
种群密度和食物密度变化对蒙古裸腹溞产雄的诱导效应
2014-07-12 08:19赵满意王伟洪张健兵钟爱华储张杰
浙江海洋大学学报(自然科学版) 2014年1期
赵满意,王伟洪,张健兵,钟爱华,储张杰,楼 宝
(1.浙江海洋学院水产学院,浙江舟山 316022;2.浙江省海洋水产研究所,浙江舟山 316100)
蒙古裸腹溞是目前唯一可在海水中大量培养的盐水枝角类[1],作为卤虫的替代产品,其在海水鱼苗培育中的应用和生产性培养正日益受到重视。种群的数量变动理论上是出生率与死亡率的变动,其中出生率与生殖量、种群性比等指标相关,因此,雄体产生机理和诱导因素的研究,是研究枝角类等生物饵料种群数量动态变化机理和规模化培养研究的重要内容。一般认为,枝角类孤雌生殖和两性生殖两种生殖方式均与环境条件相关[2-5],但至今关于枝角类的生殖机制研究不多,尤其是两性生殖机理很不清楚。相关的前期研究表明[1],蒙古裸腹溞的雄体来源于母体的产出,进行孤雌生殖的母体在良好环境条件(温度、盐度和食物等适宜,低溞密度条件)下,产出的幼体几乎都是孤雌生殖雌体,产雄率极低,而随着溞密度的上升及温度和饵料密度等因子的改变,产雄率变化很大。本研究在这些相关研究基础上,着重研究较高温度条件下(25~30℃),种群密度和食物密度变化对蒙古裸腹溞产雄的诱导效应,以期进一步掌握蒙古裸腹溞的繁殖特性,并为蒙古裸腹溞的规模化培养和枝角类生殖生理等研究提供参考资料。
1 材料与方法
1.1 溞种来源
实验溞种由上海海洋大学生理生态重点实验室提供,源于1982年采自晋南地区半咸水湖,在浙江海洋学院水产养殖实验室内以海水连续孤雌生殖繁衍保存下的溞种。
1.2 实验条件与饵料
实验在 RXZ 人工气候箱内进行,设定温度为(28±0.5)℃,光照 3 000~4 000 lx,光暗周期(L:D)为 12:12;盐度27.5±0.5;培养液由实验条件下培养至指数生长期的海水小球藻Chlorella sp.配制而成。
1.3 实验设计和方法
1.3.1 实验设计
实验设初始溞数量为1 ind/mL(低溞密度)、3 ind/mL(中溞密度)、6 ind/mL(高溞密度)三个水平,按培养液更换间隔设固定和机动两组,固定组设1次/12 h和1次/6 h更换培养液两组,机动组是藻液更换次数视藻液滤淡(清)情况而定。各实验组均设3个重复。
1.3.2 实验用溞
实验用溞均为实验条件下,0.2 ind/mL密度下4 h内产出的同龄幼体。在更换的藻液密度 (3~4)×106cells/ml,24 h更换培养液一次的实验条件下,选取单个培养连续3个世代以上的溞1个,产出第一窝幼溞后,移走母溞,培养幼溞,并以同样方法,始终控制培养密度在0.2 ind/mL以下,扩大培养至第一窝产出幼溞能满足实验用溞数量需要。实验开始时,选取在4 h内基本同步产出的幼溞,挑至1只1 000 mL烧杯中混合,然后再用吸管随机吸至各实验组。
1.3.3 实验方法
实验均在50 mL烧杯中进行,控制培养容积误差(50±5)mL,溞密度为低、中、高各处理初始实验组幼溞数分别为50个、150个、300个,培养液(藻液)更换密度均为(3~4)×106cells/mL。每次产幼前,及时随机抓取育儿囊高起快产幼的溞10个,放至空白烧杯继续培养至幼体全部产出后,将母溞放回原烧杯,计数产出幼溞总数和其中的雄体数,另在固定1次/12 h各溞密度组产第一窝时,随机抓取育儿囊高起快产幼的溞27个(即每个烧杯抓取3个母溞),在空白烧杯中1杯1个培养至幼体全部产出后,将母溞放回原烧杯,计数产出幼溞总数和其中的雄体数。实验至第四窝雌雄鉴别后结束。
1.4 数据处理与分析
窝产幼数=每窝产出幼体数
总产幼数=每窝产出幼体数之和
窝生殖量=每窝产出幼体数/(存活溞总数-形成的两性生殖雌体数)
窝雄体百分率=每窝产出幼体中的雄体数/每窝产幼数(窝生殖量)×100%
平均雄体百分率=累计产出幼体中的雄体数/累计产幼总数(总生殖量)×100%
窝平均产出时间=Σ(次产幼数×次产幼时间)/窝产幼数,次产幼数指每次观察时已产出的幼体数,次产幼时间均取相邻检查记录时间的中值
雄体产出效率=(累计)产幼总数×(平均)雄体百分率/平均产出时间×1 000 mL/50 mL(ind/d·L),指 1 L水体平均每天产出的雄体数;其中,平均产出时间=Σ(次产幼数×次产幼时间)/累计产幼总数
实验数据方差分析取P<0.05为差异显著标准,均值比较均按Duncan法分析。
2 结果
2.1 不同培养条件下的雄体产出率
至第4窝产幼结束,不同培养条件下的平均雄体产出率范围为0.14%~13.01%,以溞密度3 ind.ml-1×培养液更换1次/12 h组最高,溞密度1 ind/mL×培养液机动更换组最低。从图1和图2可以看出,平均产雄率,相同培养液更换条件下,不同溞密度间差异较大,以中溞密度(3 ind/mL)最高,高溞密度(6 ind/mL)其次,低溞密度(1 ind/mL)最低;而在相同溞密度条件下,以固定组1次/12 h最高,固定组1次/6 h和机动组则差异不大,约为固定组1次/12 h的50%左右。从图3可知,不同培养液更换条件,固定组1次/12 h和1次/6 h均表现为中溞密度>高溞密度>低溞密度规律,尤以固定组1次/12 h的差异更明显,机动组则表现为随溞密度降低而降低规律,而在不同溞密度下,中、低溞密度时均表现为固定组1次/12 h>固定组1次/6 h>机动组规律,高溞密度时则表现为机动组﹥固定组1次/12 h>固定组1次/6 h。方差分析结果表明,溞密度和培养液更换方式对平均产雄率有较强的交互作用。
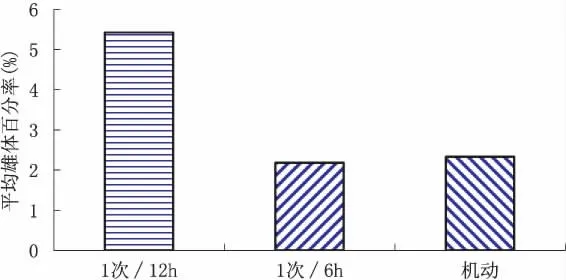
图1 不同培养液更换方式下至第4窝的平均雄体百分率Fig.1 Average male ratio of 4 broods neonates in different algae replace
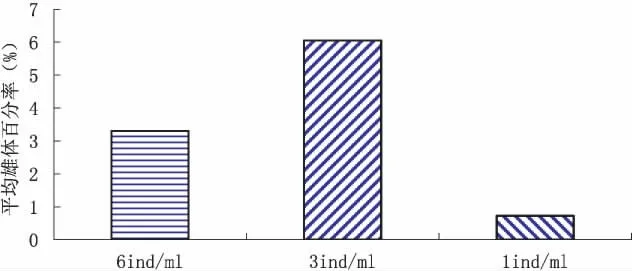
图2 不同溞密度下至第4窝的平均雄体百分率Fig.2 Average male ratio of 4 broods neonates in different population density

图3 不同培养条件下至第4窝的平均雄体百分率Fig.3 Average male ratio of 4 broods neonates in different experimental combination
2.2 不同培养条件下的窝雄体产出率
由表1可知,不同培养条件下,第1窝均有雄体产出,且窝产雄率差异极大,变动范围 0.54%~29.00%,以溞密度 3 ind/mL、培养液更换1次/12 h组最高,溞密度1 ind/mL、培养液机动更换组最低;第2窝只有部分组别有雄体产出,且产出率远低于或低于同组别第1窝,而第3、第4窝各组别均未发现有雄体产出。
由表2可知,在27个单个母溞的第1窝产出幼体中,有13个母溞产出幼体全部都是雌体(雄体产出率为0),占全部母溞的48.15%;其他14个都有雄体产出,占全部母溞的51.85%,窝雄体产出率最高可达60.00%,最低16.67%,其中高于或等于50%的有3个,40%~49%的有2个,30%~39%的有3个,低于30%的6个,但没有1个母溞1窝产出幼体是全雄的。

表1 不同培养条件下的窝雄体百分率(%)Tab.1 Male ratio of different broods in different experimental combination(%)
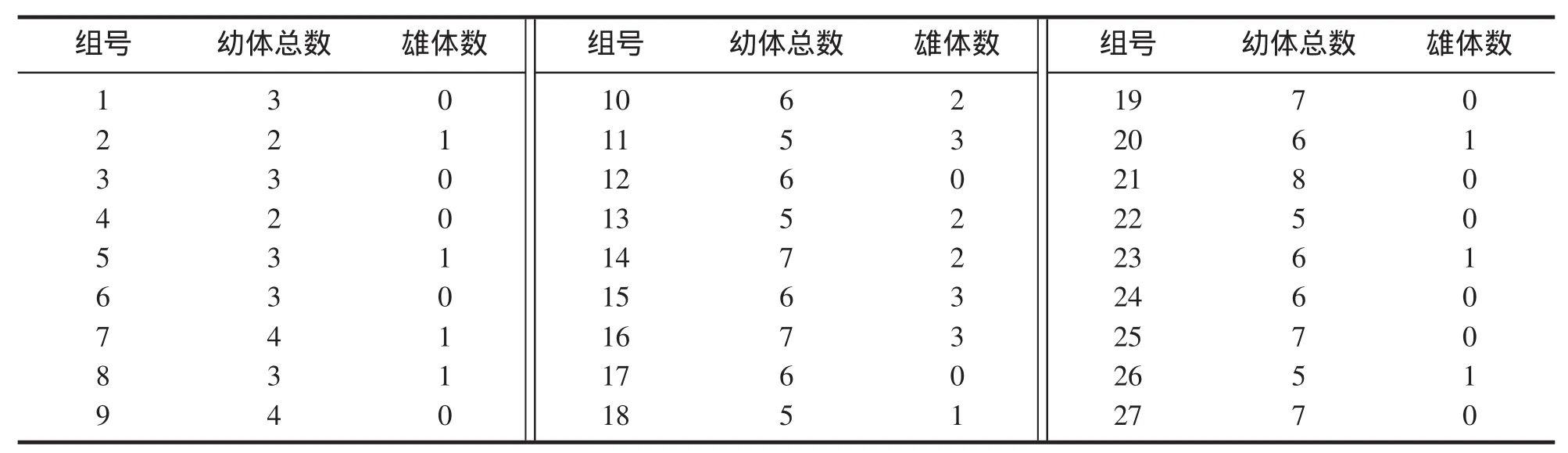
表2 固定组1次/12 h×3 ind/mL组合的第1窝幼体数和雄体数Tab.2 The number of neonates and males in the first brood of 1times/12 h×3 ind/mL combination
2.3 不同培养条件下的雄体产出效率
由于雄体的产出主要集中于第1窝,第3窝和第4窝均未有雄体产生,不同培养条件下的雄体产出效率基本上都随培养时间增加而降低。由表4可知,至第一窝产出,雄体产出效率范围为0.15~7.78 ind/(d·L),以固定组1次/12 h×3 ind/mL组合最高,机动组×6 ind/mL组合其次,机动组×1 ind/mL组合最低,至第4窝产出,最高的固定组1次/12 h×3 ind/mL组合从7.78 ind/(d·L)下降到3.52 ind/(d·L)。
从图4可以看出,至第4窝的平均雄体产出效率,相同溞密度时,高溞密度下,培养液更换两固定组差异不显著,机动组显著高于二者;低溞密度下,三组均较低,机动组接近于0,两固定组略高于机动组且两者差异不显著;中溞密度下,三组相对均较高,其中固定组1次/6 h与机动组差异不显著,但固定组1次/12 h远高于二者;相同培养液更换方式下,两组固定组均表现为中溞密度时最高,高溞密度和低溞密度均较低且两者间差异不显著,而机动组则表现为随溞密度降低而显著降低。
3 讨论
3.1 蒙古裸腹溞的产雄特性
曹文清等[6]通过染色体组型研究发现,蒙古裸腹溞的幼体在育儿囊中已产生雌雄性分化,即雄体在胚胎发育期形成,但胚胎发育的雌雄性分化是卵的差异还是胚胎发育过程中受外界环境因素诱导所致,目前尚不清楚。关于枝角类雄体产生的机理,是内部因素控制的生存对策,还是环境因子的诱导作用,抑或两者的共同作用,不同学者间存在分歧[8-18]。在本研究实验用溞制备过程中,检测单个母溞的窝生殖量和产雄率时发现,窝生殖量8~13个,产出幼体几乎都是孤雌生殖雌体,偶有雄体发现,即在很低溞密度和相对较高藻密度情况下,有个别母溞可有雄体产出,但产出率极低。依据本研究实验结果,作者初步分析认为,蒙古裸腹溞雄体的产出应与母溞个体有关,不同个体产雄率的差异主要在于个体差异和诱导因素强弱,排入育儿囊的不同卵之间可能不同质,即可能存在基因型及卵的大小、卵黄含量多少等差异,不同卵在胚胎发育时也可能不均等,这些因素都可能导致初生幼溞的个体差异,由此,不同个体在生长发育过程中对外界环境条件的反应敏感性有一定差异,从而导致相同条件下个体间存活率、寿命、生殖量、产雄率等差异。
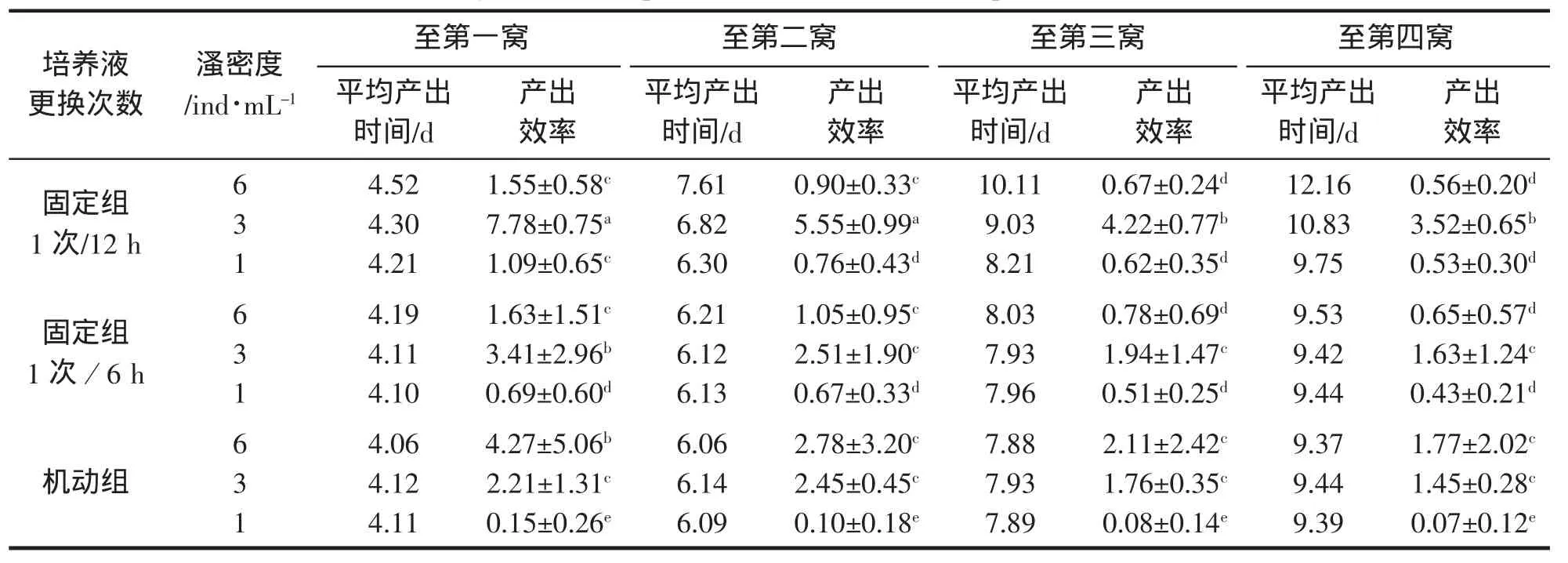
表3 不同培养条件下的雄体产出效率Tab.3 Efficiency of male reproduction in different experimental combination

图4 不同培养条件下至第4窝的雄体产出效率Fig.4 Efficiency of male reproduction in different experimental combination
在本研究检测的27个母溞中,有48.15%的母溞一窝幼体是全雌的,但没有1个母溞一窝幼体是全雄的,这与卢芳野[7]等在食物不足、低溞密度及拥挤水等条件下,出现雄体的72窝中,全是雄体的窝数占65%的研究结果完全不同。处理本研究实验数据时发现,不同培养条件下的平均窝生殖量表现为随溞密度增加而下降,随培养液更换次数增加而增加,变动范围2.14~6.02个,在溞密度6 ind/mL-1、培养液更换1次/12 h组合中,平均窝生殖量已降至2~3个,但也没有发现有产全雄的窝数。蒙古裸腹溞是否具产全雄窝次特性及单依赖溞密度和藻密度波动是否能诱导蒙古裸腹溞产全雄窝次尚有待进一步研究。至于同一个体第2窝之后产雄率的降低或不产雄,作者认为这可能是母溞的环境适应性所致,环境因子诱导母溞产出雄体必定与母溞对环境因子的敏感性有关,母溞从自身产出到产第一窝幼体处于幼龄至成龄生长发育期,是发育的关键阶段,可能对环境因子的变化较敏感,从而易导致雄体产生,而从第二窝起则都是成龄生长期,经历幼龄期的环境适应后,母溞对环境因子的敏感性可能已下降或已不敏感,处于“相对适宜”的环境条件状态而使产雄率下降或不产雄。改变环境因子的强度是否能提高母溞第2窝以后的雄体产出率尚有待进一步研究。
3.2 不同培养条件下蒙古裸腹溞的产雄率与产雄效率
本研究中,窝产雄率指每窝产出幼体中的雄体百分率,与每窝中的雄体产出数和产出幼体总数即窝生殖量相关,反映一定条件下的单窝产雄特性;平均产雄率是指至第4窝产幼结束时累计产出幼体中的累计雄体百分率,反映整个生殖过程的产雄特性。本研究结果表明,不同培养条件下,蒙古裸腹溞雄体的产出基本集中于第1窝,第2窝至第4窝产出雄体数大幅下降或无,4窝的平均产雄率随窝次增加而降低,这反映在特定培养群体中的雄体比率必然随培养时间波动,在种群增长前期很低,快速增长期和相对生长下降期相对较高,相对稳定期则又较低。
雄体产出效率,即1 L水体平均每天产出的雄体数,由单位时间内的累计产幼总数、平均雄体百分率和平均产出时间等因素决定。检测过程发现,中、高溞密度下有4%~11.89%的母溞转化形成两性生殖雌体,低溞密度下则无或偶有产生,这导致中、高溞密度下实际可产幼孤雌生殖母溞数下降。另外,从表4可知,幼体的平均产出时间有随溞密度增加而增加,随培养液更换次数增加而降低趋势,在第1窝生殖时差异基本不显著,至第2窝最大差异1.55 d,至第4窝则可达近3 d的差异。低溞密度时窝产雄率和平均产雄率低,高溞密度时可产幼孤雌生殖母溞数和窝生殖量均下降,产幼总数降低,同时幼体平均产出时间增加等,由此,雄体产出效率在中溞密度时相对较高,而雄体产出基本集中于第1窝的特性,使雄体产出效率随培养时间增加而降低。
3.3 溞密度和藻密度对蒙古裸腹溞产雄的诱导效应
本研究结果充分表明溞密度和藻密度波动是诱导蒙古裸腹溞雄体产生的两个重要因素。对雄体产出率,溞密度和培养液更换方式两者有较强交互作用,低溞密度下,不同培养液更换方式下均很低,但中、高溞密度时则与培养液更换方式相关。作者分析认为,溞种群的密度效应包含溞个体间的碰撞频率和代谢分泌物浓度效应,与溞种群的数量、个体大小、运动能力与强度等有关,并受温度、光强、藻密度等因素影响,而在固定更换藻液密度条件下,培养液更换方式实质影响的是藻密度的波动大小、溞群体代谢分泌物浓度变化及培养液pH等整体环境因子,使这些因子呈现周期性动态变化规律,培养液更换间隔越短,波动越小,间隔越长,波动越大,其中,影响最大的是藻密度的波动幅度。由于溞群体的滤食效应和藻细胞的生长繁殖,低溞密度下,溞群体的滤食效应与藻细胞的生长繁殖速度可能差异不大,在一个更换期内藻密度的波动均较小,而高溞密度下,溞群体的滤食效应强于藻细胞的繁殖速度,在一个更换期内,藻密度波动程度较大,固定组1次/12 h二次更换期间有3~6 h的藻液滤清时间,固定组1次/6 h在第一次产幼前培养液更换前后藻密度变化不大,之后也有较大的下降。相关研究表明[10-18],溞密度和食物密度对枝角类个体的呼吸率、消化率、摄食强度及活动和代谢等一系列生命活动有一定影响,目前尚未有相关证据表明这是溞密度和藻密度诱导蒙古裸腹溞雄体产生的内在机理,但根据本研究结果可以推论,增加溞密度和藻密度波动幅度均有提高蒙古裸腹溞雄体产出率的效应。
[1]王伟洪,王 岩,楼 宝.温度、种群密度和食物密度对蒙古裸腹溞两性生殖雌体形成的影响[J].海洋与湖沼,2006,37(增刊):94-100.
[2]郑 重.淡水枝角类的生殖[J].动物学杂志,1959(1):22-28.
[3]郑 重,曹文清.中国海洋枝角类研究(生殖)[J].海洋学报,1984,6(3):377-388.
[4]成永旭.生物饵料培养学[M].北京:中国农业出版社,2005.
[5]赵云龙.枝角类动物生殖生物学研究进展[J].生物学教学,2012,37(1):2-4.
[6]曹文清,林元烧,郭东晖,等.蒙古裸腹溞染色体组型研究[J].台湾海峡,1999,18(1):71-75.
[7]卢芳野,何志辉.蒙古裸腹溞休眠卵诱发规律的研究[J].大连水产学院学报,1999,14(1):1-8.
[8]曹文清,林元烧,郭东晖.若干生态因子对蒙古裸腹溞生殖方式转变效应的实验[J].台湾海峡,2001,20(增刊):190-195.
[9]王金秋,张佳忆,黄 博.生态因子的波动对多刺裸腹溞雄体发生的影响[J].应用生态学报,2000,11(1):135-137.
[10]BANTA A M,BROWN L A.Control of sex in Cladocera.Ⅲ.Localization of the critical period for control of sex[J].PNAS,1929,15:71-811.
[11]HOBAEK A,LARSSON P.Sex determination in Daphnia magna[J].Ecology,1983,18:37-46.
[12]ANDERS H,PETTER L.Sex determination in Daphnia magna[J].Ecology,1990,71(6):2 255-2 268.
[13]CARVALHO G R,HUGHES R N.The effect of food availability,female culture-density and photoperiod on ephippia productionin Daphnia magna Straus(crustacean:cladocera)[J].Freshwater Biology,1983,13:37-46.
[14]ABU~REZQ T S,YULE A B,TENG S K.Ingestion,fecundity,growth rates and culture of the hqrpacticoid copepod,Tisbe furcata,in the laboratory[J].Hydrobiologia,1997,347:109-118.
[15]BURNS C W.Crowding-induced changes in growth,reproduction and morphology of Daphnia[J].Frshwater Biology,2000,43:19-29.
[16]TESSIER A J,GOULDEN C E.Cladoceran juvenile growth[J].Limno Oceanogr,1987,32(3):680-686.
[17]BONON C A,SAINT-JEAN L.The regulation mechanisms and yield of brackish water populations of Moina micrura reared in tanks[J].Aquaculture,1998,160:69-70.
[18]STROSS R G,HILL J C.Diapause induction in Daphnia requires two stimili[J].Science,1965,150:1 462-1 464.
猜你喜欢
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
蒙古学问题与争论(2021年0期)2022-01-19
蒙古学问题与争论(2021年0期)2022-01-19
中学生物学(2021年8期)2021-11-02
生物学通报(2021年5期)2021-03-16
海洋通报(2020年3期)2020-10-20
水产养殖(2020年9期)2020-09-24
山西农业大学学报(自然科学版)(2020年1期)2020-03-04
阅读与作文(小学高年级版)(2019年8期)2019-10-16
蒙古学问题与争论(2019年0期)2019-03-29
