
山东半岛南部三疣梭子蟹放流前后渔业生物群落结构初步分析
2014-08-03 09:29邱盛尧张金浩耿宝龙
烟台大学学报(自然科学与工程版) 2014年3期
李 增,邱盛尧,张金浩,耿宝龙
(烟台大学海洋学院,山东 烟台 264005)
随着我国近海海域污染的加重和捕捞强度的增强,渔业资源量锐减,渔业生物群落结构也随之发生了很大的改变.为了改善现有海域的渔业种群结构,提高渔业资源量,各地相继开展了品种繁多的增殖放流活动.然而,国外相关资料表明,增殖放流在提高放流物种资源量的同时,还必须保证放流水域生态系统的稳定性[1].为此,国内许多学者对山东半岛南部海区生物群落结构进行了研究,金显仕等[2]对山南水域春季游泳动物群落结构变化进行了研究分析,李涛等[3]则对山南近海渔业资源群落结构变化进行了相关分析.在此基础上,本文通过对2012年三疣梭子蟹增殖放流前后放流点渔业群落的动态研究,初步探讨三疣梭子蟹增殖放流前后渔业生物群落结构的变化,旨在为三疣梭子蟹放流对山东南部近海的生态环境损益评价提供科学依据.
1 材料与方法
1.1 调查方法
对三疣梭子蟹增殖放流的影响进行了2个航次的调查,分别是三疣梭子蟹放流前的本底调查(5月20日—5月30日)和放流后的资源结构变化调查(6月15日—6月25日).两个航次的调查取样均在无风晴天的白天进行(一般为9∶00—14∶00左右),调查船为功率14.7 kW的渔船,每个放流海湾设置了1~8个调查站位(图1),调查网具为单船底拖网.网具规格为网口1 400目,网目尺寸56 mm,网口周长78.4 m,囊网网目20 mm,每站拖网30 min,拖速1.5节.现场对捕获的渔获物进行分类和测定,记录每种渔获物的数量、重量以及相关生物学数据.
1.2 数据分析
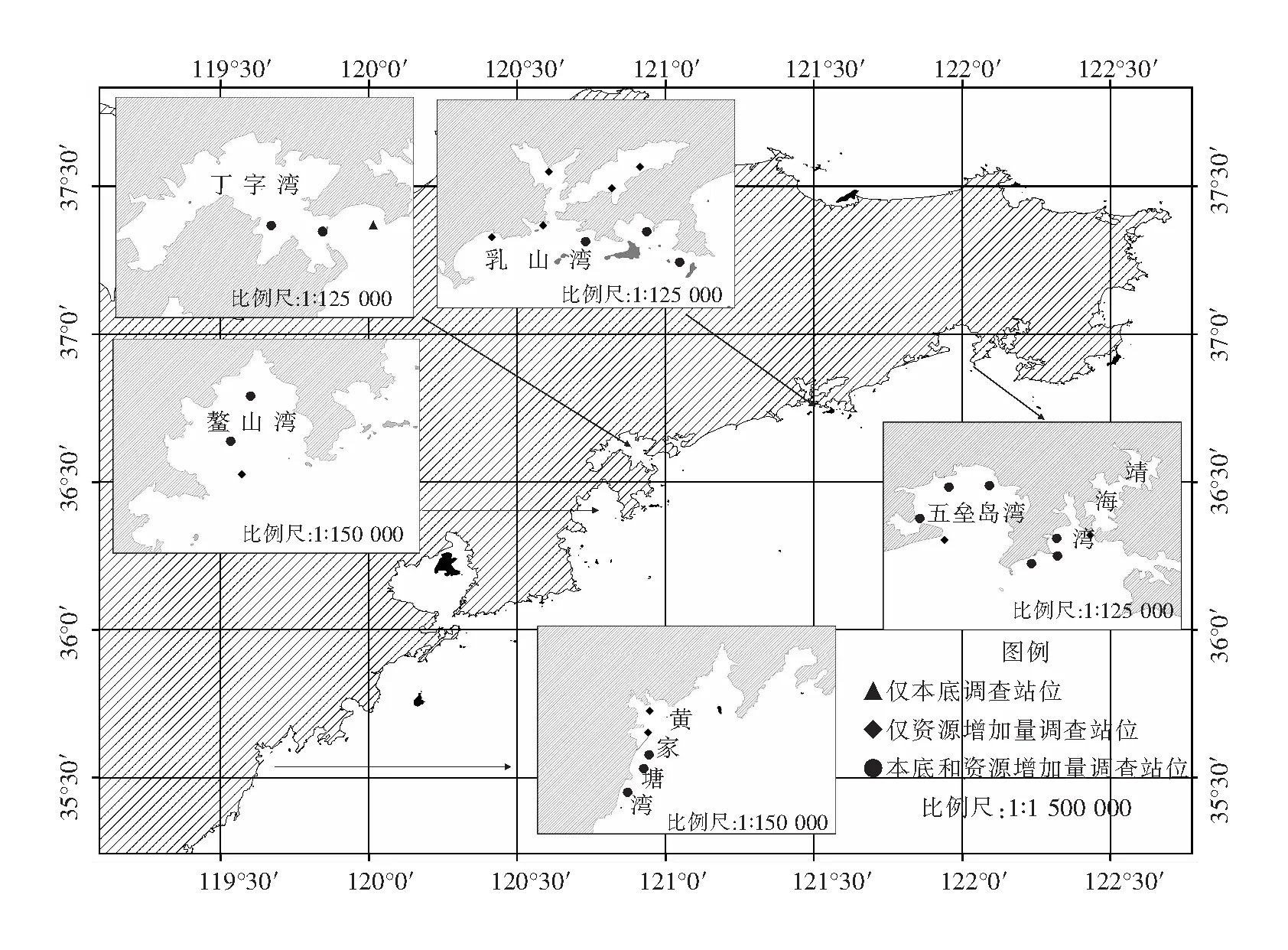
图1 调查站位分布图
1.2.1 渔获量 渔获数量和重量根据捕捞系数进行修正,其中,中下层鱼类和虾类捕捞系数为0.5,底栖鱼类和蟹类为0.8[4],换算为单位/小时的标准质量和标准数量.
1.2.2 优势种 利用Pinkas相对重要性指数(Index of Relative Importance,简称IRI),确定种类在群落中的重要性[5-6].
IRI=(N+W)F,
其中:N为渔获物的数量比例,W为质量比例,F为各海湾出现频率.当IRI>1 000时,该种类为优势种;100 1.2.3 生物多样性 利用Margalef的种类丰富度指数R、Shannon-Wiener多样性指数H′、Pielou均匀度指数J′来分析渔业资源群落生态多样性[3,5]. 种类丰富度指数R: R=(S-1)/lnM, 多样性指数H′: H′=-ΣPilnPi, 均匀度指数J′: J′=H′/lnS. 其中:S为种类数,M为总渔获量(g),Pi为i种渔获量占总渔获量的比例. 1.2.4 群落结构 利用大型多元统计分析软件PRIMER 6.0中的等级聚类CLUSTER和非度量多维标度(Multi-Dimensional Scaling,简称MDS)来展现群落结构格局的分布;SIMPER分析法分析渔获物对样本组内和组间相异性的平均贡献率,本文定义贡献率≥5%的为主要特征种[7-9].根据丰度生物量比较曲线(Abundance Biomass Comparison Curve,简称ABC曲线),对渔业资源群落受影响的程度进行分析[10]. 2个航次调查共鉴定渔业生物种类43种,其中鱼类22种,甲壳类19种,头足类2种,其中放流前的种类共计40种,其中鱼类21种,甲壳类17种,头足类2种;放流后的种类共计43种,其中鱼类22种,甲壳类19种,头足类2中(表1).群落的种类组成比例相对稳定,以鱼类和甲壳类为主体,头足类较少.放流前后渔业资源种类变化不大,其中鱼类以短吻红舌鳎(Cynoglossusjoyneri)为主,花鲈(Lateolabraxjaponicus)、星康吉鳗(Congermyriaster)、黄姑鱼(Nibeaalbiflora)、白姑鱼(Argyrosomusargentatus)等也为各海湾常见种类,许氏平鲉(Sebastesschlegeli)、鮻(Chelonhaematocheilus)、石鲽(Platichthysbicoloratus)、鲬 (Platycephalusindicus)等有零星分布;甲壳类以口虾蛄(Oratosquillaoratoria)、鹰爪虾(Trachypenaeuscurvirostris)和日本蟳(Charybdisjaponica)为主,中国对虾(Penaeuschinensis)为后期新增种类,但数量相对较少;头足类渔获种类没有变化. 放流前后优势种组合见表2. 表1 放流前后渔获种类统计 表2 放流海湾优势种和重要种组成 注:N为渔获物数量比例,W为渔获物质量比例,F为各海湾出现频率. 由表2可见,放流前,以戴氏赤虾(Metapenaeopsisdalei)、口虾蛄(Oratosquillaoratoria)、鹰爪虾(Trachypenaeuscurvirostris)、日本关公蟹(Dorippejaponica)以及日本蟳(Charybdisjaponica)为群落优势种,合计占渔获总数量的56.7%,占总重量的49.0%;以红线黎明蟹(Matutaplanipes)、短吻红舌鳎、日本鼓虾(Alpheusjaponicus)、红狼牙鰕虎鱼(Odontamblyopusrubicundus)、脊尾白虾(Exopalaemoncarinicauda)、日本枪乌贼(Loligojaponica)、矛尾刺鰕虎鱼(Acanthogobiushasta)、星康吉鳗、三疣梭子蟹(Portunustrituberctdatus)、周氏新对虾(Metapenaeusjoyneri)、黄姑鱼、花鲈、葛氏长臂虾(Palaemongravieri)、小黄鱼(Pseuclosciaenapolyactis)等为重要种,合计占渔获总数量的33.9%,占总重量的39.8%. 放流后,优势种为口虾蛄、鹰爪虾和日本蟳,合计占渔获总数量的49.6%,占渔获总重量的37.6%,重要种略有变化,为日本关公蟹、日本枪乌贼、短吻红舌鳎、红线黎明蟹、日本鼓虾、日本枪乌贼、白姑鱼、红狼牙鰕虎鱼、纹缟鰕虎鱼(Tridentigerbarbatus)、星康吉鳗、周氏新对虾、栉鰕虎鱼(Ctenotrypauchenchinensis)、葛氏长臂虾、花鲈、叫姑鱼(Johniusgrypotus)和戴氏赤虾,合计占渔获总数量的40.1%,占渔获总重量的49.5%(表2). 生物多样性分析见表3. 表3 放流海湾生物多样性特征分析 注:S为渔获种类,R为丰度指数,H′为多样性指数,J′为均匀度指数. 放流前,各湾渔获种类(S)为11~27,平均为17种,丰富度指数(R)变化范围1.25~3.02,平均值为1.91;多样性指数(H′)变化范围为1.74~2.76,平均值为2.18;均匀度指数(J′)变化范围为0.64~0.86,平均值为0.78.丰富度指数和多样性指数最高值均出现在乳山湾,均匀度指数最高值则出现在黄家塘湾;丰富度指数最低值出现在五垒岛湾,多样性指数及均匀度指数最低值则出现在鳌山湾. 放流后,各湾渔获种类(S)为11~27,平均为17种,丰富度指数(R)变化范围1.26~3.11,平均值为1.89;多样性指数(H′)变化范围为2.01~2.88,平均值为2.32;均匀度指数(J′)变化范围为0.74~0.89,平均值为0.83.丰富度指数及多样性指数最高值出现在乳山湾,均匀度指数最高值出现在鳌山湾;丰富度指数最低值出现在五垒岛湾,多样性指数最低值出现在靖海湾,均匀度指数最低值则出现在黄家塘湾(表3). 由聚类分析以及MDS分析(图2)可以看出,在胁迫系数为0.01的情况下,调查前后的渔业生物群落大致可以划分为2个组群,组群Ⅰ为丁字湾和靖海湾,组群Ⅱ为五垒岛湾、乳山湾、黄家塘湾以及丁字湾.放流前,组群Ⅰ组内相似性为19.99%,组群Ⅱ组内相似性为59.16%,放流后,2个组群组内相似性发生改变,组群Ⅰ组内相似性升高(组群Ⅰ排列由疏变密),为36.38%,组群Ⅱ组内相似性降低(组群Ⅱ排列由密变疏),为32.10%,但总体分布趋势不变,仍可划分为2个组群. ANOSIM分析表明,放流前组群间的种类组成差异性极显著(R=0.698,P<0.001),放流后组群间的种类组成差异性仍然显著(R=0.883,P<0.05);其中,组群Ⅰ放流前后的种类组成差异不显著(R=0.073,P>0.05),而组群Ⅱ放流前后的种类组成差异显著(R=0.133,P<0.05). SIMPER分析结果显示,放流前对组群Ⅰ贡献度较高的种类为日本蟳、日本关公蟹、口虾蛄、红线黎明蟹以及短吻红舌鳎等,相似性系数贡献率总和为93.42%;对组群Ⅱ相似性贡献度较高的种类为日本蟳、鹰爪虾、戴氏赤虾、口虾蛄、日本关公蟹、红线黎明蟹以及短吻红舌鳎等,相似性系数贡献率总和为85.51%(表4),组群Ⅰ与组群Ⅱ差异性系数为83.35%,主要差异种为戴氏赤虾、鹰爪虾、周氏新对虾、脊尾白虾以及口虾蛄等(表5). 放流后,对组群Ⅰ贡献度较高的种类为周氏新对虾、红线黎明蟹、日本关公蟹以及日本蟳等,相似性系数贡献率总和为91.00%,对组群Ⅱ贡献度较高的种类为鹰爪虾、红狼牙鰕虎鱼、日本蟳、口虾蛄以及红线黎明蟹等,相似性系数贡献率总和为81.22%(表4);组群Ⅰ与组群Ⅱ差异性系数为81.51%,主要差异种为鹰爪虾、周氏新对虾、日本鼓虾以及日本对虾等(表5). 图2 三疣梭子蟹放流前后各海湾渔业资源聚类分析及MDS分析 表4放流前后对组群平均相似性贡献率≥5%的主要特征种 Tab.4 The main characteristic species whose average similarity distribution rate≥5% % 种类放流前组群Ⅰ组群Ⅱ放流后组群Ⅰ组群Ⅱ 日本蟳Charybdis japonica31.648.316.02 日本关公蟹Dorippe japonica27.128.3816.42 口虾蛄Oratosquilla oratoria17.3313.195.75 短吻红舌鳎 Cynoglossus joyneri9.045.31 红线黎明蟹Matuta planipes8.295.8723.545.71 鹰爪虾rachypenaeus curvirostris29.0155.66 戴氏赤虾Metapenaeopsis dalei23.75 红狼牙鰕虎鱼Odontamblyopus rubicundus8.08 周氏新对虾Metapenaeus joyneri42.73 相似性贡献率合计93.4285.5191.0081.22 表5放流前后对组群间差异性贡献率≥5%的主要特征种 Tab.5 The main characteristic species whose average differ-ence distribution rate≥5% % 种类差异性系数放流前放流后 戴氏赤虾Metapenaeopsis dalei17.54 鹰爪虾rachypenaeus curvirostris17.0034.97 周氏新对虾Metapenaeus joyneri12.269.33 脊尾白虾Exopalaemon carinicauda7.68 口虾蛄Oratosquilla oratoria6.21 日本鼓虾Alpheus japonicus5.29 日本囊对虾Marsupenaeus japonicas5.02 放流前,丰度曲线高于生物量曲线,表明该时段各湾渔业种群收到严重干扰;放流后,生物量曲线逐步升上,与丰度曲线重叠翻转,表明该时段渔业种群结构发生变化,生物量开始大幅增加,种群干扰程度由重度干扰变为中度干扰,种群结构质量在一定程度上发生了改善(图3). 图3 渔业群落ABC曲线分析 山东半岛南部海域是山东近海渔业生物重要的产卵、索饵和越冬区,其生物群落结构的变化直接影响着山东近海渔业经济的发展和生态系统的稳定.由于人为因素的干扰和理化环境的变化,使得我国近海渔业生物群落结构变化较大,严重影响了海区的生态稳定[4].调查期间,各湾的生物群落主要呈现以下基本特征:一是甲壳类生物组成比重提高,压缩鱼类比重.三疣梭子蟹放流后,甲壳类组成比重由42.50%提高到44.19%,鱼类由52.50%减少到51.16%.渔业经济种类以甲壳类为主体,鱼类种类较少.在IRI≥500的种类中,甲壳类有11种,其中经济价值较高的鹰爪虾、口虾蛄以及日本蟳的IRI>1 000,为群落优势种;经济鱼类中,除短吻红舌鳎外,白姑鱼、小黄鱼等传统经济性鱼类的IRI<500.二是群落组成种类变化较小,增殖放流和季节性洄游虽然对组群间聚类水平有所提高,但作用不明显,放流前后的种类组成差异不明显(R=0.034,P>0.05),组群间聚类现象也不明显;甲壳类既是组群内相似的主要因素,又是组群间差异的主要原因. 放流后,鳌山湾、黄家塘湾、靖海湾物种丰富度指数下降,单位面积水域内一定生物量的生物个体中物种数目降低,而丁字湾、乳山湾、五垒岛湾则相反;对于种类数一定的群落总体,各种间数量分布均匀时,群落多样性指数最高.黄家塘湾的多样性指数降低,加剧了各种间数量的不均匀性,而其余放流海湾多样性指数升高,表明种间数量的均匀性升高.由于水文和盐度被认为是影响生物多样性空间分布的主要因素[11],调查期间,除黄家塘湾外其余各湾的多样性指数变化趋势基本一致,呈现上升趋势,可能是因为近岸水温回暖,部分洄游性种类洄游入湾,改变了湾内群落结构.另一方面是增殖放流活动的开展,大量三疣梭子蟹、海蜇以及中国对虾苗种相继投放入海,吸引了以放流苗种为食的生物种类的到来.加之调查期间不是捕捞旺季,尚未形成大面积捕捞力量,对群落结构影响较小.各方面因素共同作用才成了海区生物多样性指数的变化. ABC曲线可以将渔业群落结构变化与捕捞活动相联系,捕捞活动具有个体选择性,大个体生物的捕捞死亡率明显高于小个体生物,同时大个体的死亡使得小个体的被捕食压力得到释放,小个体生物会进一步增多,同时使ABC曲线发生变化.同时,最近的研究发现,外界环境和幼鱼的补充量变化也可影响生物个体大小组成使这类指标发生与对捕捞方式相同的影响,使ABC曲线发生变化.本文调查时间段前期,各海湾渔获种类组成简单,主要以中小型个体中为主,数量多但生物量较低,表现为丰度曲线高于生物量曲线,群落受到严重干扰,调查后期,随着外界资源群体的加入,降低了群落内的捕食压力,群落内个体纷纷成长起来,加之此时尚未形成大面积的集中捕捞力量,使得群落生物量提高,生物量曲线发生改变,与丰度曲线接近重合,最终发生交叉,群落恢复到中度干扰.但ABC曲线只能表明群落受到低、中、高3种干扰,评价结果往往比较粗略[10]. 参考文献: [1]李继龙,王国伟,杨文波,等.国外渔业资源增殖放流状况及其对我国的启示[J].中国渔业科技,2009,27(3):111-123. [2]金显仕.山东半岛南部水域春季游泳动物群落结构的变化[J].水产学报,2003,27(1):20-24. [3]李涛,张秀梅,张沛东,等.山东半岛南部近岸海域渔业资源群落结构的季节变化[J].中国海洋大学学报:自然科学版,2011,41(1/2):41-50. [4]山东省海洋与渔业厅.山东近海经济生物资源调查与评价[M].北京:海洋出版社,2010. [5]乔凤勤,邱盛尧,张金浩,等.山东半岛南部中国对虾放流前后渔业资源群落结构[J].水产科学,2012,31(11):651-656. [6]于振海,金显仕,李显森.黄海中南部主要鱼种的生态位分析[J].渔业科学进展,2010,31(6):1-8. [7]周红,张志南.大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J].青岛海洋大学学报:自然科学版,2003,33(1):58-64. [8]吕振波,徐炳庆,李凡,等.黄海山东海域春、秋季鱼类群落结构[J].水产学报,2011,35(5):692-699. [9]吴荣军,李瑞香,朱明远,等.应用PRIMER软件进行浮游植物群落结构的多元统计分析[J].海洋与湖沼,2006,37(4):316-321. [10]程家骅,姜亚洲.捕捞对海洋鱼类群落影响的研究进展[J].中国水产科学,2008,15(2):359-366. [11]程济生.东、黄海冬季底层鱼类群落结构及其多样性[J].海洋水产研究,2000,21(3):1-8.2 调查结果
2.1 种类组成
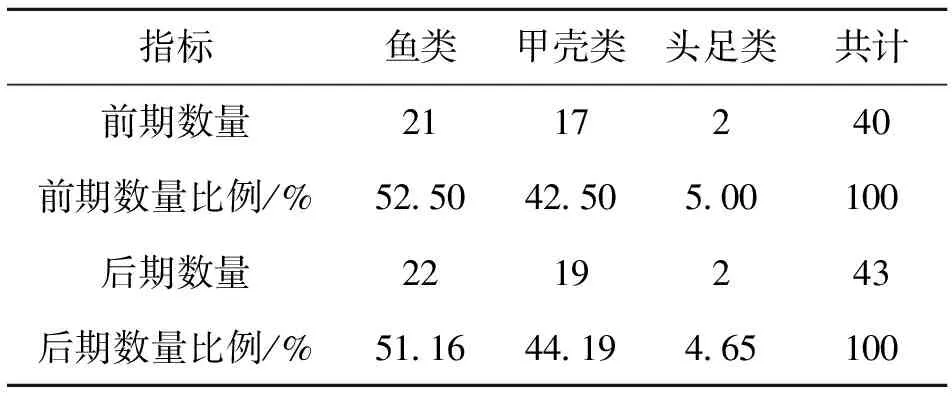
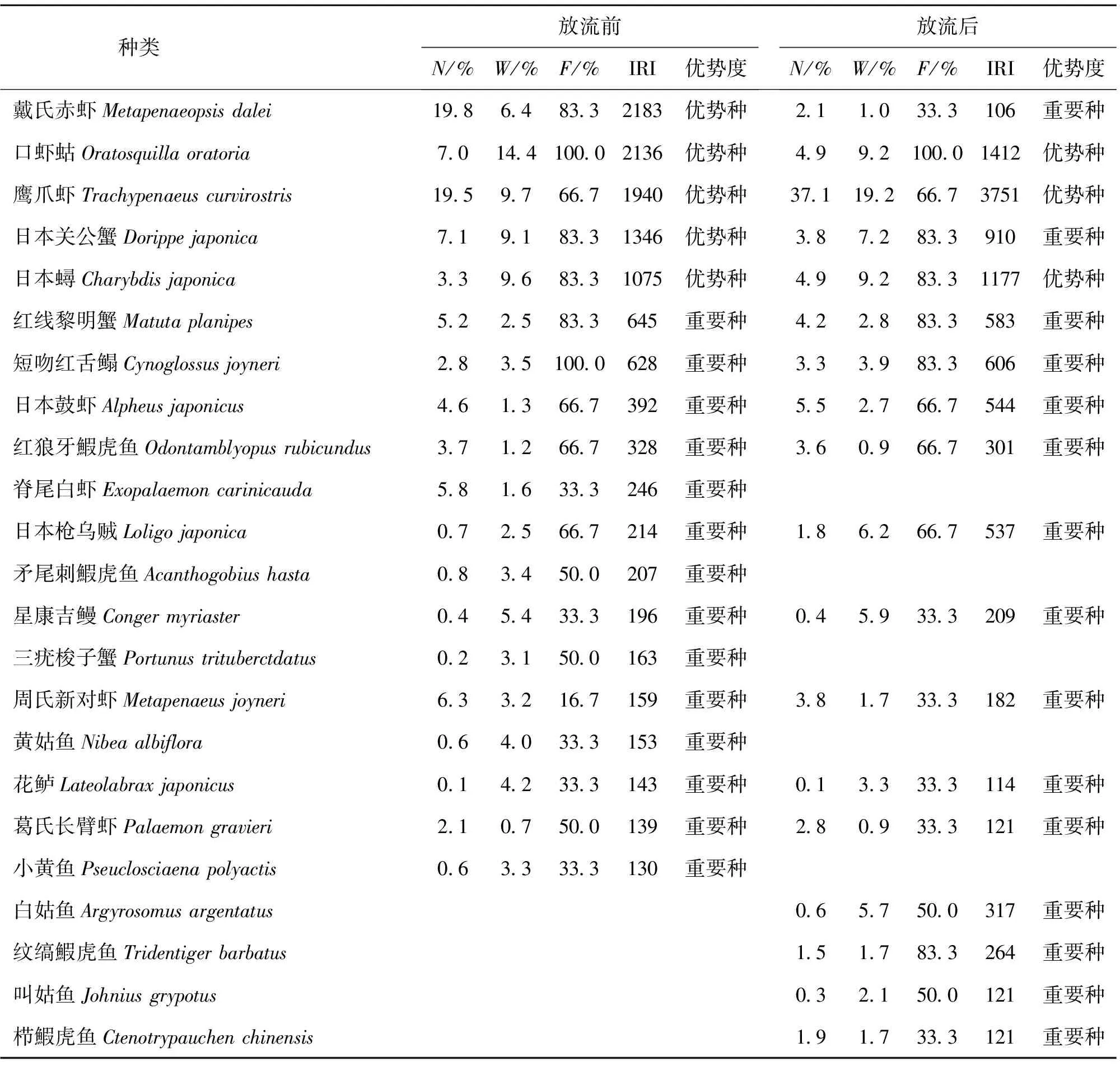
2.2 优势种
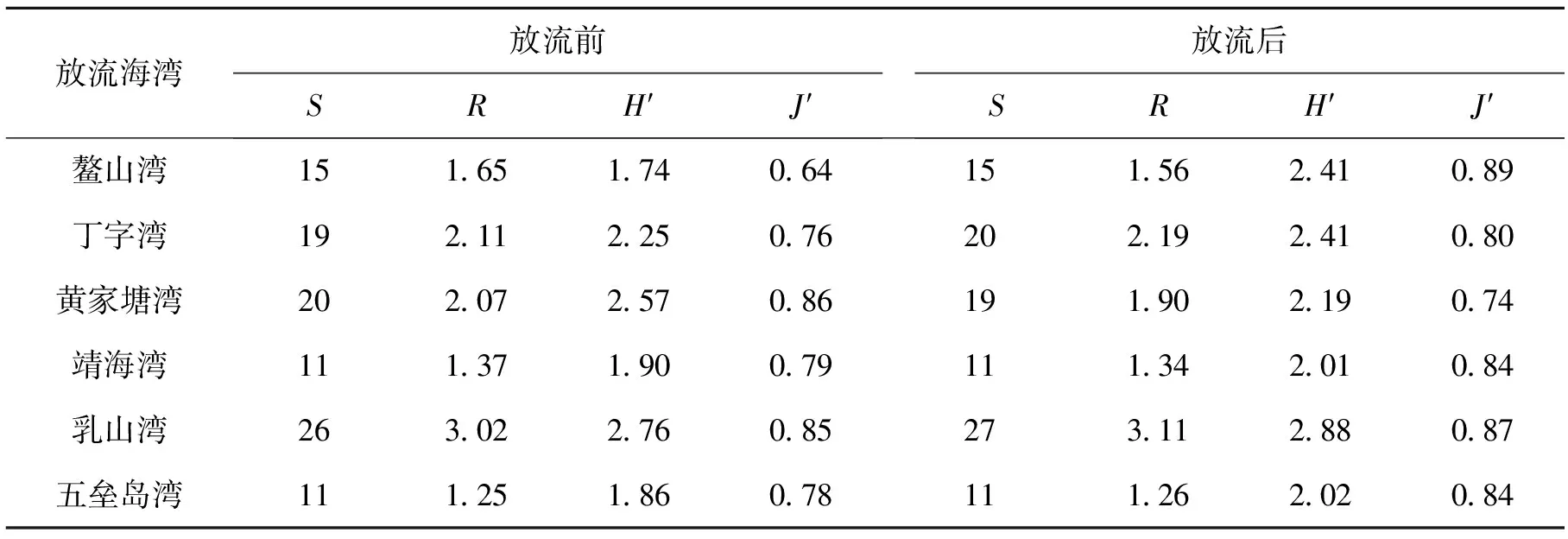
2.3 生物多样性
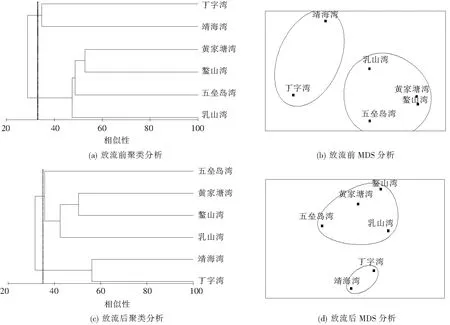
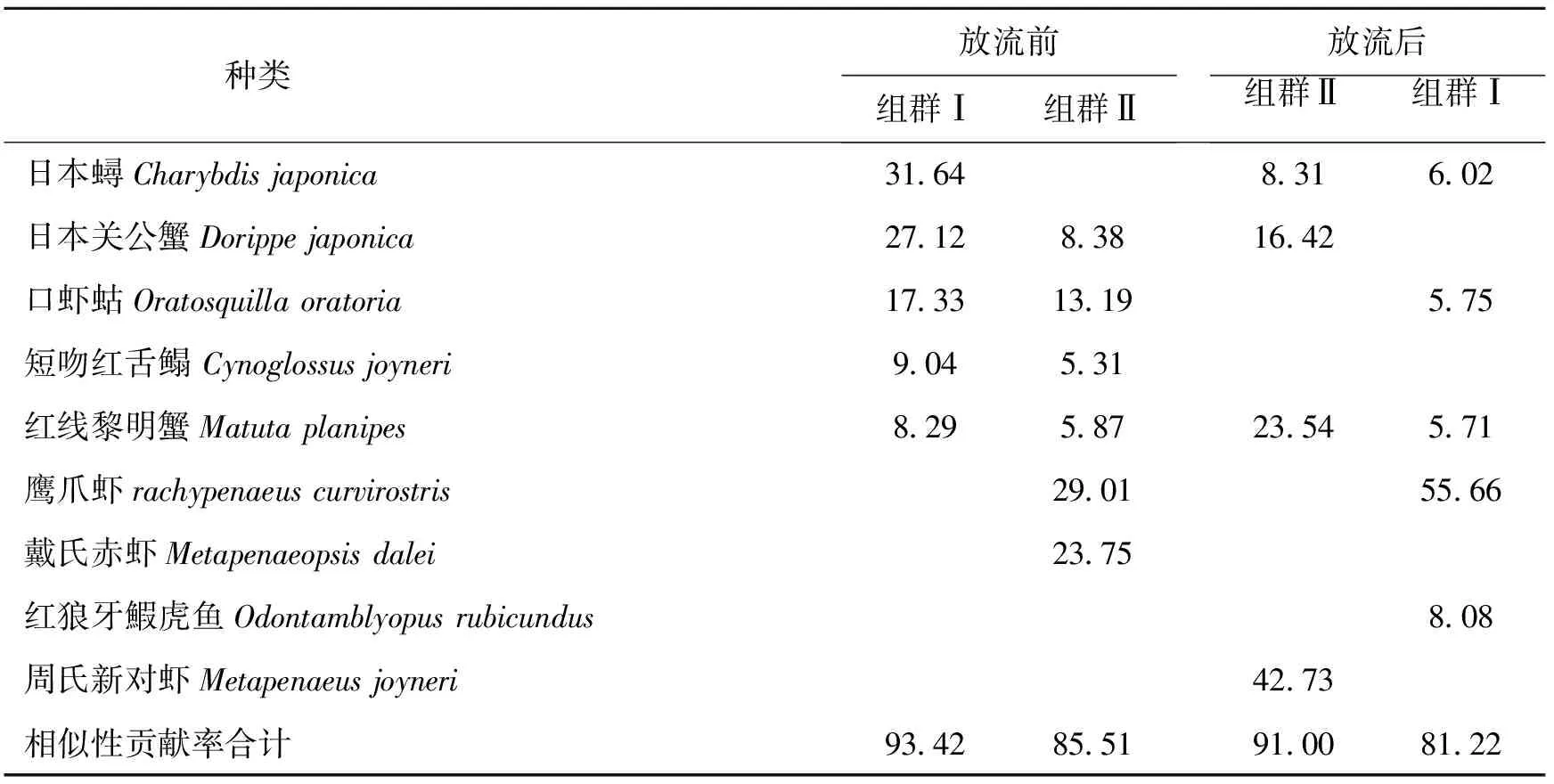
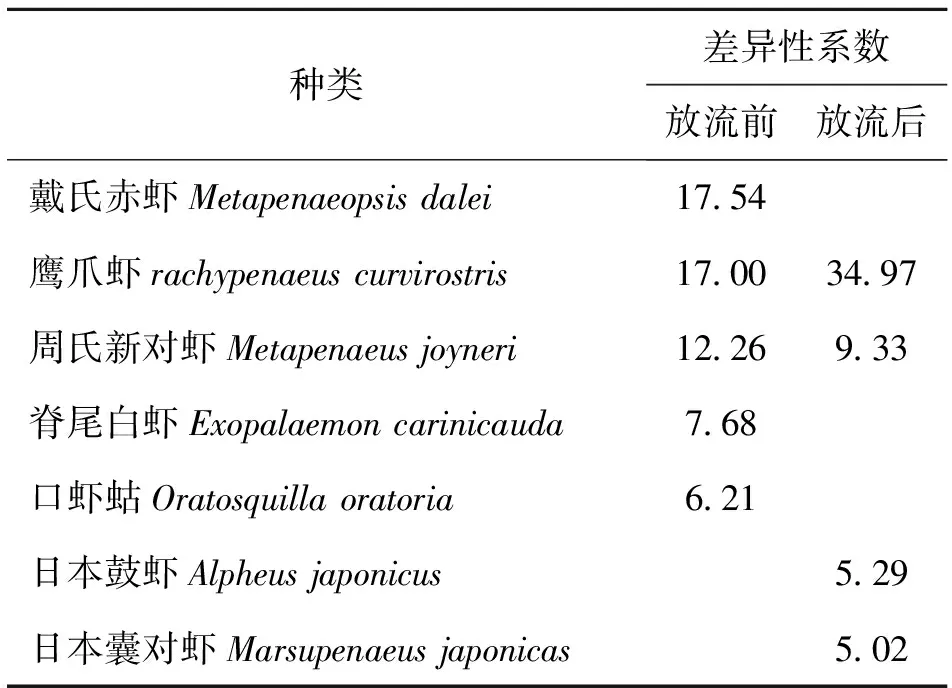
2.4 群落结构
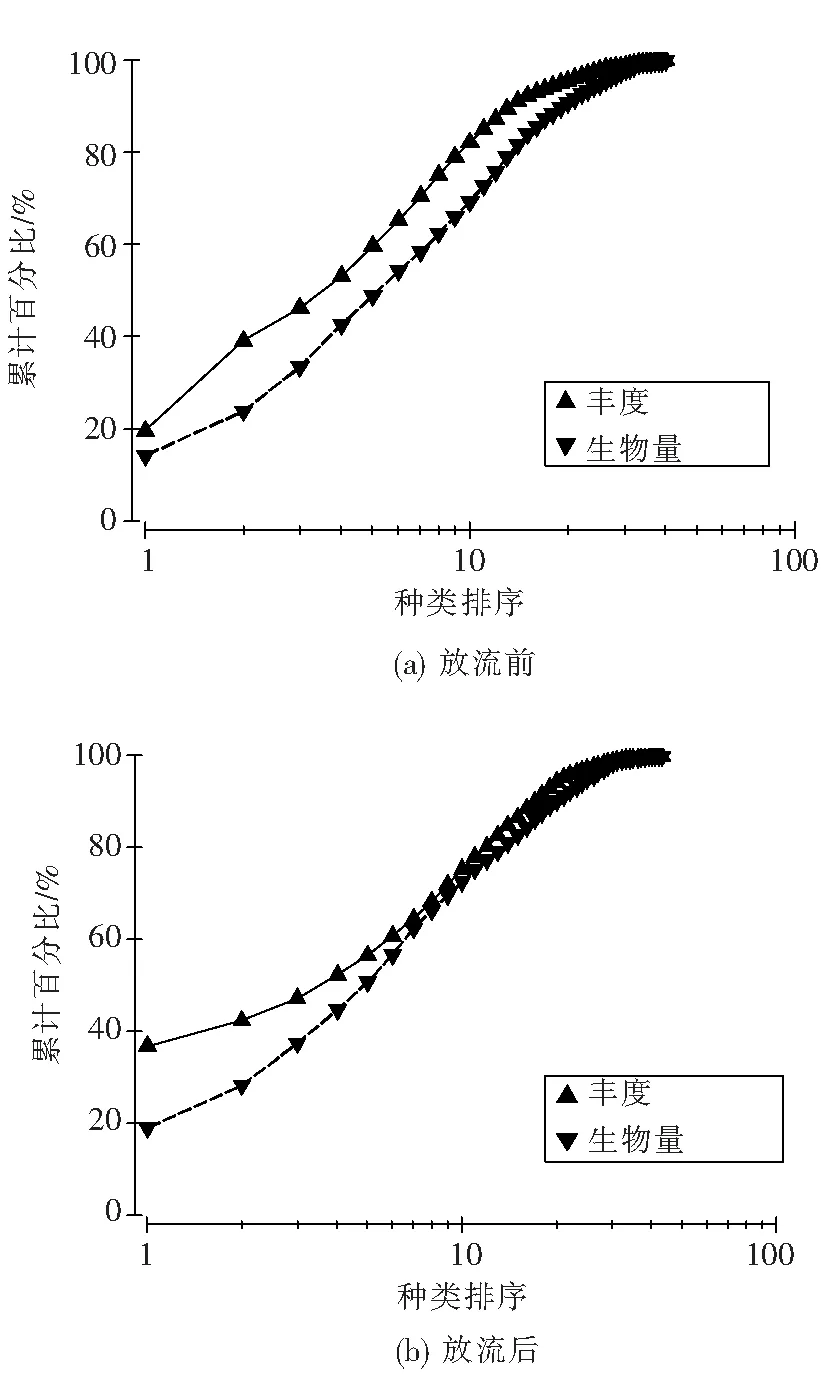
2.5 ABC曲线
3 讨 论
3.1 海区生物群落基本特征
3.2 群落结构变化分析
3.3 ABC曲线在群落结构分析中作用
猜你喜欢
现代苏州(2021年3期)2021-09-10
中国水产(2021年6期)2021-07-01
江苏教育研究(2020年28期)2020-11-23
农家之友(2020年9期)2020-04-21
浙江海洋大学学报(自然科学版)(2019年5期)2019-03-21
中国卫生质量管理(2018年6期)2018-12-04
海洋渔业(2018年5期)2018-10-26
收藏界(2018年1期)2018-10-10
创新作文(小学版)(2018年31期)2018-05-16
摄影之友(影像视觉)(2017年1期)2017-07-18
