
江西武夷山-黄岗山西北坡森林繁殖鸟类多样性调查
2014-09-19 10:59程松林毛夷仙袁荣斌
生态学报 2014年23期
程松林,毛夷仙,袁荣斌
(江西武夷山国家级自然保护区, 铅山 334500)
鸟类,特别是繁殖鸟类与环境的关系十分密切。根据鸟类既依赖于环境又反映环境的变化,并对环境变化高度敏感的特性,鸟类被作为生物区系分析的重要对象之一,常常被用于监测环境变化或作为生物多样性的指示物,而且在环境评价和保护规划中都是重要的评价指标[1- 3]。在诸多影响鸟类的环境因子中,植被直接影响到鸟类的分布和多度,是鸟类生存和栖息必须的第一因子[4]。成功的繁殖关系到整个动物种群的存在与灭绝[5],动物的繁殖必须在食物条件、身体状况及社群结构等协调一致的情况下才能进行[6],否则在不良的环境中要保持正常的繁殖成功率,亲体必须增大自身的繁殖代价[7]。良好的巢址是保障鸟类繁殖成功的先决条件[8],在长期的自然选择过程中,鸟类倾向于选择自身存活代价低且繁殖成效高的地方营巢[9- 10],以最大限度地降低天敌捕食、同类干扰和其他不利因素的影响[10- 12]。
武夷山是我国生物多样性的热点地区和亚热带中山森林保存完好的交汇地带,属于生物多样性优先保护区中需要最优先保护的区域[13],其主峰黄岗山海拔2 160.8 m,为秦岭-黄河以南,大巴山-武陵山-乌蒙山-哀牢山以东陆域的最高峰[14]。武夷山的木本植物物种丰富度在我国东部主要山地中仅次于海南尖峰岭和广西猫儿山[15]。自19世纪40年代英国人福琼(R.Fortune)首次到武夷山进行植物标本采集开始[16],武夷山就一直受到生物学界的高度关注。1938—1948年和1963年,郑作新等人先后在福建邵武、崇安、建阳等县域的武夷山区进行了鸟类调查,记录到鸟类47科234种(3亚种)[17];2004—2011年程松林等先后在江西武夷山国家级自然保护区记录鸟种17目51科151属269种[18- 19]。
但有关武夷山的鸟类多样性与植被、景观多样性之间的关系研究尚未见报道。希望通过本研究掌握研究区森林繁殖鸟多样性状况,特别是研究区森林繁殖鸟的海拔梯度分布、植被不同演替阶段和类型森林中繁殖鸟的差异等,为进一步开展研究区森林生态效益评估,提高武夷山区的生物多样性保护管理内涵提供基础数据。
1 研究区域和数据处理
1.1 研究区域
研究区属于江西武夷山国家级自然保护区范围,地处武夷山脉北段、主峰黄岗山的西北坡,地理坐标为27°48′11″—28°00′35″ N,117°39′30″—117°55′47″ E,面积16007 hm2。
研究区属于亚热带中部季风湿润区,年均气温14.2℃,年均降水量2 583 mm,年均蒸发量778 mm,气候特征表现为:春季回暖迟多雾天、夏季凉爽多雨、秋季短暂变温快、冬季寒冷常结冰;地貌为中低山山地地貌,海拔高度介于300—2 160.8 m、平均海拔约1 200 m,山体平均坡度>40°。
植物区系特征表现为:原生状态保存完整、种类丰富,热带性质科占明显优势,中亚热带向北亚热带过渡性明显,与华南植物区系联系密切,珍稀濒危物种众多,区系起源古老,新老特有种并存等;植被垂直带谱典型发育,植被型多样、代表性强,乔灌层物种丰富、郁闭度高,草本层和层间物种较少。记录有高等植物2 800余种,脊椎动物515种。研究区人口密度<5人/ km2,海拔900 m以上无村庄。
1.2 数据来源与处理
1.2.1 数据来源
一是近几年研究区文献数据[18- 19],二是于2011—2012年进行的跟踪观察补充数据。
(1)样线设计
2011—2012年补充调查样线与研究区文献记录调查样线相同。样线设计综合考虑研究区海拔高度、代表性植被类型、人类活动干扰情况和便于长期监测等因素,设计了篁村(海拔430 m,坐标27°57′07″ N、117°46′26″ E)—桐木关(海拔1 161 m,坐标27°49′00″ N、117°42′46″ E)—黄岗山(海拔2 160.8 m,坐标27°51′35″ N、117°47′01″ E)主样线1条,长约39 km,该样线贯穿了研究区所有生境类型、代表性植被类型、不同强度人类干扰活动区,其中篁村至桐木关段为山区公路(长19.5 km、宽4—6 m),日均车流量<30辆,属于保护区实验区范围内,桐木关至黄岗山段为废弃30多年的军用公路(长19.6 km、宽约4 m),日均车流量<10辆,属于保护区的缓冲区和核心区内。辅助样线从东北至西南依次为:擂鼓岭(海拔300 m,坐标28°00′26″ N、117°52′40″ E)—山羊坑(海拔631 m,坐标27°58′30″ N、117°54′02″ E),长约8.5 km;分水关(海拔743 m,坐标27°56′10″ N、117°50′21″ E)—七仙山(海拔1 379 m,坐标27°57′32″ N、117°50′29″ E),长约10.5 km;篁村(海拔430 m,坐标27°57′07″ N、117°46′26″ E)—桐梓关(海拔1 137 m,坐标27°55′17″ N、117°47′54″ E),长约8.0 km;西坑(海拔680 m,坐标27°53′32″ N、117°44′59″ E)—杨家州(海拔1 062 m,坐标27°53′07″ N、117°43′09″ E),长约7.0 km;大岩(海拔670 m,坐标27°52′22″ N、117°39′56″ E)—大龙坑(海拔1 374 m,坐标27°53′58″ N、117°41′01″ E),长约7.0 km。辅助样线为废弃的生产便道或林间小路,路宽1.5—4.0 m,各条辅助样线之间距离不小于5 km,并涵盖了研究区所有生境类型、代表性植被类型、不同强度人类干扰活动区和保护区功能区。
(2)调查设备
双筒望远镜(施华洛世奇EL 10X42WB),数码照相机(佳能40D,镜头2.8×200 mm),数码摄像机(索尼3CCD,变焦12×),GPS(麦哲伦Triton500),以及其它记录工具等。
(3)补充调查时间与频率
调查于2011年5月8日—7月23日和2012年5月5日—8月16日的繁殖季,选择晴天的06:00—08:00和16:00—18:00进行,2个繁殖季内主样线累计调查37次,辅助样线累计调查21次(每条辅助样线、每个繁殖季调查1—3次)。
(4)调查方法
每次调查由2—4人组成调查组。主样线调查分为篁村至桐木关段和桐木关至黄岗山段进行调查用机动车代步,速度控制在15 km /h左右,采用目视和双筒望远镜观察,有条件的进行拍照或摄像,发现目标及时记录物种名、时间、地点、生境、海拔、坐标等相关信息,两段样线相邻调查时间间隔不超过3d。辅助样线每个繁殖季,采用步行调查,速度为2—3 km /h,其他调查方法、记录因子与主样线调查相同。
1.2.2 数据处理
将研究区鸟类名录中的冬候鸟和游禽、涉禽等主要依赖湿地生境的鸟种,以及调查中仅记录1—2只次、且没有见到巢或雏鸟等确凿繁殖证据的鸟种剔除,形成研究区森林繁殖鸟名录。
多样性分析评估采用G-F指数法[20],相似性系数采用公式[21]:
Sc=[2C/(A+B)]×100%
鉴于隼形目(FALCONIFORMES)和雨燕目(APODIFORMES)鸟种活动范围较大、佛法僧目(CORACIIFORMES)鸟种喜好选择有较多水面的栖息地、形目(PICIFORMES)鸟种喜好选择有较大树木的栖息地[22]、夜鹰目(CAPRIMULGIFORMES)和戴胜目(UPUPIFORMES)鸟种数过于单一等原因可能致使可比性差,在分析研究区所有鸟种的G-F指数和相似性系数基础上,再选择活动范围相对较小[23]、物种数量多的雀形目(PASSERIFORMES)鸟种单独进行G-F指数和相似性系数分析比较。
2 结果
2.1 繁殖鸟多样性资源
研究区动物地理区划上属于东洋界华中区东部丘陵平原亚区(ⅥA)[24]。在研究区共记录到森林繁殖鸟43科115属188种,分别占研究区鸟类科、属、种数的84.31%、76.16%、69.89%,繁殖鸟中有留鸟114种、夏候鸟74种。研究区森林繁殖鸟种数占东部丘陵平原亚区繁殖鸟种数的71.76%(188/262)[25](表1)。
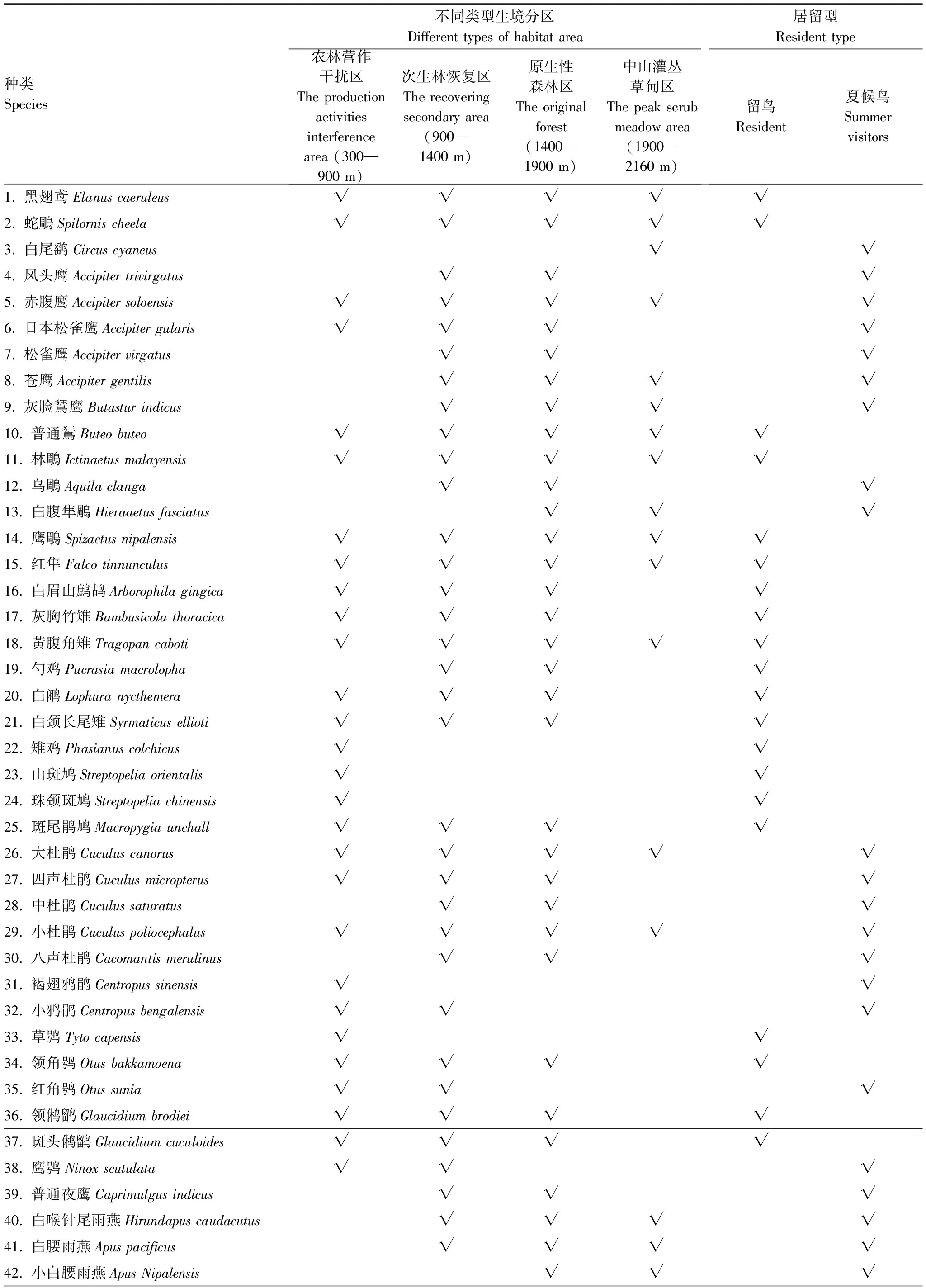
表1 研究区森林繁殖鸟名录
2.2 不同生境森林繁殖鸟分布
2.2.1农林营作干扰区(海拔300—900 m)
该区域主要植被为自然恢复20a左右的常绿阔叶林、灌丛,有经营活动的毛竹(Phyllostachysheterocycla)林和少量的人工杉木(Cunninghamialanceolata)林、茶(Cameriasinensis)地,以及农田、旱地、道路、村庄和较为开阔、水流较为平缓的河流等。生境表现出景观特异化明显、人为干扰较大的特征。见于本区的鸟种有119种,其中有30种仅见于该区。
2.2.2次生林恢复区(海拔900—1 400 m)
该区域主要植被为自然恢复20—40a之间的常绿阔叶林、常绿落叶阔叶混交林、针叶阔叶混交林和灌丛,林龄为正在向顶级群落演替的中龄林、近成熟林,一些陡峭的裸岩山地残存有成熟林,在海拔900—1200 m之间有少量的毛竹林和茶地。生境体现为植被类型和林龄多样化。见于本区的鸟种有138种,其中有5种仅见于该区。
2.2.3原生性森林区(海拔1400—1900 m)
该区域森林基本为原生状态的过熟林或成熟林,植被主要为常绿落叶阔叶混交林、针叶阔叶混交林和针叶林,在海拔1800—1900 m区间有一以落叶阔叶树为主的苔藓矮曲林带,部分沟谷在海拔1700 m左右仍可见到常绿阔叶林群落。该区以植被原生状态保存完好、乔木-灌木层次丰富、少有人为干扰为显著特征。见于本区的鸟种有127种,其中有9种仅见于该区。
2.2.4中山灌丛草甸区(海拔1900—2160 m)
在林线附近或背风处的草甸中,散生有明显矮化的台湾松(Pinustaiwanensis)、云锦杜鹃(Rhododendronfortunei)和匍匐状的波叶红果树(Stranvaesiadavidianavar.undulate),山顶处则基本由野青茅(Deyeuxiaarundinacea)、芒(Miscanthussinensis)为主成份的草甸,期间常见孤石和裸岩。以风力大、气温低、紫外线辐射强,地表水相对缺乏为显著特征。见于本区的鸟种有51种,其中有6种仅见于该区。
2.2.5 生境适应广泛性和狭窄性鸟种
在研究区4类生境均能见到的鸟种有31种、占总数的16.5%,见于3类生境的有47种、占25.0%,见于2类生境的60种、占31.9%,仅见于1类生境中的鸟种50种、占26.6%。对生境适应较为广泛(见于3—4类生境)的鸟种78种,生境适应较为狭窄(见于1—2类生境)的鸟种有110种。
2.3 G-F指数与相似性系数
研究区鸟类G-F指数分析结果。所有鸟种DG-F=0.823,雀形目DG-F=0.740(表2)。
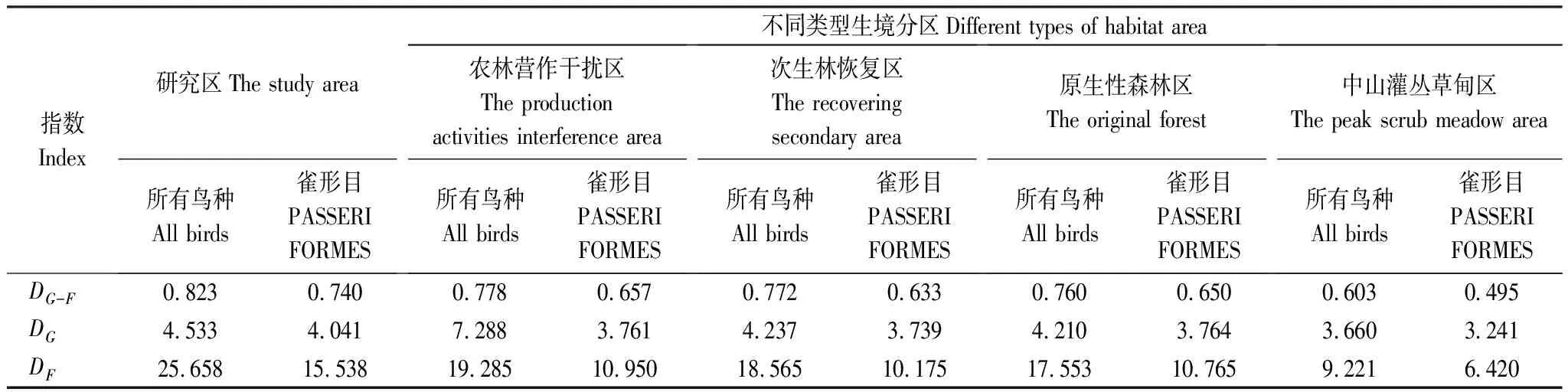
表2 研究区繁殖鸟G-F指数
相似性分析结果。研究区繁殖鸟的科、属、种的相似性,均以次生林恢复区和原生性森林区的相似性系数最高、农林营作干扰区和次生林恢复区次之、原生性森林区与中山灌丛草甸区再次之,农林营作干扰区和中山灌丛草甸区的相似性系数最低(表3)。对雀形目所有鸟种和雀形目夏候鸟、留鸟分别进行相似性分析,结果与所有繁殖鸟种的结果一致。
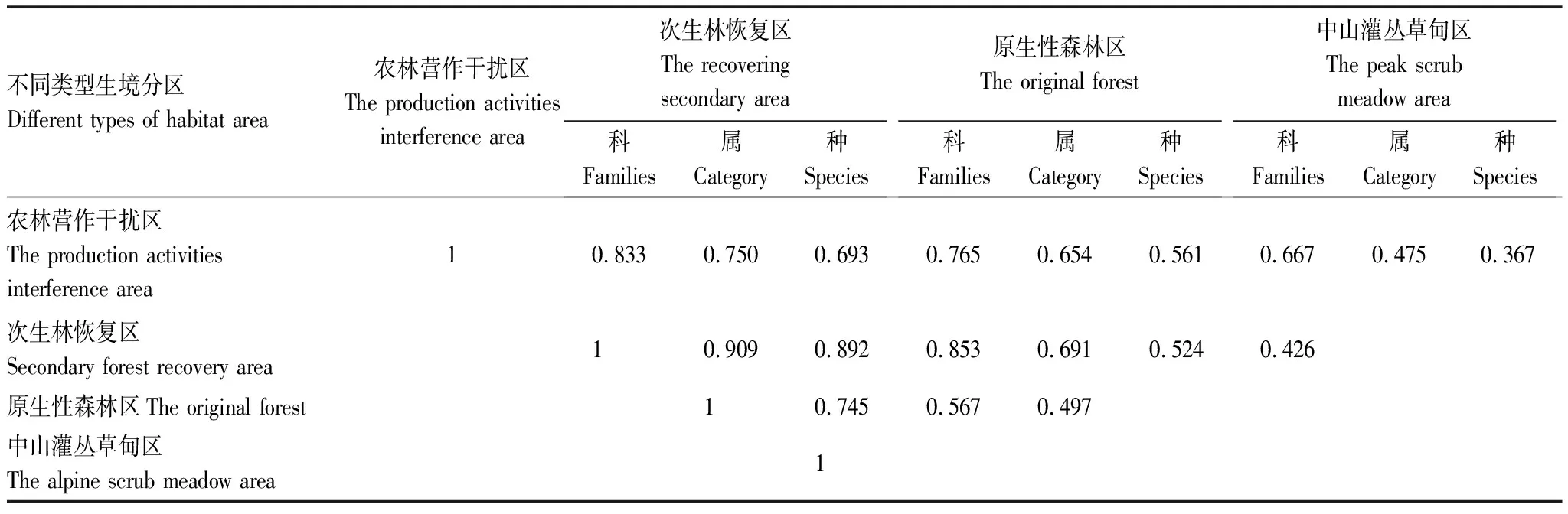
表3 研究区所有鸟种相似性系数
3 讨论
3.1 森林群落特征对森林繁殖鸟多样性的影响
由表1可见,研究区188种森林繁殖鸟中雀形目占68.6%,处于绝对优势地位,其中物种数在10种及以上的科有鸫科(Turdidae)13种、鹟科(Muscicapidae)10种、画眉科(Timaliidae) 21种、莺科(Sylviidae)22种,这4个科的鸟种数占研究区鸟种数的35.1%、雀形目鸟种数的51.2%。在4类生境中,中山灌丛草甸区鸟种数明显偏少,可能与植被过于单一和低矮有关。其它3类生境中,鸟种数以居中间位置的次生林恢复区最多,并且在科、属、种3个分类阶元上均与原生性森林区的相似性系数最高,说明演替到接近顶级群落的植被,其生态功能对于森林繁殖鸟来说,已经成为区域多数鸟种的适宜繁殖栖息地;同时,除中山灌丛草甸区由于生境条件的差异各项指标数值均最小,其余3类生境(海拔300—1 900 m),无论是总的鸟种数、雀形目鸟种数,还是4个优势科鸟种数,均支持中间膨胀效应(Mid-domain effect)。与广西猫儿山鸟类种数和G-F指数随着海拔的升高逐渐减少有所区别[26],这些差别是否与调查方法不同,或者是其他因子影响有待进一步研究。
3.2 景观和微环境多样性对繁殖鸟多样性的影响
农林营作干扰区是研究区景观多样性和微环境结构最复杂的区域,自然景观、半自然景观、人工景观,以及陡峭边缘(Abrupt dege)、过度边缘(Intermediate dege),精细边缘(Subtle edge) 相互交错。其中相对较多的湿地能够满足佛法僧目、戴胜目等湿地型鸟类对生境的特殊要求;村庄、房屋则为家燕(Hirundorustica)、金腰燕(Cecropisdaurica)、山麻雀(Passerrutilans)等鸟种提供了合适的巢址。在农林营作区,鸟种绝对数虽略少于次生林恢复区和原生性森林区,但G-F指数各项指标在研究区4类生境中均位列第一(表2),说明多样的景观和复杂的微环境结构能够为更多的、对生境有不同需求的繁殖鸟提供合适的繁殖场。
3.3 植被不同演替阶段和森林内部层次对森林繁殖鸟多样性的影响
根据样方调查,研究区的次生林恢复区和原生性森林区的植被基本为近成熟林、成熟林或过熟林,相对于农林营作干扰区,乔木层树木高(8—35 m)、胸径大,可分为2—3个亚层,灌层物种丰富、高差大(1—7 m),常与小型乔木相互交错、疏密相间,使其单位面积可利用森林内部空间必然更大,微环境更为多样。而复杂多样的森林内部结构和较大的森林空间,有利于森林鸟类在栖息基层、取食基层及取食方式上的分离和集团内部的各个种间对取食高度的划分[27],同时植被类型决定着巢稳固因子、巢隐蔽条件和周边树因子[12],植被结构特征影响着巢卵被捕食率[28- 29],有的甚至会影响繁殖率,比如在有较高大的乔木和较低的灌木盖度森林内,杂色山雀(Parusvarius)的繁殖成功率就较高[30]。从G-F指数结果看,原生性森林区对雀形目鸟种多样性表现出一定的积极意义;从相似性系数结果看,经过30a左右自然演替的次生林恢复区,生态功能与顶级群落已比较接近。该结果与Andrade在哥伦比亚亚马逊的研究结果:早期次生林与中期次生林、原始林相似性小,中期次生林与原始林相似性大基本相似[31]。
3.4 研究区鸡形目鸟类的生态位分异
在研究区,鸡形目鸟类生态位分异最为典型。研究区共有鸡形目鸟类7种,其中体型较大的雉鸡仅见于海拔700 m以下的森林-农耕地带,勺鸡(Pucrasiamacrolopha)主要分布于海拔1000—1900 m范围,白颈长尾雉(Syrmaticusellioti)则主要见于二者之间的海拔650—1500 m范围;体型较小的灰胸竹鸡(Bambusicolathoracica)和白眉山鹧鸪(Arborophilagingica)的主要栖息地大致以海拔1200—1300 m为分水岭,灰胸竹鸡主要在低海拔地带、白眉山鹧鸪在高海拔地带种群密度则明显高于低海拔地带;黄腹角雉(Tragopancaboti)和白鹇(Lophuranycthemera)是研究区栖息地适应范围较广的物种,但黄腹角雉主要分布于海拔690—1900 m林内,偏好选择在灌木层为木本植物、且相对较为稀疏的林地,并有较多的树上觅食或歇息行为,白鹇主要分布于海拔1 600 m以下的森林,偏好选择灌层盖度更大和有较多小型竹类(Bambusoideae)的栖息地,没有发现树上觅食、活动现象。
致谢:江西省科学院林剑声先生,江西武夷山国家级自然保护区的邹思成等同志参加了野外调查,特此致谢。
:
[1] Padoa-Schioppa E, Baietto M, Massa R, Bottoni L. Bird communities as bioindicators: the focal species concept in agricultural landscapes. Ecological Indicators, 2006, 6(1): 83- 93.
[2] Furness R W, Greenwood J J D, Jarvis P J. Can birds be used to monitor the environment? // Furness R W, Greenwood J J D, eds. Birds as Monitors of Environmental Change. London: Chapman and Hall, 1993: 1- 41.
[3] Zhao H F, Lei F M. Birds as Monitors of Environmental Change. Chinese Journal of Zoology, 2002, 37(6): 74- 78.
[4] Michael M L. Bird populations as indicators of environmental change. Current Ornithology, 1986, 14: 430- 448.
[5] Shang C Y. Behavioral Ecology. Beijing: Peking University Press, 1998.
[6] Bronson F H. Mammalian reproduction: an ecological perspective. Biology of Reproduction, 1985, 32(1): 1- 26.
[7] Sikes R S, Ylönen H. Considerations of optimal litter size in mammals. Oikos, 1998, 83(3): 452- 465.
[8] Kokko H, Harris M P, Wanless S. Competition for breeding sites and site dependent population regulation in a highly colonial seabird, the common guillemotUriaaalge. Journal of Animal Ecology, 2004, 73(2): 367- 376.
[9] Zheng G M. Ornithology. Beijing: Beijing Normal University Press, 1995.
[10] Valkama J, Korpimaeki E, Tolonen P. Habitat utilization, diet and reproductive success in the Kestrel in a temporally and spatially heterogeneous environment. Ornis Fennica, 1995, 7(2): 49- 61.
[11] Martin T E. Avian life history evolution in relation to nest sites, nest predation, and food. Ecological Monographs, 1995, 65(1): 101- 127.
[12] Shi L M, Liu N F, Ding W, Zhao Q S, Fang F J, Bao X K. Nest-site selection and breeding success of the rufous-tailed shrike. Chinese Journal of Zoology, 2012, 47(6): 7- 13.
[13] Li D Q, Song Y L, Ouyang Z Y. Research on the National Forestry Nature Reserve System Plan. Beijing: China Land Press, 2003.
[14] Wang X P, Wang Z H, Fang J Y. Mountain ranges and peaks in China. Biodiversity Science, 2004, 12(1): 206- 212.
[15] Fang J Y. Exploring altitudinal patterns of plant diversity of China′s mountains. Biodiversity Science, 2004, 12(1): 1- 4.
[16] Luo G H. Biological exploration in Wuyi mountain by modern westerners. The Chinese Journal for the History of Science and Technology, 2002, 23(1): 31- 37.
[17] Zheng Z X, Jiang Z H, Tang R Y. Preliminary studies on the avifauna of MT. Wuyi Area. Wuyi Science Journal, 1981, 1: 153- 167.
[18] Cheng S L, Lin J S. A survey on avian diversity in Wuyishan national nature reserve, Jiangxi. Chinese Journal of Zoology, 2011, 46(5): 66- 78.
[19] Cheng S L, Liu J N, Zhang Y Y. The Birds of Wuyishan National Nature Reserve. Beijing: Science Press, 2001.
[20] Jiang Z G, Ji L Q. Avian-mammalian species diversity in nine representative sites in China. Biodiversity Science, 1999, 7(3): 220- 225.
[21] Zhang Y L. Coefficient of similarity-an important parameter in floristic geography. Geographical Research, 1998, 17(4): 429- 434.
[22] Luo W Z, Song Y J. Preying behavior of spotted woodpecher. Chinese Journal of Ecology, 1992, 11(5): 25- 27.
[23] Terborgh J, Robinson WST, Parker TA III, Munn C A, Pierpont N. Structure and organization of an Amazonian forest bird community. Ecological Monographs, 1990, 60(2): 213- 238.
[24] Zhang R Z. China Animal Geographic. Beijing: Science Press, 1999.
[25] Wang K F, Zhang J R, Lei F M. Geographical distribution pattern and its temporal and spatial variation of birds breeding in zoogeographical Subregion of China. Acta Zootaxonomica Sinica, 2010, 35(1): 145- 157.
[26] Su T P, Wang S N, Jiang A W. Species composition and vertical distribution pattern of the birds in Mao′ershan region, Northeastern Guangxi. Chinese Journal of Zoology, 2012, 47(6): 54- 65.
[27] Liu H, Han L X. Birds foraging guilds in Xujiaba evergreen broad-leaved forest of Ailao mountain. Zoological Research, 2008, 29(5): 561- 568.
[28] Clark R G, Shutler D. Avian habitat selection: Pattern from process in nest-site use by ducks. Ecology, 1999, 80(1): 272- 287.
[29] Martin T E, Roper J J. Nest predation and nest site selection of a western population of the hermit Thrush. Condor, 1988, 90(1): 51- 57.
[30] Li L, Wan D M, Liu H, Yin J X, Li Q J, Huo Y P. Nest site selection and reproductive success of Parus varius in man-made nest boxes. Acta Ecologica Sinica, 2011, 31(24): 7492- 7499.
[31] Andrade G I, Rubio-Torgley H. Sustainable use of the tropical rain forest: Evidence from the avifauna in a shifting-cultivation habitat mosaic in the Colombian Amazon. Conservation Biology, 1994, 8(2): 545- 554.
参考文献:
[3] 赵洪峰, 雷富民. 鸟类用于环境监测的意义及研究进展. 动物学杂志, 2002, 37(6): 74- 78.
[5] 尚昌玉. 行为生态学. 北京: 北京大学出版社, 1998.
[9] 郑光美. 鸟类学. 北京: 北京师范大学出版社, 1995.
[12] 施丽敏, 刘迺发, 丁未, 赵青山, 房峰杰, 包新康. 荒漠伯劳巢址选择和繁殖成功. 动物学杂志, 2012, 47(6): 7- 13.
[13] 李迪强, 宋延龄, 欧阳志云. 全国林业系统自然保护区体系规划研究. 北京: 中国大地出版社, 2003.
[14] 王襄平, 王志恒, 方精云. 中国的主要山脉和山峰. 生物多样性, 2004, 12(1): 206- 212.
[15] 方精云. 探索中国山地植物多样性的分布规律. 生物多样性, 2004, 12(1): 1- 4.
[16] 罗桂环. 近代西方人对武夷山的生物学考察. 中国科技史料, 2002, 23(1): 31- 37.
[17] 郑作新, 江智华, 唐瑞于. 福建武夷山地区鸟类区系初探. 武夷科学, 1981, (1): 153- 167.
[18] 程松林, 林剑声. 江西武夷山国家级自然保护区鸟类多样性调查. 动物学杂志, 2011, 46(5): 66- 78.
[19] 程松林, 刘江南, 张雁云. 武夷山自然保护区鸟类. 北京: 科学出版社, 2011.
[20] 蒋志刚, 纪力强. 鸟兽物种多样性测度的G-F指数方法. 生物多样性, 1999, 7(3): 220- 225.
[21] 张镱锂. 植物区系地理研究中的重要参数: 相似性系数. 地理研究, 1998, 17(4): 429- 434.
[22] 罗维桢, 宋榆钧. 大斑啄木鸟取食行为的研究. 生态学杂志, 1992, 11(5): 25- 27.
[24] 张荣祖. 中国动物地理. 北京: 科学出版社, 1999.
[25] 王开锋, 张继荣, 雷富民. 中国动物地理亚区繁殖鸟类地理分布格局与时空变化. 动物分类学报, 2010, 35(1): 145- 157.
[26] 粟通萍, 王绍能, 蒋爱伍. 广西猫儿山地区鸟类组成及垂直分布格局. 动物学杂志, 2012, 47(6): 54- 65.
[27] 刘菡, 韩联宪. 云南哀牢山徐家坝常绿阔叶林的鸟类取食集团. 动物学研究, 2008, 29(5): 561- 568.
[30] 李乐, 万冬梅, 刘鹤, 殷江霞, 李其久, 霍雅鹏. 人工巢箱条件下杂色山雀的巢位选择及其对繁殖成功率的影响. 生态学报, 2011, 31(24): 7492- 7499.
猜你喜欢
区域治理(2023年6期)2023-03-02
中国农学通报(2022年32期)2022-11-28
湖南大学学报(自然科学版)(2022年8期)2022-09-02
大众摄影(2021年3期)2021-03-22
——以北长山岛为例
海洋开发与管理(2020年9期)2020-10-13
现代农业科技(2020年10期)2020-06-04
数理化解题研究(2018年34期)2018-12-27
数理化解题研究(2018年16期)2018-07-12
岷峨诗稿(2014年1期)2014-11-15
中国期刊年鉴(2013年0期)2013-02-16
