
小鼠β-防御素-2对肿瘤细胞SiHa生长的抑制作用及其机制
2014-07-13 01:34吴红红李明远魏晓丽
新疆医科大学学报 2014年3期
郭 琼, 吴红红, 李明远, 魏晓丽
(新疆医科大学1基础医学院组织胚胎学教研室, 2附属中医医院幸福路社区医院内科, 乌鲁木齐 830000;3四川大学基础医学与法医学院微生物学教研室, 成都 610041; 4新疆医科大学基础医学院免疫学教研室, 乌鲁木齐 830011)
抗菌肽是生物体抵御外界病原生物侵袭而产生的一种小分子多肽,具有广谱的抗菌活性,并有抗真菌、病毒等生物学活性,而对正常细胞无伤害,是机体在长期进化过程中保留下来的一种古老的宿主防御机制[1]。哺乳动物体内的抗菌肽主要是防御素(defensins),作为生物体内天然防御系统的重要组成部分,连接天然免疫和获得性免疫反应,对机体抵抗外界生物感染起着关键的作用[2]。此外,许多科学研究表明防御素也具有抗肿瘤效应[3-4],但其机制尚不完全清楚。现阶段已发现的500多种抗菌肽中,有18种抗菌肽的抗肿瘤活性已得到实验证实。宫颈癌是倍受世界关注的女性高发肿瘤之一,其发病率和死亡率居高不下,在全球位列女性恶性肿瘤的第二位,宫颈癌的防治成为保护妇女儿童健康的全球共同关注问题。随着生物技术的发展和肿瘤发生分子机制的深入研究,生物治疗已成为肿瘤综合治疗的第4种模式,并具有广泛的应用前景。本课题前期研究工作中,采用pcDNA3.1(+)/mBD2和具有明确穿膜功能的小鼠IgGΚ链的信号肽基因替代mBD2信号肽基因,形成IgGΚ链、成熟肽2个部分的重组防御素分子质粒pcDNA3.1(+)/rmBD2转染SiHa细胞,制备了宫颈癌细胞瘤苗SiHa/M、SiHa/R[5-6]。本研究对细胞的特性进行了体外研究,为进一步观察其体内抗宫颈癌效果奠定基础,同时为将防御素应用于肿瘤免疫治疗提供实验依据。
1 材料与方法
1.1材料
1.1.1 细胞株及质粒 稳定转染pcDNA3.1(+)/mBD2质粒、pcDNA3.1(+)/rmBD2质粒宫颈癌细胞株SiHa/M、SiHa/R本室保存,宫颈癌SiHa细胞由四川大学华西医学中心免疫学教研室魏大鹏老师惠赠,用RPMI1640细胞培养基、10%胎牛血清在37℃、50 mL/L CO2条件下进行培养。
1.1.2 试剂 RPMI1640、胎牛血清(GIBCO公司),PVDF膜(invitrogen公司),5 %脱脂奶粉(invitrogen公司),羊抗鼠mBD2抗体 (Santa cruz公司),兔抗羊IgG抗体、DAB(Fermentas 公司),Annexin Ⅴ、PI、Rnase酶(Bioscience公司),TUNEL试剂盒(Roche公司),细胞活力分析采用BECKMAN VICELLA AUTO 100/240进行测定。
1.2方法
1.2.1 细胞裂解上清mBD2蛋白表达 收集并裂解各组转染细胞提取总蛋白,用考马斯亮蓝法蛋白定量后进行Tricine-SDS-PAGE (Tricine sodiumdodecyl sulfatepolyacry lamidegele lectrophoresis , Tricine-SDS- PAGE )电泳,分离蛋白,采用湿转法将蛋白质转移至聚偏氟乙烯( polyvinyli denedifluoride,PVDF)膜,5%脱脂奶粉封闭过夜后,分别用1∶100稀释羊抗鼠mBD2抗体孵育2 h,TBST洗膜3次(10 min /次),再用1∶1 000稀释兔抗羊IgG抗体孵育1 h,TBST洗膜3次(10 min /次),DAB显色检测结果。
1.2.2 细胞活力测定 取对数生长期的稳定转染细胞及未转染的SiHa细胞,计数并调整细胞浓度为1×105个/mL,分别接种于12块6孔板中,每组细胞设3个复孔。在含有10%胎牛血清的RPMI1640培养液、5%CO2的条件下培养。细胞贴壁后于每天同一时间分别消化3组细胞,用无血清培养基重悬后加入测量杯中,于细胞活力分析仪上进行细胞活力测定,并绘制细胞活力曲线。
1.2.3 PI、Annexin Ⅴ双标检测细胞凋亡 将各组细胞用胰酶消化离心收集细胞, PBS洗涤并用台盼蓝染色测定活细胞百分比,用PBS将细胞数调整为1×106个/mL。分别用含Rnase的Annexin Ⅴ染色液、PI避光染色各30 min,流式细胞仪检测并记录实验结果。
1.2.4 TUNEL法测定细胞凋亡 将对数生长期细胞以1×105个/mL于6孔板中进行细胞爬片培养至细胞铺满60%左右,用冰预冷的95%乙醇于-20℃固定20 min,室温中晾干;依次在100%、95%、70%的乙醇依次脱水2 min,并采用3%H2O2甲醇溶液室温30 min,0.1%Triton X-100于4℃ 2 min,滴加正常羊血清,室温孵育10 min,滴加TdT 及FITC 标记的核苷酸混合反应液50 μL,37℃ 60 min后终止标记反应(1×TdT Stop Buffer),并采用DAPI复染,荧光显微镜观察凋亡结果并计算凋亡指数。凋亡指数按照在显微镜高倍镜下(×400)观察5个视野,每个视野数100个细胞,计算凋亡细胞所占百分比,即为凋亡指数(AI)。
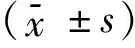
2 结果
2.1细胞裂解上清Westernblot分析结果显示,SiHa/M、SiHa/R组在4~6 Ku区域出现目的条带,而SiHa组无目的条带(图1)。

图1 细胞裂解上清的Western blot分析
2.2mBD2对细胞活力的影响第1天SiHa/M、SiHa/R组与SiHa组比较,细胞活力差异无统计学意义(P=0.091);第2、3天SiHa/M、SiHa/R组细胞活力明显高于对照组,差异有统计学意义(P=0.018);第4、5天3组细胞活力差异无统计学意义(P=0.374)(图2)。
2.3mBD2对细胞凋亡的影响
2.3.1 Annexin Ⅴ联合PI检测结果 结果显示SiHa/M、SiHa/R及SiHa组Annexin Ⅴ+/PI+双阳性细胞的百分率分别为2.4%、2.3%、2.3%,差异均无统计学意义(P=0.743);Annexin Ⅴ+/PI-细胞分别为0.9%、1.2%、1.5%,差异无统计学意义(图3)。

*P<0.05,compared with SiHa at the same time图2 稳定转染细胞的细胞活力分析结果
2.3.2 TUNEL检测结果 细胞大小形态不一,可见凋亡细胞胞核固缩浓染,核质致密,出现核边缘化,呈半月形;正常细胞胞核相对疏松,无核浓染现象(图4~6)。SiHa/M、SiHa/R组凋亡细胞发生率较高,分别为(40.6±4.67)%、(30.2±3.19)%,而SiHa组只有(7.4±1.94)%,SiHa/M、SiHa/R组凋亡细胞发生率明显高于SiHa组,差异有统计学意义(P=0.004);SiHa/M组与SiHa/R组比较,SiHa/M组凋亡发生率高于SiHa/R组,差异有统计学意义(P=0.003)。

A: SiHa组 B: SiHa/M组 C: SiHa/R组
图3AnnexinⅤ检测各组细胞的凋亡结果

A: DAPI 染色 B: FITC染色 C: A、B重叠结果
图4SiHa/M凋亡结果(×600)

A: DAPI 染色 B: FITC染色 C: A、B重叠结果
图5SiHa/R凋亡结果(×600)

A: DAPI 染色 B: FITC染色 C: A、B重叠结果
图6SiHa凋亡结果(×600)
3 讨论
机体对“非己”的免疫反应需要天然免疫和获得性免疫系统的协同作用。天然免疫反应不仅提供了抵抗微生物的第一道防线,同时也包含指导获得性免疫所必需的“危险信号”。防御素是宿主执行免疫防御能力的细胞所分泌的、具有杀微生物活性和细胞毒性的小分子多肽,提供直接、即刻的抗微生物效应以保护宿主免受入侵病原微生物损害,是固有免疫中重要的效应分子和调节分子,广泛用于抗菌、抗病毒等作用的研究。自首次报道防御素能裂解肿瘤细胞之后,不断有研究提示防御素与肿瘤免疫有关[7-9]。研究表明,mBD2可与细胞表面CCR6受体结合,呈剂量依赖性的诱导未成熟树突状细胞(dendritic cells,DCs)CD34+祖细胞源性DCs的移动[10]。进一步的研究显示,mBD2可与Toll样受体4 (toll like receptor,TLR)结合,诱导骨髓源性DCs成熟[11]。此外,由于肿瘤细胞膜表面特性发生改变而使得防御素能够与之结合并产生抗瘤作用也可能是防御素抗肿瘤的机制之一。防御素对肿瘤细胞的杀伤作用远大于非肿瘤细胞,但这种选择性杀伤的机制目前尚不清楚[12]。
本研究通过细胞活力测定,显示SiHa/M、SiHa/R组细胞在贴壁后2~3 d细胞活性明显低于正常SiHa细胞。随后,随着细胞数目增加,培养液中营养物质消耗增加,细胞代谢产物堆积,细胞活性开始下降,提示mBD2蛋白对细胞活力有一定的抑制作用。这可能与防御素作用于细胞线粒体从而影响肿瘤细胞呼吸、导致细胞代谢紊乱、细胞活力下降有关。SiHa/M、SiHa/R组细胞活力比较,SiHa/M始终低于SiHa/R组,但两者差异无统计学意义,表明2种蛋白信号肽分子对蛋白活性影响不大。
本研究采用Annexin V 联合PI法和TUNEL 2种方法测定转染细胞的凋亡情况。2种方法分别检测凋亡发生的不同时相,反映诱发凋亡的不同机制。采用Annexin Ⅴ检测方法观察细胞凋亡,发现3组细胞凋亡指数无明显差别;采用TUNEL染色,结果显示SiHa/M、SiHa/R组凋亡细胞发生率较高,与对照组相比差异有统计学意义;SiHa/M组与SiHa/R组比较,SiHa/M组凋亡发生率高于SiHa/R组。
综上所述,可以认为mBD2蛋白所诱导的SiHa细胞凋亡主要以肿瘤细胞DNA出现断裂损伤为主,这与Hariton等[13]研究结果一致。而由于细胞膜损伤导致PS外翻通过Annexin Ⅴ检测方法未能得到验证,可能是由于早期凋亡所产生的PS外翻过程较为短暂,采用Annexin Ⅴ未能捕捉到早期凋亡信号,故检测阳性率较低。由于SiHa/M与SiHa/R的主要区别在于两者所转染的质粒中插入序列信号肽区域基因有所不同,是否信号肽及其调控区序列在成熟肽形成中具有一定的调节功能,从而对成熟肽功能产生一定影响,还需实验进一步研究证实。
参考文献:
[1] Andreas RK, Robert B. Antimicrobial peptide current status and therapeutic potential[J]. Drugs,2003, 63(4):389-406.
[2] Yurong Y, Yibao J, Ruiping S, et al. Effects of chicken intestinal antimicrobial peptides on humoral immunity of chickens and antibody titres after vaccination with infectious bursal disease virus vaccine in chicken[J]. AAN-Archiv Fur Tierernahrung, 2006, 60(5):427-435.
[3] Chen JY, Lin WJ, Wu JL, et al. Epinecidin-1 peptide induces apoptosis which enhances antitumor effects in human leukemia U937 cells[J]. Peptides, 2009, 30(12):2365-2373.
[4] Hoskin DW, Ramamoorthy A. Studies on anticancer activities of antimicrobial peptides[J]. BBA-Biomembranes, 2008,1778(2):357-375.
[5] 魏晓丽,施桥发,李虹,等.鼠β-防御素2(mBD2)真核表达质粒的构建及稳定表达株的鉴定[J].细胞与分子免疫学杂志,2007,23(1):28-31.
[6] 魏晓丽,李明远,李虹,等.信号肽重组mBD2在宫颈癌细胞中的稳定表达及活性测定[J].四川大学学报:医学版,2007,38(2):222-225.
[7] Presicce P, Giannelli S, Taddeo A, et al. Human defensins activate monocyte-derived dendritic cells, promote the production of proinflammatory cytokines, and up-regulate the surface expression of CD91[J]. J Leukoc Biol, 2009, 86(4):941-948.
[8] Lapteva N, Aldrich M, Rollin SL, et al. Attraction and activation of dendritic cells at the site of tumor elicits potent antitumor immunity[J]. Molecular Therapy, 2009, 17(9):1626-1636.
[9] Biragyn A, Coseia A, Nagashima K, et al. Murine beta-defensin 2 promotes TLR-4/MyD88-mediated and NF-kappaB-dependent atypical death of APCs via activation of TNFR2[J]. J Leukoc Biol, 2008, 83(4):998-1008.
[10] Kulkarni MM, McMaster WR, Kamysz W, et al. Antimicrobial peptide-induced apoptotic death of leishmania results from calcium-de pend ent, caspase-independent mitochondrial toxicity[J]. J Biol Chem, 2009, 284(23):15496-15504.
[11] Kesting MR, Loeffelbein DJ, Hasler RJ, et al. Expression profile of human beta-defensin 3 in oral squamous cell carcinoma[J]. Cancer Invest, 2009, 27(5):575-581.
[12] Lapis K. Host defense peptides and peptide in etics as new weapons for Cancer treatment[J]. Magyar Onkologia, 2010, 54(1):47-58.
[13] Hariton-Gazal E, Feder R, Mor A, et al. Targeting of nonkaryophilic cell-permeable peptides into the nuclei of intact cells by covalently attached nuclear localization signals[J]. Biochem, 2002, 41(29):9208-9214.
猜你喜欢
生物技术通报(2021年6期)2021-08-11
江西农业学报(2021年4期)2021-04-20
当代陕西(2020年9期)2020-08-04
浙江农业学报(2017年3期)2017-04-08
中国烟草学报(2016年1期)2016-11-16
中国工程咨询(2016年6期)2016-01-31
中国卫生(2015年7期)2015-11-08
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
