
Ca2+对CcRIPII-2基因转化宁阳大枣的影响
2014-07-05 16:25:03黄艳艳罗磊牛庆霖刘峰翁曼丽刘静冯殿齐
山东农业大学学报(自然科学版) 2014年4期
黄艳艳,罗磊,牛庆霖,,刘峰,翁曼丽,刘静,冯殿齐*
Ca2+对CcRIPII-2基因转化宁阳大枣的影响
黄艳艳1,罗磊1,牛庆霖2,1,刘峰2,翁曼丽3,刘静1,冯殿齐1*
1.泰安市泰山林业科学研究院,山东泰安271000
2.山东农业大学,山东泰安271018
3.中国科学院遗传与发育生物学研究所,北京100101
本研究应用农杆菌介导的方法,采用L9(34)正交试验设计,对影响宁阳大枣基因转化的各项因子进行了分析比较,并着重探索了Ca2+浓度对侵染转化的影响。结果表明:侵染效率最高的组合为1/2侵染液浓度,20 min侵染时间和0 mg/L Ca2+浓度。Ca2+浓度在提高宁阳大枣基因转化效率方面没有明显促进效果。Ca2+浓度仅对平均分化芽数影响较大,浓度为0 mg/L时,平均分化芽数最多。本试验共获得27株转化苗,试验转化效率为33.33%。
农杆菌介导法;香樟核糖体失活蛋白基因;枣疯病;Ca2+浓度
枣疯病是我国枣树的一种严重病害,由MLO病毒感染引起[1]。防治方法主要有两类:一是常规防治方法,二是化学防治方法,但防治效果都不理想[2]。随着基因工程技术的发展,基因工程已成为现代植物改良育种的有效手段。本试验针对当地特有优良枣树品种宁阳大枣(Ningyang Jujube)深受枣疯病困扰的现实,以自主构建的香樟CcRIP II抗病基因为目的基因,利用该基因产生的对病毒和真菌的广谱抗性,采用农杆菌介导的方法,使用已建立良好再生体系的优良枣树品种宁阳大枣组培苗为试验材料,进行基因转化试验。
农杆菌介导法受多重因素的影响。本试验设立了侵染液浓度,侵染时间与Ca2+浓度3个因素,设计正交试验。试验在以往枣树农杆菌介导试验的基础上,将Ca2+的浓度作为一个试验变量来进行研究,着重探索了Ca2+浓度对农杆菌侵染过程的影响,探索更为高效的农杆菌介导法,以提高当地特有品种宁阳大枣的基因转化效率,并进一步提高宁阳大枣抗枣疯病的能力,也为枣树基因转化研究和防治枣疯病做些有益探索。
1 材料与方法
1.1正交试验设计

表1 L9(34)正交试验设计表Table 1 Orthogonal experiment design L9(34)

表2 农杆菌基因转化L9(34)正交试验设计Table 2 Agrobacterium-mediated gene transformation orthogonal experiment design L9(34)
1.2受体材料
本试验受体材料为宁阳大枣(Ningyang Jujube)长红品种,为泰安市泰山林业科学研究院生物技术中心自主培养的组培苗。
1.3基因来源
本研究所采用的香樟核糖体失活蛋白基因CcRIP II,由中国科学院遗传与发育生物学研究所和泰安市泰山林业科学研究院共同分离克隆。
1.4实验步骤
1.4.1 预培养取宁阳大枣长红组培苗为待用材料,选择生长健壮的茎段作为侵染试材。每个茎段(1 cm左右)至少要保留一对芽,切两端,平铺到培养基上。试验共9个组合,每个组合3瓶。每个组合对应2个对照组,共45瓶。将按试验设计做好的瓶苗作好标记并注明日期,,暗培养2~6 d。
1.4.2 菌种活化将含有香樟核糖体失活蛋白基因(CcRIP II-2)的农杆菌按150µL:100 mL体积比接种于含50 mg/L卡那霉素和50 mg/L利福平的YEP液体培养基中,在摇床上于28℃,180 r/min条件下培养过夜。菌液OD值:0.4~0.6。
1.4.3 侵染将已预培养的材料取出来,在无菌条件下进行适当刻伤处理(对照组同样处理),根据正交设计加入相应的侵染液进行侵染。其中原液即为含有CcRIP II-2基因的正常摇菌所得的农杆菌菌液浓度(菌液OD值0.4~0.6);1/2菌液为含一半CcRIP II-2基因的农杆菌菌液(菌液OD值0.4~0.6)和一半MS母液的混合液体;1/3菌液为含有1/3 CcRIP II-2基因的农杆菌菌液和2/3MS母液的混合液体。在侵染过程中要进行间歇性的摇动。
1.4.4 共培养按试验设计侵染结束,将材料从侵液中取出放到带有无菌滤纸的培养皿上,吸干表面菌液,接种在分化培养基上,共27瓶转化材料和18瓶对照材料,一起暗共培养2 d。
1.4.5 卡那霉素筛选共培养结束,将受体材料和相应的对照材料转接于含有卡那霉素的筛选培养基上继续培养。卡那筛选培养基的选择压力设定2个不同的梯度:30 mg/L(一筛培养基)、50 mg/L(二筛培养基)。选择过程为2个周期,每周期依次设置7 d,30 d。选择培养结束可对初选出的抗性苗进行采样、提取DNA和进行PCR检测。
1.5试验调查
侵染材料转入卡那霉素一筛培养后开始对转化材料进行相关指标的调查。分别调查一筛Kan 30 mg/L,二筛Kan 50 mg/L培养基上苗木生长情况数据。
本试验主要是针对筛选后植物材料的成活率%、每株枯死程度(全部枯死,每株枯死3/5、每株枯死1/5)、平均分化芽数/株、平均新芽高度(cm)等指标进行调查分析。
1.6分子检测
对经过第二次筛选培养30 d的宁阳大枣转化苗进行采样,按试验组合顺序编号。本试验共获得81个存活的抗性芽苗,因有的芽苗过小,采样时只采集到27个株号的组培苗。对这27个号提取DNA并进行PCR扩增检测。根据所提供的CcRIP II-2基因的序列资料,设计了一对引物:
引物1:5'-TCATGGCACCTGATATCACCACCGCT-3'
引物2:5'-TATTAGGCAGCGGATGGGGCGGGAA–3'
2 结果与分析
2.1第一次卡那霉筛选结果
在共培养结束后,将受体材料茎段转移至一筛培养基中(卡那霉素浓度为30 mg/L)培养,7 d后对每个组合中的成活苗、枯死苗、每株枯死1/5与每株枯死3/5的茎段所占的比例以及平均新芽高度、平均分化芽数以及生长状态做了调查统计。见表3。

表3 卡那霉素第一次筛选调查Table 3 The first selection of Kanamycin
根据表3数据,可以直观看出受体材料一筛培养时的成活率和分化生长情况,见图1、图2:
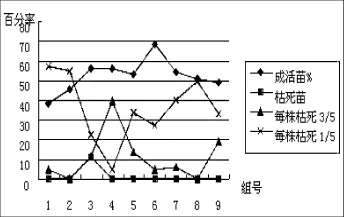
图1 一筛成活率Fig.1 Survival rate of the first screening culture
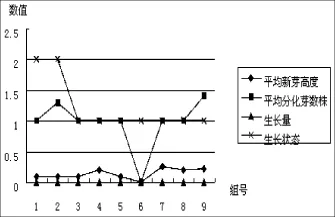
图2 一筛分化生长情况Fig.2 Differential growth of the first screening culture
从图1可直观看出,成活苗最多的是组合6(1/2侵染液浓度+20 min侵染时间+440 mg/L Ca2+浓度),成活率为68.3%,与成活率最低的组合1(侵染液原液+10 min侵染时间+0 mg/L Ca2+浓度)相差30.2%。从受体材料枯死程度看,组合2和组合8枯死枯死程度为3/5的数值最低,不到10%,组合4最高达40%;枯死程度为1/5的组合中,组合1最高,达60%,组合4(1/2侵染液浓度+10 min侵染时间+880 mg/L Ca2+浓度)最低,不到10%。综合各组合受体材料的成活率表现,成活率差异较大,基本无枯死苗;枯死程度为1/5的和枯死程度为3/5的数量呈相反增减趋势,这一点从组合3至组合9的趋势最明显。因此,最有利于提高受体材料成活率的是组合6(1/2侵染液浓度+20 min侵染时间+440 mg/L Ca2+浓度)。
从图2看出,受体材料生长状态逐渐增强,组合1、2长势一般,组合3至组合9长势稳定增强;分化新芽数量差异不大,每组数量都在1~1.5个之间,组合9最多,为1.4个;各组合的新芽高度都很低,都不到0.5 cm,无明显生长,这和培养时间短有关。综合以上分析,一筛培养时,受体材料生长状况还处于新芽萌发和适应生长阶段,较适合新芽萌发的是组合9(1/3侵染液浓度+20 min侵染时间+0 mg/L Ca2+浓度)。
根据表3第一次筛选结果,对受体材料成活率、平均新芽高度、平均分化芽数进行极差分析,结果见表4:

表4 第一次筛选培养极差分析Table 4 orthogonal analysis to the first screening culture
由表4分析得出,对成活率影响最大的因素为侵染液浓度,其次为Ca2+浓度,侵染时间影响最小。在各因素中,侵染液浓度以1/2侵染液浓度为最佳,侵染时间以20 min最佳,Ca2+浓度为440 mg/L时最佳。因此成活率最高的组合是:1/2侵染液浓度+20 min侵染时间+440 mg/L Ca2+浓度。
对平均新芽高度影响最大的因素为侵染液浓度,其次为侵染时间,影响最小的是Ca2+浓度。得到平均新芽高度最高的组合为:1/3侵染液浓度+10 min侵染时间+880 mg/L Ca2+浓度。
对平均分化芽数影响最大的因素为Ca2+浓度,其次为侵染液浓度,影响最小的是侵染时间。得到平均分化芽数最多的组合为:1/3侵染液浓度+10 min侵染时间+0 mg/L Ca2+浓度。
综合一筛培养直观分析和极差分析结果,在成活率和新芽分化数量方面基本一致。
2.2第二次卡那霉筛选
在一筛培养结束后,将受体材料茎段转入二筛培养基(卡那霉素浓度为50 mg/L)继续培养,30 d后调查每个组合中的成活苗、枯死苗、每株枯死1/5与每株枯死3/5的茎段所占的比例以及平均新芽高度、平均分化芽数、生长量以及生长状态,结果见表7。

表5 卡那霉素第二次筛选Table 5 The second screening culture of Kanamycin
根据表5数据,可以直观看出受体材料二筛培养时的成活率和分化生长情况,见图3、图4:
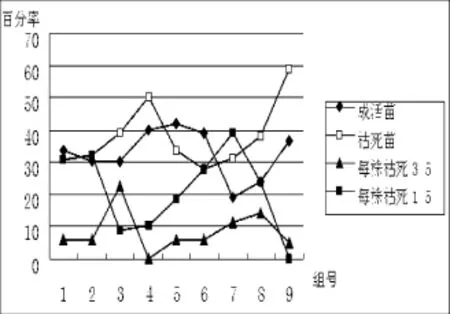
图3 二筛成活率Fig.3 Survival rate of the second screening culture

图4 二筛分化生长情况Fig.4 Differential growth of the second screening culture
从图3看出,在二筛培养中变化最明显的是枯死苗数量显著增加,呈明显上升趋势,组合9枯死率最高,有接近60%的受体材料都枯死了,其次是组合4,枯死率最低的是组合6,不到30%,与最高枯死率相差30%左右;各组合成活苗数量也出现明显差异,成活率最高的是组合5,为42%,最低的是组合7(1/3侵染液浓度+10 min侵染时间+440 mg/L Ca2+浓度),仅19%,两者相差23%;枯死程度3/5和1/5的表现较一致,枯死率呈现减—增—减的数量整体趋向。综上,成活苗最多的是组合5(1/2侵染液浓度+15 min侵染时间+0 mg/L Ca2+浓度),枯死率最高的是组合9(1/2侵染液浓度+20 min侵染时间+440 mg/L Ca2+浓度)。
从图4看出,二筛受体材料生长状态呈现前后一般中间较强的特点,组合1、2、3、8、9为同一水平,长势一般,组合4、5、6、7为同一水平,长势较强,无明显突出的组合;各组合分化芽数差异不大,都在1个~1.5个之间,其中组合2最多,为1.5个;各组合生长量差异不大,都在0.5 cm~1.5 cm之间,组合2最大,为1.4 cm,比最小的组合8多0.68 cm;新芽高度差异不显著,最大的组合7为1.17 cm比最小的组合5高0.65 cm。综上,除生长状态外,各组合的新芽高度、分化新芽数量和生长量生长趋势基本一致,差异幅度都不大。组合2(侵染原液+15 min侵染时间+440 mg/L Ca2+浓度)分化芽数最多,生长量最大,组合7(1/3侵染液浓度+10 min侵染时间+440 mg/L Ca2+浓度)新芽最高。
根据表5第二次筛选所获数据进行正交分析,结果见表6。
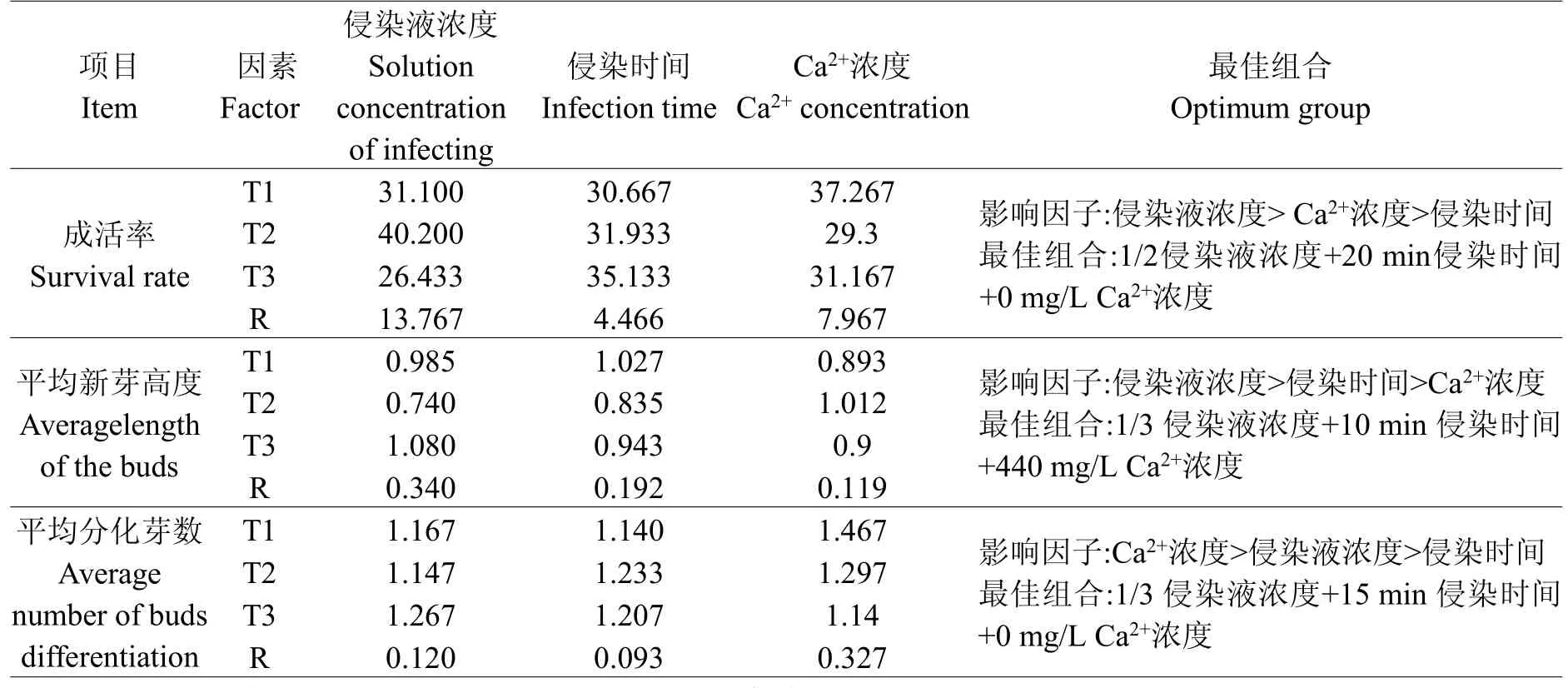
表6 第二次筛选培养极差分析Table 6 Range analysis to the second screening culture
从表6计算的受体材料成活率、平均新芽高度、平均分化芽数极差结果,得出:
各因素对成活率的影响依次为:侵染液浓度>Ca2+浓度>侵染时间。得到成活率最高的组合为:1/2侵染液浓度+20 min侵染时间+0 mg/L Ca2+浓度。
各因素对平均新芽高度的影响依次为:侵染液浓度>侵染时间>Ca2+浓度。得到平均新芽高度最高的组合为:1/3侵染液浓度+10 min侵染时间+440 mg/L Ca2+浓度。
各因素对平均分化芽数的影响依次为:Ca2+浓度>侵染液浓度>侵染时间。得到平均分化芽数最多的组合:1/3侵染液浓度+15 min侵染时间+0 mg/L Ca2+浓度。
综合二筛极差分析结果,侵染液浓度对成活率和新芽高度影响最大,Ca2+浓度对分化芽数影响最大。
综合二筛培养直观分析和极差分析结果:
影响受体材料成活率和新芽高度最大的因素是侵染液浓度,影响分化芽数最大的因素是Ca2+浓度。
成活率最高的是组合5:1/2侵染液浓度+15 min侵染时间+0 mg/L Ca2+浓度;
枯死率最高的是组合9:1/2侵染液浓度+20 min侵染时间+440 mg/L Ca2+浓度;
分化芽数最多,生长量最大的是组合2:侵染原液+15 min侵染时间+440 mg/L Ca2+浓度;新芽生长最高的是组合7:1/3侵染液浓度+10 min侵染时间+440 mg/L Ca2+浓度。
综合比较一筛和二筛极差分析结果,见表7:

表7 极差分析比较Table 7 Comparison of the range analysis
从表7看出,在一筛培养时,对受体材料成活率和新芽高度影响最大的是侵染液浓度,对分化芽数影响最大的是Ca2+浓度;二筛培养时,对受体材料成活率和新芽分化数影响最大的是Ca2+浓度,侵染液浓度对新芽高度影响较大。综上,两次筛选培养中,Ca2+浓度对新芽分化数影响都最大,成活率受侵染液浓度和Ca2+浓度双重影响,侵染液浓度都是影响新芽高度的最重要因素。
对一筛、二筛的最佳组合进行比较,分析两次培养的差异,见表8。

表8 最佳组合比较Table 8Aggregate optimum group
综合分析两次筛选培养最佳组合,结果基本类似,只略有不同。
Ca2+浓度对两次筛选培养中受体材料成活率和新芽高度有明显影响,在侵染液浓度和侵染时间相同情况下,一筛培养的Ca2+浓度比二筛培养的浓度高440 mg/L。在两次培养中,适合提高成活率的侵染液浓度都是1/2侵染液浓度,侵染时间都是20 min;适合新芽高生长的侵染液浓度都是1/3侵染液浓度,侵染时间都是10 min;
Ca2+浓度对新芽分化数影响不明显,两次筛选培养中都是不添加Ca2+时分化芽数最多。侵染时间对两次筛选的影响差异最大,在侵染液浓度和Ca2+浓度相同情况下,一筛培养选用10 min侵染时间,二筛培养选用15 min侵染时间能分化最多新芽。
本试验中,根据第二次筛选培养结果,可初步判断受体材料基因转化效率,主要以筛选后转化受体的成活率为标准,所以转化效率最高的组合为:1/2侵染液浓度+20 min侵染时间+0 mg/LCa2+浓度。
2.3转基因植株PCR分子鉴定

图5 PCR产物电泳图谱Fig.5 PCR electrophoretogram
由PCR电泳图可见,检测的27个株号转基因植株全部显示出明显的阳性目的条带,而未转化植株没有显示出目的条带。这初步表明外源香樟核糖体失活蛋白基因已整合到宁阳大枣基因组内。
3 讨论
本试验采用的核糖体失活蛋白基因RIP是一种专一性较强的毒蛋白,可抑制某些蛋白质的生物合成,从而达到提高抗性的目的。RIP的抗性基因工程已在烟草、小麦、玉米、番茄等很多植物中稳定表达,且对相应的病原菌具有一定程度的抗性[3]。因此转RIP基因工程正发展成为防治植物病虫害的新途径[4]。本试验所采用的CcRIP II-2基因包含了RIP基因的这一特性,通过对宁阳大枣进行农杆菌介导试验,获得27个抗性单株,经PCR检测,初步确定CcRIP II-2基因已经整合到宁阳大枣基因组中。这些抗性单株的获得,为宁阳大枣抗枣疯病品种选育和果树抗性育种研究开辟了一条新途径。
有研究证明,农杆菌介导的植物基因转化效率受多个因子的影响,包括农杆菌的株型、质粒的构建、外植体的再生频率、筛选标记的可靠性,以及转化过程中外部环境的控制等,其中前几个因子在转化方案确定后不会有很大的改变,而转化的外部环境可以根据实验操作的需要作出较大的调整,这些调整在提高转化效率上往往收益显著[5]。因此,我们把Ca2+作为转化过程中的一个外部影响因素加以考虑。Ca2+信号通路是植物中信号转导已确认的主要途径,它参与了多种生理活动的调节[6]。大量研究表明,Ca2+也参与了植物----微生物互作的信号传递[7]。我们推测在农杆菌侵染过程中Ca2+也发挥了重要的作用。
通过试验分析,发现Ca2+在农杆菌介导法转化CcRIP II-2基因过程中对受体材料转化效率确有明显影响但对促进转化效率提高作用不明显,未添加Ca2+的侵染液较添加了不同浓度Ca2+的获得了较高的转化率。尽管Ca2+对提高宁阳大枣基因转化效率促进作用不明显,但其对宁阳大枣(Ningyang Jujube)受体材料成活率和新芽高度却有显著促进作用,这说明,Ca2+作为迄今为止唯一被证实的植物细胞内信号,在植物的生长发育及其对环境的反应和适应中起着十分重要的作用[8]。
另外,Ca2+浓度对新芽分化数也有影响,侵染液中不添加Ca2+时宁阳大枣受体材料分化芽数比添加了Ca2+的要多,说明侵染液中的Ca2+对于受体材料芽组织的分化生长促进作用不明显。
本试验中,在筛选培养基中添加不同浓度的卡那霉素,同时为了抑制农杆菌的过度生长对植物受体造成伤害,还添加了抑菌素,这些都对转化受体材料具有一定毒害作用,也在一定程度上影响了植物材料的正常生长。张和臣等认为,植物对细胞外部的逆境信号需要细胞感受并跨越细胞膜才能把信息输进细胞,从而诱导相应基因的表达,产生各种生物反应,引起细胞膜渗透压的增加,进而引起细胞壁间的机械胁迫以及膜的流动性等。植物通过膜的变化诱发植物产生钙信号并诱发产生由膜外到膜内的信号传递过程[9],因此,受体材料的状态和生长环境都对试验结果有影响。
因本试验样本较小,对Ca2+浓度的设计梯度有所限制,故Ca2+浓度水平对枣树基因转化的确切影响还有待进一步试验研究,对获得的转基因植株的稳定性表现也还需进一步检测。
4 结论
(1)Ca2+对宁阳大枣基因转化效率有显著影响,不添加Ca2+可获得较高的转化率。经PCR检测,本试验共获得27个转化植株,转化率为33.33%。由此看出,Ca2+对提高宁阳大枣转化香樟核糖体失活蛋白基因CcRIP II-2转化效率促进作用不明显;
(2)Ca2+浓度对宁阳大枣受体材料成活率和新芽高度有明显影响,在侵染液浓度和侵染时间相同情况下,添加Ca2+范围在0 mg/L~440 mg/L对转化受体成活率影响相同,添加Ca2+范围在440 mg/L~880 mg/L对转化受体新芽高度影响相同;成活率受侵染液浓度和Ca2+浓度双重影响,侵染液浓度是影响新芽高度的最重要因素;
(3)Ca2+浓度对新芽分化数有显著影响,侵染液中不添加Ca2+时宁阳大枣受体材料分化芽数最多;
(4)影响受体材料成活率和新芽高度最大的因素是侵染液浓度,影响分化芽数最大的因素是Ca2+浓度;(5)添加Ca2+的宁阳大枣(Ningyang Jujube)抗MLO病毒基因转化试验最佳组合为:效率最高的组合:1/2侵染液浓度+20 min侵染时间+0 mg/LCa2+浓度;成活率最高的组合:1/2侵染液浓度+15 min侵染时间+0 mg/L Ca2+浓度;枯死率最高的组合:1/2侵染液浓度+20 min侵染时间+440 mg/L Ca2+浓度;分化芽数最多,生长量最大的组合:侵染原液+15 min侵染时间+440 mg/L Ca2+浓度;新芽生长最高的是组合7:1/3侵染液浓度+10 min侵染时间+440 mg/L Ca2+浓度。
[1]王祈楷,徐绍华,陈子文,等.枣疯病的研究[J].植物病理学报,1981,11(1):15-18
[2]程丽芬,毛静琴,梁凤玉,等.枣疯病的发病规律及防治[J].山西林业科技,1995(3):36-37
[3]单丽波,贾旭.核糖体失活蛋白及其在植物抗真菌病基因工程中的应用[J].生物工程进展,2000,20(6):74-78
[4]李建国.核糖体失活蛋白研究进展[J].分子植物育种,2005,3(4):566-570
[5]郑进,康薇,洪华珠.影响农杆菌基因转化效率因素的研究[J].黄石理工学院学报,2008,24(5):30-33
[6]张君诚,孟玉环,宋育红,等.植物Ca2+-CaM信号系统及其调控研究进展[J].重庆师范大学学报(自然科学版),2005,22(4):55-58
[7]杨民和,王国红.Ca2+对植物—微生物互作反应的调控[J].江西农业大学学报,2001(2):43-48
[8]简令成,王红.Ca~(2+)在植物细胞对逆境反应和适应中的调节作用[J].植物学通报,2008,(3):255-267
[9]张和臣,尹伟伦,夏新莉,等.非生物逆境胁迫下植物钙信号转导的分子机制[J].植物学通报,2007,24(1):114-122
Effect of Ca2+on CcRIP II-2 Gene Transformation of Ningyang Jujube
HUANG Yan-yan1,LUO Lei1,NIU Qing-lin2,1,LIU Feng2,WENG Man-li3, LIU Jing1,FENG Dian-qi1*
1.Taishan Institute of Forestry Science,Tai'an271000,China
2.Shandong Agricultural University,Tai'an271018,China
3.Institute of Genetics and Developmental Biology,Chinese Academy of Sciences,Beijing100101,China
In this study,the concentration effect of Ca2+on transformation ofRIPgene(Ribosome Inactivating Protein Gene) cloned fromCinnamomum camphoratoNingyang Jujube was investigated in detail with L9(34)orthogonal design,through the analysis of factors which affected the transformation.The results showed that the concentration of Ca2+had no crucial effect on transformation and the best combination was the bacterium solution with 0.4-0.6 OD value was double diluted,20-minutes infection and without Ca2+.27 transformation seedlings were obtained and the transformation rate was 33.33%.
Agrobacterium-mediated transformation;Cinnamomum camphoraL.Ribosome-inactivating protein(RIP); Jujube Witches Broom;Ca2+concentration
S722.3文献标示码:A
1000-2324(2014)04-0481-08
2012-10-18
2013-03-12
抗枣疯病植原体(MLO)基因转化项目
黄艳艳(1978-),女,工程师,主要从事林果花卉生物技术研究.E-mail:Yanhuang0321@163.com
*通讯作者:Author for correspondence.E-mail:fengdianqi@163.com
猜你喜欢
当代作家(2024年3期)2024-06-29 22:18:20
中国民间疗法(2021年7期)2021-07-22 06:44:12
中等数学(2020年3期)2020-08-24 07:59:18
红豆(2018年4期)2018-04-20 02:28:52
红豆(2018年4期)2018-04-20 02:28:52
海峡姐妹(2018年1期)2018-04-12 06:44:30
丝路艺术(2018年7期)2018-04-01 22:05:29
阅读与作文(小学高年级版)(2016年3期)2016-03-08 21:12:23
云南中医学院学报(2015年3期)2015-07-31 18:09:27
中医研究(2014年12期)2014-03-11 20:29:56
