
红地球与夏黑无核成熟期光合日变化规律研究
2014-09-10 06:28聂松青石雪晖田淑芬刘昆玉杨国顺钟晓红倪建军
河北林业科技 2014年1期
聂松青 ,石雪晖 ,田淑芬 ,刘昆玉 ,杨国顺 ,钟晓红 ,倪建军 ,徐 丰 ,白 描
(1.湖南农业大学园艺园林学院,湖南 长沙 410128;2.湖南省葡萄工程技术研究中心,湖南 长沙 410128;3.天津市农科院葡萄研究中心,天津 300112)
葡萄叶片的光合特性和光合能力直接影响到葡萄浆果的品质和产量的形成,光合作用生成的有机化合物不仅为生物提供能量,也是生物用以建造自身躯体的原料。随着便携式光合测定系统的日益普及和对光合效率与作物产量关系认识的深入,人们越来越广泛地把光合特性和光合效率用作选育和鉴定优良资源的重要指标。目前,红地球葡萄是国内栽培面积最大的葡萄品种之一,夏黑无核葡萄因其早熟、粒大、无核等优良品性得以迅速推广。在南方高温高湿、避雨栽培环境下,研究红地球和夏黑无核葡萄的光合特性,有利于采取合理的栽培措施和管理模式,对提高设施葡萄叶片光能的利用效率,提高果实的产量和品质具有重要意义[1]。
1 材料与方法
1.1 试验材料
供试葡萄品种为湖南农业大学葡萄教学科研基地7a生红地球和夏黑无核葡萄,南北行向,避雨栽培,架式为小平棚。选择生长发育良好、生长势基本一致的红地球葡萄和夏黑无核葡萄品种各3株,每株选择同侧节位无病虫害、长势中庸的结果枝上第6节位功能叶作为测定叶片,每片叶重复测定3次。
1.2 试验方法
在夏黑无核成熟期(7月上旬)和红地球成熟期(8月下旬)的大晴天,使用美国LI-COR公司生产LI-6400XT便携式光合仪,采用自然光源,于8:00~18:00每2h测定1次,每次重复3次。记录的主要参数有净光合速率(Pn,μmol·m-2·s-1)、光照有效辐射(PARi,μmol·m-2·s-1)、蒸腾速率(Ts,mmol H2Om-2·s-1)、气孔导度(Gs,cm·s-1)、胞间CO2浓度(Ci,μmol·mol-1)、相对湿度(RH,%)、叶片温度(Tleaf,℃)。
1.3 数据处理
采用Excel 2007处理数据及绘制图表。
2 结果与分析
2.1 两个葡萄品种成熟期环境因子的日变化
2.1.1 光合有效辐射(PAR)日变化
光合有效辐射对葡萄的生长发育影响很大,在一定光照强度范围内,随着光照强度的增加,光合效率也相应增加。在测定期间,从试验数据发现(图1),太阳光照强度日变化表现为先增加后减少的“单峰型”曲线,即清晨光强较弱,在上午 8:00~10:00增长迅速,午间 13:00左右最强,之后又逐渐减弱,回到上午8:00的光合有效辐射水平。
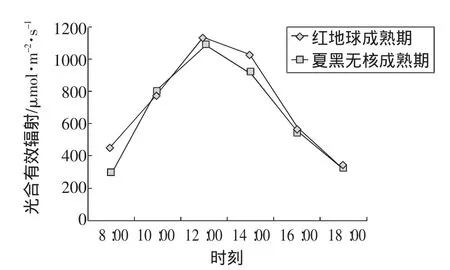
图1 光合有效辐射日变化
2.1.2 叶片温度(Tleaf)日变化
南方夏季炎热,白天高温天气持续时间长。红地球和夏黑无核葡萄的叶片气温日变化表现为先增加后减少的“单峰型”曲线变化趋势,即早上8:00开始就有32℃左右的高温,随着时间的推移,温度增加迅速,午间14:00左右最高,达到最高温41.9℃(红地球)和41.4℃(夏黑无核),午后又逐渐降低(图2)。下午18:00,光强逐渐减弱,但平棚内温度并未大幅降低,保持在37℃左右。

图2 叶片温度日变化
2.1.3 相对湿度(RH)日变化
叶片的湿度受气温和光照影响明显。如图3所示,红地球和夏黑无核葡萄叶片相对湿度日变化表现为先下降后上升的趋势。一天当中,设施内空气相对湿度变化范围为30%~50%。上午8:00的空气湿度较大,随着时间的递增,空气湿度逐渐变小,中午14:00左右空气湿度最小,14:00过后空气湿度有所增加。主要原因是上午温度相对较低,水分蒸发速度较慢;中午温度高,水分蒸发快;午后光合作用和蒸腾作用又开始增强,空气湿度又有所增加,而后随着温度的降低和光照的减弱逐渐减小。空气相对湿度的变化趋势与温度变化趋势相反,可能与大气温度、光强等环境因子直接相关。
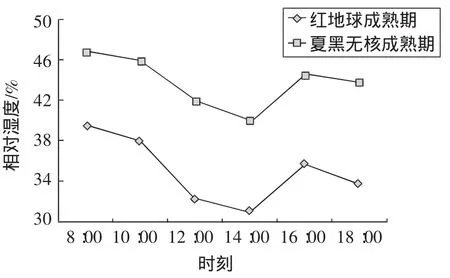
图3 相对湿度日变化
2.2 两个葡萄品种成熟期光合生理因子的日变化
2.2.1 胞间CO2浓度(Ci)日变化
随着时间的推移,红地球葡萄胞间CO2浓度呈现出“W型”曲线变化,夏黑无核葡萄呈现“V型”曲线变化。清晨,叶片气孔敞开,大量吸收大气CO2而净光合速率较低,固定CO2能力较低,使细胞间隙CO2浓度最高;随着温度、有效光辐射的升高,较多的CO2被固定,引起细胞间隙CO2浓度下降。随着气温升高和光照增强,10:00左右当净光合速率最高时,红地球葡萄胞间CO2浓度下降到第1个低值,夏黑无核细胞间隙CO2浓度达到最低值。光合“午休”时红地球葡萄因气孔关闭阻止细胞内外CO2浓度交换,胞间CO2浓度相对较低,而此时夏黑无核葡萄浓度没有降低反而上升是因为有非气孔限制因素。午后,虽然叶片气孔再次敞开,红地球葡萄胞间CO2浓度迅速回升。此后随着时间推移,光合作用减弱,胞间CO2浓度又有所增加,如图4。
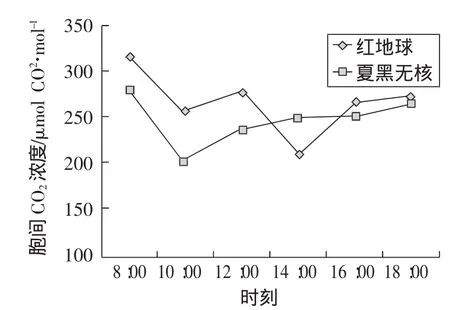
图4 胞间CO2浓度日变化
2.2.2 气孔导度(Gs)日变化
在光照强度较低的情况下,气孔导度(Gs)随光照强度的升高而增大,随CO2浓度的升高而减小。在不同光照强度下,气孔导度随CO2浓度变化的幅度有所不同,但总的来说都是随着二氧化碳浓度的增加而减小。由图5可知,红地球和夏黑无核葡萄叶片气孔导度日变化规律是双峰曲线。从早上8:00开始,随光强的增加和气温的上升,气孔逐渐打开,蒸腾作用加强,第一个峰值都出现在上午10:00左右,随着时间的推移逐渐下降。午间的高温强光使叶温上升,气孔阻力增大,气孔导度迅速减小,在下午14:00左右出现低谷,之后气温回落,在一定适宜温度范围内,气孔再度张开,气孔导度又经过短期的增长到16:00左右出现第2个峰值,之后随着温度的下降气孔导度逐渐减小呈下降趋势。
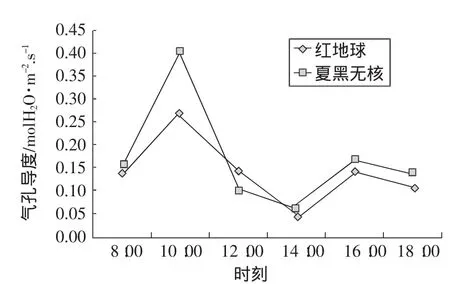
图5 气孔导度的日变化
2.2.3 蒸腾速率(Tr)日变化
蒸腾速率取决于叶片内外的蒸气压差和扩散途径阻力的大小,光照、温度、大气相对湿度是影响蒸腾速率的主要因素。气孔导度和蒸腾速率的变化趋势基本一致,说明蒸腾速率在很大程度上决定于气孔的活动状态。上午8:00左右,叶片气孔导度较小,蒸腾作用较弱。随着光照增强,气温逐渐增高,叶片水汽压差也随之增大,蒸腾速率不断升高,10:00左右出现第1次峰值;之后随着时间推移,光强增大,气温升高,叶片气孔逐渐缩小直至关闭,蒸腾速率随之降低;午后“午休”过后,蒸腾作用随光合作用增强而增强,16:00左右达到第2次峰值,此后蒸腾速率随时间推移递减(图 6)。

图6 蒸腾速率日变化
2.2.4 净光合速率(Pn)日变化
晴朗天气条件下,红地球和夏黑无核葡萄叶片成熟期Pn日变化趋势双峰曲线。在光饱和点以下,随时间的推移和光合有效辐射的增强,光合速率急剧增大,第1次峰值出现在上午10:00左右,达到一天中最大净光合速率(即红地球Pnmax 为 12.8μmol/m2·s和夏黑无核 Pnmax为11.8μmol/m2·s)。而后由于光照过强,引起气温升高,空气湿度下降,导致叶片水热不协调,进而导致气孔收缩,气孔阻力增大,气孔导度迅速减小,CO2吸收量减小,光合速率下降。在高温、低CO2浓度的午间,产生光抑制现象,致使光合作用减弱,14:00左右叶片出现明显的光合“午休”现象。午后随着气温降低,在一定温度范围内,光合速率又缓慢回升,第2次峰值出现在下午16:00左右。第2峰值相对第1峰值较低。之后,随着温度的下降光合速率逐渐减小。

图7 红地球与夏黑无核葡萄净光合速率日变化
2.3 关于光合日变化的影响因子及相关性
本研究发现,在果实成熟期,无论是红地球,还是夏黑无核葡萄,环境因子光合有效辐射和叶片温度均呈先升后降的“单峰”型曲线变化趋势,而相对湿度呈相反的变化趋势,即先下降后上升再下降。两种葡萄在成熟期温度变化受光照强度的影响,两者变化呈正相关;而湿度与光照变化趋势相反,呈负相关。光照强度是影响温度和湿度的主要因子,同时三者的相互作用又对叶片的光合特性产生影响。
两个葡萄品种叶片净光合速率(Pn)与主要生理生态因子之间存在密切关系,净光合速率(Pn)分别与叶片蒸腾速率(Tr)和气孔导度(Gs)呈极显著正相关。蒸腾速率、气孔导度与光合速率日变化趋势相似且升降幅度相对较大,且一天中的最大值为第1峰值。
3 讨论与结论
植物光合作用的日变化类型主要有4种:正规曲线型、平坦型、变动型和中午降低型[2]。在果树上主要有3种表现:单峰型曲线[3-4]、双峰型曲线[5-6]和极少数的三峰型曲线[7]。强光、高温、低湿、蒸腾速率高、气孔关闭等生理生态因子可能是午间叶片出现光合“午休”现象的原因。Pn中午降低的因素主要有2个:气孔因素和非气孔因素。气孔因素是由于中午光照、温度和湿度等环境因子变化引起植物气孔部分关闭,CO2进入叶片受阻而使光合下降,而非气孔因素是由于叶肉细胞自身羧化酶活性的下降而引起Pn降低。根据Farquhar和Sharkey[8]的观点,只有当 Pn和 Ci二者同时减小,且Ls(1-Ci/Ca)降低时,才可以认为Pn下降主要是由气孔引起的,否则Pn下降要归因于叶肉细胞羧化能力的降低。通过对成熟期红地球葡萄的Ci、Ls与Pn之间的关系进行分析,Ls在进入光合“午休”阶段呈下降的趋势,此时Ci、Pn均呈下降的变化。表明红地球叶片光合“午休”导致Pn下降的主要原因均是气孔因素。同时通过对成熟期夏黑无核葡萄的Ci、Ls与Pn之间的关系进行分析,可推断出,Ls在进入光合“午休”阶段呈下降的趋势,而此时Ci呈上升、Pn下降的变化,表明夏黑无核叶片光合“午休”导致Pn下降的主要原因均是非气孔因素。
葡萄的生长发育与光合作用息息相关,葡萄光合作用合成的有机物不仅为浆果发育提供能量来源,也是其建造自身躯体的原料。光合生理生态因子之间都是相互联系相互作用的,众多因子间的相互作用对光合作用的影响至今都不清楚。今后,有关光合作用与环境因子的关系还需进一步探索研究。
[1]陈冲,杨国顺,石雪晖,等.避雨栽培条件下红宝石无核葡萄光合特性初探[J].中外葡萄与葡萄酒,2011,(3):21-24.
[2]吕忠恕.果树生理[M].上海:上海科学技术出版社,1982:55.
[3]彭少兵,郭军战.不同树莓和黑莓品种的光合特性研究[J].西北农林科技大学学报:自然科学版,2007,35(3):116-120.
[4]张志华.核桃光合特性的研究[J].园艺学报,1993,20(4):319-323.
[5]李延菊,李宪利.3个扁桃品种的光合特性[J].林业科学,2006,42(11):23-28.
[6]郭春芳,孙云,陈常颂,等.茶树品种光合与水分利用特性比较及聚类分析[J].作物学报,2008,34(10):1797-1804.
[7]王文江.大磨盘柿树光合特性的研究[J].园艺学报,1993,20(2):105-110.
[8]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Ann Rev PlantPhysiology,1982,33:317-345.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
