干,透射电镜(TEM, JEM-200 CX, Jeol. Ltd., Japan)下观察。
1.2.8 Western blotting检测外泌体标记物
提取外泌体,用细胞裂解液(0.1%曲拉通X-100,10 mmol·L-1Tris (pH =7.4),10%甘油,150 mmol·L-1氯化钠,5 mmol·L-1EDTA,1 mmol·L-1原钒酸钠,1 mmol·L-1苯甲基磺酰氟(PMSF)和 0.1%蛋白酶抑制剂)在4 ℃裂解30 min,13000 r·min-14 ℃离心10 min后,吸取蛋白上清。用Bradford蛋白定量方法测定蛋白浓度,向样品中加入 5×上样缓冲液,95 ℃条件下,5 min使蛋白质变性。然后采用SDS-PAGE凝胶电泳分离蛋白后转膜至PVDF膜,膜用含 5%脱脂奶粉的 TBST缓冲液进行非特异性封闭,加入抗 HSP90小鼠单克隆抗体 4 ℃孵育过夜,TBST洗涤3次,加入二抗辣根过氧化物酶标记山羊抗小鼠IgG(H+L)室温孵育45 min。洗膜后,加入化学发光液,用多功能激光扫描成像系统(美国Amersham公司Typhoon 9410)进行扫描和图像分析。
1.2.9 外泌体与接收细胞的相互作用
将对数生长期的 3 万个 H460细胞接种在 35mm带无菌小圆玻片的小皿内。24 h后,细胞用浓度为 15 μg·mL-1的绿色细胞膜染料DiO在室温下染色15 min,PBS洗涤两次以去除未结合染料。同时,用浓度为 10 μg·mL-1的红色细胞膜染料 FM 4-64 在37 ℃下孵育外泌体15 min后,PBS洗涤2次,洗掉未结合染料。然后将染色后的外泌体加入到染色后的细胞中于37 ℃培养30 min。细胞用4%多聚甲醛固定10 min后,用DAPI染色,加抗荧光淬灭液封片,于倒置荧光显微镜下观察、摄片。
1.2.10 细胞增殖实验
接种10,000个细胞于96孔板中,24 h后,更换新鲜培养液或含外泌体的新鲜培养液,继续培养24 h后,轻轻吸出培养液,PBS轻柔冲洗细胞,吸干后,用0.5%的结晶紫甲醇溶液室温染色15 min,吸走结晶紫甲醇溶液,加入100 μL柠檬酸钠 (pH =4.2)/50% 乙醇混合液震荡30 min溶解结晶紫;用酶标仪(BIO-TEK Power Wave XS, USA)检测样品在540nm处的吸光度值。以单纯新鲜培养液组作为对照,对其它组样品数据进行标准化处理。
1.2.11 统计学分析
本研究采用Origin 8软件进行统计和作图,采用的统计学方法为Student’s t-test,p<0.05时,认为组间差异有统计学意义。所有数据均来源于独立的至少重复3次的实验结果,文中用“平均数±标准差”表示。
2 结果
2.1 X-射线可在 H460细胞中诱导经培养液介导的旁效应
本研究采用条件培养液转移法,探索了X-射线在H460细胞中诱导经培养液介导的旁效应。如图1所示,将 5 Gy X-射线照射后 18 h的条件培养液(RCM)转移到未做任何处理的旁效应细胞,旁效应细胞的微核形成率相对于未受照射条件培养液(SCM)转移组增加了 3.5-5倍,细胞克隆存活率降低了8.3%。这些结果表明X-射线可在H460细胞中诱导由培养液介导的旁效应;并且5 Gy X-射线照射后的条件培养液和10 Gy X-射线照射后的条件培养液在旁效应细胞中产生的微核形成率之间的差异不具统计学意义,表明两个不同剂量的电离辐射诱导相似的旁效应。
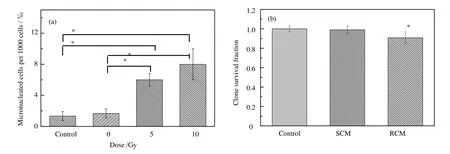
图1 X-射线在H460细胞中诱导由培养液介导的旁效应: (a) H460旁效应细胞经未受照和受照射后18 h条件培养液(SCM和RCM)培养后的微核形成率;(b) H460旁效应细胞经未受照和受照射后18 h条件培养液(SCM和RCM)培养后的克隆存活率*与新鲜培养液和未受照条件培养液对照相比, p<0.05Fig.1 X-ray irradiation induced medium-mediated bystander effects: (a) the frequency of MN formation in unirradiated bystander H460 cells cultured in SCM or RCM harvested at 18 h post-irradiation; (b) The clonogenic cell survival of unirradiated bystander cells cultured in SCM or RCM harvested at 18 h post-irradiation*p<0.05, compared with fresh medium control and SCM control
2.2 外泌体性质鉴定
已有大量文献报道外泌体是细胞间信号传递的一种方式[9-11]。本研究探索了外泌体是否在电离辐射旁效应中扮演着信号传递作用。首先分离外泌体,并鉴定其性质。如图 2a和2b所示,H460细胞不管是否受照,分泌的外泌体呈现典型的“盘杯状”特征,大小在30-150nm之间。进一步通过Western Blotting检测普遍接受的外泌体标志物HSP 90 β,确证提取物为外泌体(如图 2c)。有趣的是,不管是否受到照射,H460细胞均分泌外泌体。但是两种外泌体的粒径却完全不同,从 SCM 中提取的外泌体粒径分布在60-165nm,从RCM中提取的外泌体粒径分布在30-70nm(如图2d)。这些结果提示照射明显改变了细胞分泌外泌体的性质。未发表数据还显示细胞分泌的外泌体尺寸还依赖于细胞所受剂量和照射后时间。

图2 从SCM和RCM中提取的外泌体性质鉴定: (a)从SCM中提取的外泌体电镜图片; (b)从RCM中提取的外泌体电镜图片;(c) Western Blotting检测外泌体标志物Hsp 90 β; (d)从SCM和RCM中提取外泌体的粒径分布Fig.2 Characterization of exosomes isolated from SCM and RCM: (a) TEM images of exosomes isolated from SCM;(b) TEM images of exosomes isolated from RCM; (c) Western blot analysis of protein marker (HSP 90β) of exosomes;(d) Size distribution of exosomes isolated from SCM and RCM
2.3 外泌体与H460接收细胞之间的相互作用
为了探索外泌体是介导电离辐射旁效应的一种可能方式,将从SCM和RCM中提取的外泌体加入到H460接收细胞中孵育30 min。在此之前,将外泌体用红色膜荧光染料染色,而接收细胞用绿色膜荧光染料染色。结果如图3a和3b所示,从SCM和RCM中提取的外泌体均能迅速进入H460接收细胞中,而且可能是通过膜融合的方式[14](图中橙色代表红色和绿色的共定位)。这提示外泌体可以作为未受照射或受照射H460细胞向接收细胞传递信号物质的一种载体。
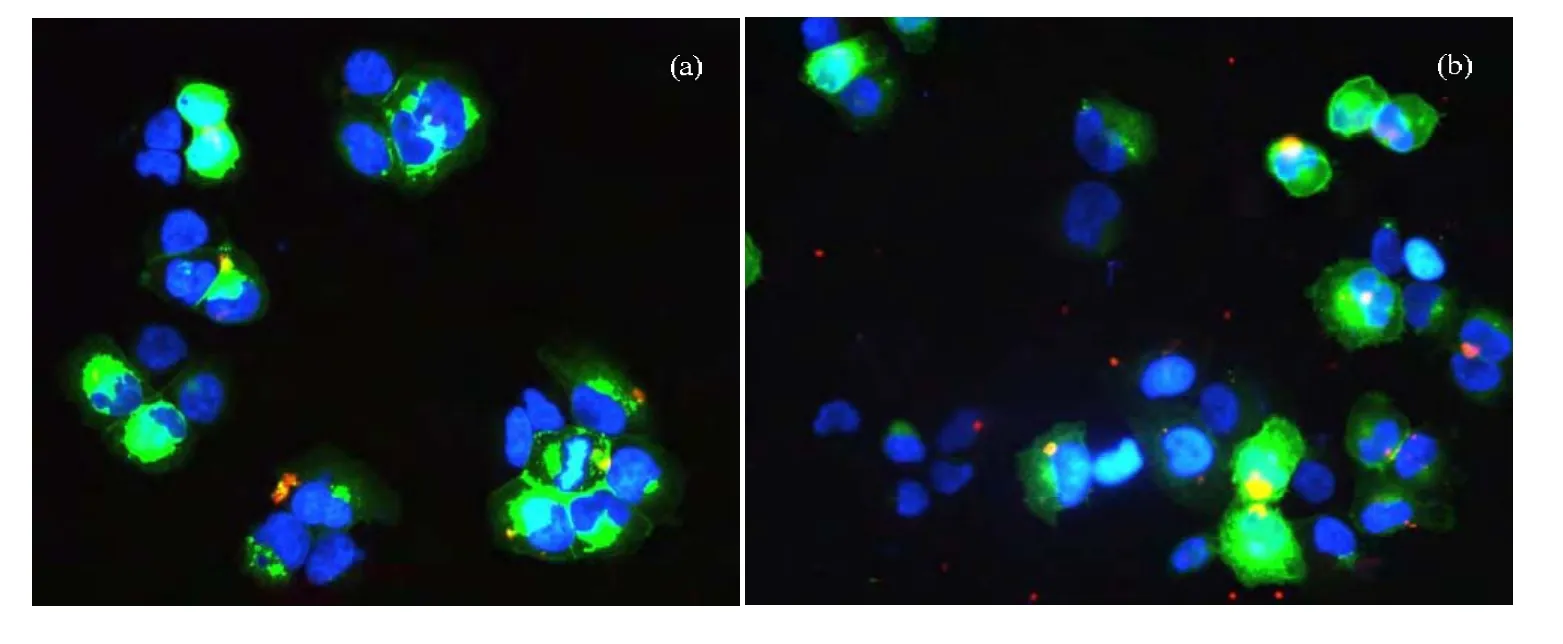
图3 从SCM(a)和RCM(b)中提取的外泌体进入接收细胞红色:FM 4-64外泌体染色;绿色:DiO细胞膜染色;蓝色:DAPI细胞核染色Fig.3 Internalization of exosomes isolated from SCM (a) and RCM (b) into the recipient cells:exosomes were labeled with FM 4-64 (red); cell membranes were labeled with DiO (green); nuclei were stained with DAPI (blue)
2.4 从条件培养液提取的外泌体可诱导接收细胞产生生物学效应
为了确证外泌体是电离辐射在H460细胞中诱导旁效应的一种介导方式,将从SCM和RCM中提取的外泌体直接加入到H460接收细胞中。与对照相比,加入从 SCM 中提取的外泌体未改变接收细胞的微核形成率,而从RCM中提取的外泌体却使接收细胞的微核形成率增加了两倍(见图 4a)。表明从受照射细胞培养液提取的外泌体与未照射条件下提取的外泌体性质不同,前者可以在接收细胞中诱导生物学变化,提示外泌体是电离辐射诱导旁效应的一种可能机制。并且当先用RNA酶在37 ℃与外泌体孵育90 min后,再将外泌体加入接收细胞,之前观察到的从RCM中提取的外泌体使接收细胞的微核形成率增加的生物学效应消失了,表明外泌体中 RNA在其诱导微核形成增加中发挥着重要作用。然而,未发表数据显示,不管是否受照,H460细胞分泌的外泌体均不改变接收细胞的克隆存活率,说明受照后条件培养液介导的克隆存活率降低可能并不是由外泌体介导的。另外,与新鲜培养液对照组相比,不管是否受照,H460细胞分泌的外泌体均能促进接收细胞的增殖(大约11%,见图4b),这表明,不管是否受照,H460细胞分泌的外泌体均可能介导细胞间信号传递,促进肿瘤细胞生长。
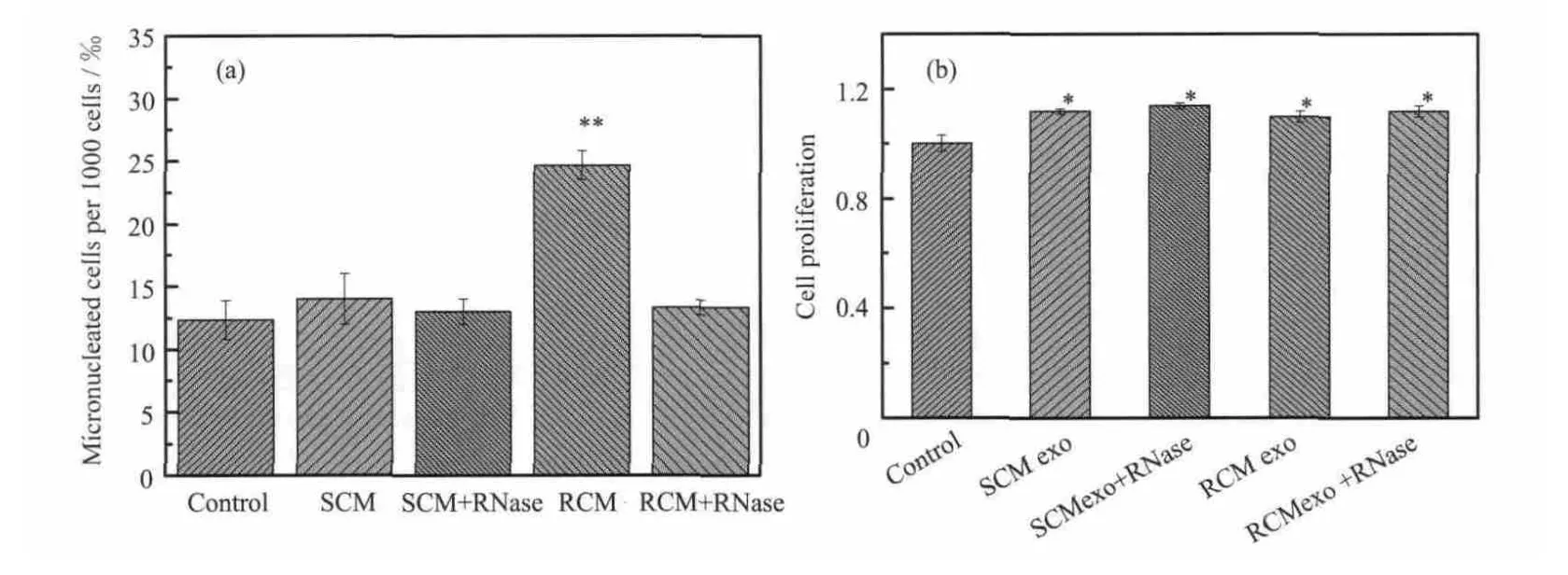
图4 从SCM和RCM中提取纯化的外泌体对接收细胞微核形成的影响(a)和从SCM和RCM中提取纯化的外泌体对接收细胞增殖的影响(b): *与新鲜培养液对照相比, p<0.05; **与新鲜培养液对照相比, p<0.01Fig.4 The frequency of MN formation in the recipient cells cultured in medium with exosomes isolated from SCM and RCM (a)and the cell proliferation of the recipient cells cultured in medium with exosomes isolated from SCM and RCM(b)* p<0.05, compared with fresh medium control; ** p<0.01, compared with fresh medium control
3 讨论
本研究证实了电离辐射可在H460非小细胞肺癌细胞中诱导条件培养液介导的旁效应(见图1),旁效应细胞的微核形成增加,细胞克隆存活率降低,DNA损伤增加(未发表数据)。这与文献报道的X-射线照射H460细胞导致细胞因子释放,从而产生表现为细胞存活和凋亡改变的旁效应相一致[15]。尽管文献[15]提示电离辐射在H460细胞中诱导的旁效应可由肿瘤坏死因子相关的凋亡诱导配体 TRAIL(TRAIL与促凋亡蛋白 PAR-4的核转位有关)介导,本研究提供证据证实H460细胞受照后分泌的外泌体是诱导旁效应的一种可能机制。
多细胞体系中,细胞间通讯对于各种类型细胞的正常组织和功能必不可少。除了被广为接受的细胞间通讯机制,例如细胞间隙连接、释放的生长因子、化学因子和小分子包括多肽、离子、核酸和脂质等,越来越多的证据表明外泌体-这种特殊的纳米膜结构囊泡,可以介导细胞间通讯[16]。外泌体是一类直径在30-150nm,可由多种细胞,包括各种肿瘤细胞、免疫细胞和正常细胞释放的囊泡[7-8]。外泌体包含丰富的脂质、蛋白质和多种核酸,特别是miRNAs和通常以蛋白质复合体形式存在并在细胞间通讯中发挥关键作用的RNAs[10-11,16-17]。本研究发现,从未受照射和受照射细胞的条件培养液中均可提取到外泌体(图2),表明H460非小细胞肺癌细胞分泌外泌体既是组成型又是诱导型。这和之前的文献报道有所不同,该报道显示未受照H460细胞并不分泌可检测的外泌体[18]。一个可能的原因是两项研究中所用细胞培养条件和外泌体提取方法的不同。然而本研究发现H460细胞受照前后,其释放的外泌体的尺寸分布不同,受照H460细胞释放的外泌体比未受照细胞分泌的外泌体尺寸小(图2)。细胞受照前后分泌的外泌体大小不同提示受照后外泌体成分可能发生了变化。未发表数据表明,受照射细胞分泌的外泌体尺寸分布与细胞所受剂量和照射后培养时间相关,提示受照射细胞在不同剂量和不同培养时间情况下可能分泌不同的信号分子。
从未受照射和受照射细胞的条件培养液提取的外泌体可以迅速进入接收细胞,可能是通过膜融合的方式[14],见图3中橙色,由不同膜染料的红色和绿色叠加形成,表明 H460细胞释放的外泌体可以直接将信号分子传递进接收细胞。有趣的是,从受照射细胞条件培养液提取的外泌体显著增加接收细胞的微核形成(图4a)表明细胞受照后分泌的外泌体内容物与未受照时不同。然而,不管受照射与否,H460细胞分泌的外泌体对接收细胞均有促增殖作用,两者之间没有统计学差异(图 4b)。这与文献报道胶质瘤细胞释放外泌体促进胶质瘤细胞增殖类似[9]。未发表数据显示不管是否受照,H460细胞分泌的外泌体均不改变接收细胞的克隆存活率。这些结果提示,不管是否受照射,H460 细胞分泌的外泌体均含有促细胞生长因子,但这些细胞因子对H460细胞的克隆存活率没有影响;并且,细胞受照后释放的外泌体中含有诱导微核形成的因子,这可能是照射后转移条件培养液导致旁效应细胞微核增加的机制之一。受照细胞释放的外泌体能增加接收细胞的微核形成,但不改变接收细胞的克隆存活率,这些结果提示导致电离辐射诱导旁效应的不同表现的机制可能不一样。这与之前 Yang等[2]的报道相似,他们证实了活性氧淬灭剂可降低 X-射线在AG01522细胞中诱导的经培养液介导的旁效应DNA损伤,但却对旁效应细胞杀伤没有影响,从而提示电离辐射诱导的不同旁效应可能存在不同的机制。
并且, RNA可能是受照细胞分泌的外泌体中重要的效应分子,因为用RNase处理外泌体可消除其增加接收细胞微核形成的效应(图 4a)。这与之前文献[6]报道外泌体中 RNA 介导电离辐射旁效应相一致。但是,在外泌体中加入RNase对接收细胞的增殖无影响(图 4b)。这进一步提示,外泌体中的促微核形成因子和促细胞增殖因子并不相同。
总之,本研究证实了X-射线照射H460细胞可诱导经培养液介导的旁效应,表现为微核增加和克隆存活率下降。另一方面,H460细胞可分泌组成型和诱导型外泌体,受照细胞分泌的外泌体与未受照射细胞分泌的外泌体在尺寸分布上有很大不同,两者都可通过膜融合的方式迅速进入接收细胞,从而促进接收细胞的增殖;并且,受照细胞分泌的外泌体可导致接收细胞的微核形成率增加,而 RNA酶可抑制这种增加;但是受照细胞分泌的外泌体不影响接收细胞的克隆存活率。这些结果表明外泌体是介导电离辐射旁效应的一种可能机制,其中 RNA发挥着重要作用。但是,未受照射和受照射细胞分泌的外泌体的蛋白质和 RNA等成分构成和引起接收细胞生物学效应的信号分子需要进一步研究。
1 Zhou H, Suzuki M, Randers-Pehrsong, et al. Radiation risk to low fluences of alpha particles may be greater than we thought [J]. Proceedings of the National Academy of Sciences USA, 2001, 98: 14410-14415.
2 Yang H, Asaad N, Held K D. Medium-mediated intercellular communication is involved in bystander responses of X-ray-irradiated normal human fibroblasts[J]. Oncogene, 2005, 24: 2096-2103.
3 Shao C, Folkard M, Michael B D, et al. Targeted cytoplasmic irradiation induces bystander responses [J].Proceedings of the National Academy of Sciences USA,2004, 101: 13495-13500.
4 Mancuso M, Pasquali E, Leonardi S, et al. Oncogenic bystander radiation effects in Patched heterozygous mouse cerebellum [J]. Proceedings of the National Academy of Sciences USA, 2008, 105: 12445-12450.
5 Hamada N, Matsumoto H, Hara T, et al. Intercellular and intracellular signaling pathways mediating ionizing radiation-induced bystander effects [J]. Radiation Research, 2007, 48: 87-95.
6 Al-Mayah A H, Irons S L, Pink R C, et al. Possible role of exosomes containing RNA in mediating nontargeted effect of ionizing radiation [J]. Radiation Research, 2012,177: 539-545.
7 Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses [J]. Nature Reviews Immunology, 2009, 9: 581-593.
8 Wolfers J, Lozier A, Raposog, et al. Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming [J]. Nature Medicine, 2001, 7:297-303.
9 Skog J, Würdinger T, Van Rijn S, et al. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers [J].Nature Cell Biology, 2008, 10: 1470-1476.
10 Record M, Carayon K, Poirot M, et al. Exosomes as new vesicular lipid transporters involved in cell-cell communication and various pathophysiologies. [J].Biochimica et Biophysica Acta, 2013, 1841: 108-120.
11 Valadi H, Ekström K, Bossios A, et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells [J]. Nature Cell Biology, 2007, 9: 654-659.
12 Lyng F M, Seymour C B, Mothersill C. Initiation of apoptosis in cells exposed to medium from the progeny of irradiated cells: a possible mechanism for bystander-induced genomic instability [J]. Radiation Research, 2002, 157: 365-370.
13 Théry C, Clayton A, Amigorena S, et al. Isolation and characterization of exosomes from cell culture supernatants and biological fluids [J]. Current Protocols in Cell Biology, 2006, 3.22.1-3.22.29.
14 Tian Y, Li S, Song J, et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy [J]. Biomaterials,2014, 35(7): 2383-2390.
15 Shareef M M, Cui N, Burikhanov R, et al. Role of tumor necrosis factor-alpha and TRAIL in high-dose radiation-induced bystander signaling in lung adenocarcinoma [J]. Cancer Research, 2007, 67:11811-11820.
16 Mathivanan S, Ji H, Simpson R J. Exosomes: extracellular organelles important in intercellular communication [J].Journal of Proteomics, 2010, 73: 1907-1920.
17 Mathivanan S, Fahner C J, Reidg E, et al. ExoCarta 2012:database of exosomal proteins, RNA and lipids [J].Nucleic Acids Research, 2012, 40: D1241-D1244.
18 Yu X, Harris S L, Levine A J. The regulation of exosome secretion: a novel function of the p53 protein [J]. Cancer Research, 2006, 66: 4795-4801.
猜你喜欢
基础医学与临床(2022年1期)2022-11-25
昆明医科大学学报(2022年2期)2022-03-29
中学生物学(2021年8期)2021-11-02
昆明医科大学学报(2021年5期)2021-07-22
老年医学研究(2021年6期)2021-03-09
绿色科技(2019年18期)2019-11-22
科学家(2017年19期)2017-10-27
现代家庭(2016年12期)2016-12-07
科学启蒙(2015年8期)2015-08-07
安徽农学通报(2014年16期)2014-09-18
