
H9N2亚型禽流感重组病毒的拯救
2014-06-17 02:38肖月蕊颜瑞萍李明义
动物医学进展 2014年10期
肖月蕊,颜瑞萍,李明义,高 轩*
(1.河北农业大学动物科技学院,河北保定071000;2.诸城市畜牧兽医管理局,山东诸城262200;3.山东信得科技股份有限公司,山东青岛266101)
禽流感病毒(Avian influenza virus,AIV)为正 黏病毒科A 型流感病毒,属于单股负链RNA 病毒。禽流感病毒的基因组分为HA、NA、NP、PA、PB1、PB2、M 和NS8个片段。其中PB1、PB2和PA 3个蛋白组成病毒的RNA 聚合酶,NP和M1称为病毒的内源蛋白,HA、NA 和M2 为外源蛋白,NS1 和NS2为非结构蛋白[1]。AIV 变异频繁,血清亚型众多,根据流感病毒表面结构蛋白血凝素(HA)和神经氨酸酶(NA)抗原性的差异可分为不同的亚型[2]。在众多亚型中,H9N2亚型禽流感病毒虽然为中低致病力毒株,但对养禽业的危害不容忽视,1996年—2000年,H9N2亚型禽流感的发病率占总禽流感发病的93.89%,成为影响我国养禽业的主要AIV 亚型[3-4]。
1994年的A/Chicken/Beijing/1/94(H9N2)分离株曾给我国养禽业造成极大损失,唐秀英等[5]也先后从发病禽中分离到大量的H9N2亚型禽流感病毒,其公共卫生意义引起了全世界的广泛关注,对该亚型病毒的研究也取得了较大的进展,其中应用比较广泛的技术为反向遗传操作技术。虽然传统疫苗在保护畜禽免受禽流感感染的过程中起到了不可替代的作用,但传统疫苗受制作工艺的限制,其在保存、使用和接种后的副作用等方面都会影响免疫效力。因此有必要研制新型疫苗来弥补其不足之处[6]。流感病毒的反向遗传操作系统可以在分子水平上研究流感病毒的生活周期,了解流感病毒的致病机制和生物学特性,并且可以研制安全有效的新型流感疫苗[7]。反向遗传操作技术最具潜力、最具吸引力的应用还在于流感疫苗株的筛选。一方面因为其疫苗株筛选的时间短,可以解决流感大流行发生时疫苗生产的时效问题,为流感流行期间流感疫苗的制备争取时间;另一方面可以构建在动物细胞传代中高度增殖的疫苗株,而不是常用的利用鸡胚进行病毒增殖的方法,这在大大降低生产成本的同时也避免了病毒增殖后鸡胚残体的焚烧,降低了对环境污染[8]。12或17质粒系统[9]的建立,开创了该系统的新篇章,与以前的一些方法相比比较简便,不再需要辅助病毒,避免了繁重的筛选工作。但其最大的缺点就是质粒比较多,工作量大,最后拯救的结果往往不是很理想,Hoffmann E等[10]创建的8质粒拯救系统为流感病毒疫苗候选株的快速筛选提供了可能,为人类在反向遗传系统的研究开启了新的篇章。
本研究以H1N1 亚型禽流感病毒的6 个基因片段(PB2、PB1、PA、M、NP和NS)和H9N2亚型禽流感病毒的2 个基因片段(NA 和HA)为基因供体,构建与H9N2亚型流感病毒具有相同基因组合方式的重组病毒。并研究了该重组病毒在293T 和MDCK 细胞上的生长特性,对其安全性、稳定性进行了初步评价,为H9 亚型禽流感疫苗的研制开辟了新的途径。
1 材料与方法
1.1 材料
1.1.1 毒株与质粒 H9N2亚型禽流感病毒山东地区分离株,由本实验室(山东信得科技股份有限公司青岛研发中心)分离、鉴定和保存。已构建好的含有H1N1流感病毒6个基因片段的pHW2000载体(pHW2000-PB2、pHW2000-PB1、pHW2000-PA、pHW2000-M、pHW2000-NP 和pHW2000-NS),用于cDNA 克隆的双向转录/表达空载体pHW2000,由本实验室保存。
1.1.2 细胞 DH 5α大肠埃希菌感受态细胞购自北京博迈德科技股份有限公司。人胚肾T 细胞(293T)、非洲绿猴肾细胞(Vero细胞),由本实验室保存。
1.1.3 实验动物 10dSPF 鸡胚,为山东省家禽研究所产品。
1.1.4 主要试剂 T4DNA 连接酶、RNasin(RNA酶抑制剂)、pMD18-T vector、DNA Marker 2 000、低分子量标准蛋白Marker、IPTG,宝生物工程(大连)有限公司产品;限制性内切酶StuⅠ (10 U/μL)、dNTPs、L-LMV 反转录酶,Fermentas公司产品;RNA 提取试剂TRIzols、转染试剂LipofectamineTM 2000、OPTI-MEM、Expand High FidelityTaq酶,无内毒素质粒提取纯化试剂盒,Invitrogen公司产品;Agrose Gel DNA 回收试剂盒,OMEGA 公司产品;TPCK 处理的胰酶,Sigma公司产品;细胞培养液DMEM、胎牛血清,GIBICO 公司产品;BsmBⅠ(10 U/μL)和BsaⅠ(10 U/μL),NEB公司产品。10mL/L 鸡红细胞按常规方法自行制备。
1.1.5 试验用溶液及其配制 氨苄青霉素、卡那霉素、庆大霉素、LB液体培养基、LB固体培养基、SOC培养基、X-gal、IPTG、50×TA、生理盐水,均由本实验室配制。
1.2 方法
1.2.1 引物的设计与合成 引物设计参考Hoffman E等[11]报道,设计扩增流感病毒HA 和NA 全基因引物,引物序列如下:Bm-HA-1:5′-TATTCGTCTCAGGGAGCAAAAGCAGGGG-3′;Bm-HA-2:5′-ATATCGTCTCGTATTAGTAGAAACAAGGGTGTTTT-3′; Ba-NA-1:5′-TATTGGTCTCAGGGAGCAAAAGCAGGAGT-3′; Ba-NA-2:5′-ATATGGTCTCGTATTAGTAGAAACAAGGAGTTTTTT-3′。
引物序列中有BsmBⅠ和BsaⅠ酶切位点,反转录通用引物为Unit12。引物由上海生工生物工程技术服务有限公司合成。
1.2.2 病毒RNA 提取和cDNA 的合成 用OMEGA 公司总RNA 提取试剂盒从含有H9N2亚型禽流感病毒的尿囊液中提取病毒RNA,按Hoffmann E等[12]的方法进行RT-PCR反应。取病毒RNA 悬液29μL,与2μL反转录引物混匀后置于65 ℃水浴5min,取出后冰浴5min,依次加入5×RT buffer 10 μL,10mmoL/L dNTPs 5μL,RNasin(40U/μL)1.5 μL,反转录酶AMV(10U/μL)1μL,42 ℃反应60 min。将HA 基因和NA 基因反转录后的cDNA产物作为PCR反应的模板。
1.2.3 HA 和NA 基因的扩增 以1.2.2中所得产物为模板扩增各基因片段,PCR反应体系为:10×Expand FidelityTaqbuffer 2.5μL,10mmoL/L dNTPs 1μL,上游引物(20μmol/L)1μL,下游引物(20μmol/L)1μL,cDNA 2μL,Expand High FidelityTaq0.5μL,最后加入ddH2O 补至25μL。PCR反应程序:95 ℃5min;94 ℃45s,55 ℃~58℃50s,72 ℃1min~2.5min,共进行30个循环;72 ℃10min。取PCR 产物,进行10g/L琼脂糖凝胶电泳鉴定。
1.2.4 转录/表达载体构建 胶回收1.2.3中HA和NA 的PCR 产物,按照pMD18-T 载体试剂盒说明书与T 载体连接并进行蓝白斑筛选,对克隆的阳性质粒命名为pMD18-T-HA 和pMD18-T-NA。分别对其进行BsmBⅠ和BsaⅠ酶切鉴定并测序。
鉴定产物用限制性内切酶BsaⅠ和BsmBⅠ进行酶切后回收,与BsmBⅠ酶切过pHW2000载体进行连接,转化入大肠埃希菌DH 5α感受态细胞中,加入5mL 培养基振荡摇菌。取出100μL 菌液涂布于含有氨苄青霉素(50μg/mL)的LB 平板,放入37 ℃培养箱培养16h,挑取白色孤立菌落,接种于5mL含有氨苄青霉素(50μg/mL)液体LB培养基,放入37℃培养箱培养12h,提取各个基因片段重组质粒进行鉴定,每个片段的阳性质粒送3个~5个克隆测序。测序正确的质粒命名为pHW-HA 和pHW-NA。
1.2.5 转录/表达载体阳性质粒活性测定 测序鉴定正确的阳性克隆一部分进行扩大培养后提取质粒,为下一步的转染做准备,一部分保存于-80 ℃超低温冰箱。取2μL质粒200倍稀释后,紫外分光光度计测定质粒的浓度和纯度,含量在100μg/mL以上、OD 260nm/OD 280nm 为1.75~2.00的质粒备转染用。
1.2.6 重组病毒的拯救 将293T 细胞和Vero细胞按2∶1的细胞量铺入6孔板,待细胞长至70%~90%融合度时用无血清无抗生素的OPTI-MEM培养基洗2 次后进行转染。将含H1N1 亚型流感病毒6个基因的阳性质粒(pHW2000-PB2、pHW2000-PB1、 pHW2000-PA、 pHW2000-M、pHW2000-NP和pHW2000-NS)与含H9N2 亚型流感病毒分离株HA、NA 基因的质粒pHW-HA 和pHW-NA 按质量比1∶1 的比例混合均匀,与转染试剂一起加入到无血清、无抗生素的DMEM 中,按说明书进行分散和结合后均匀覆盖于细胞上,在37 ℃、体积分数为5% 的CO2培养箱中吸附6h,换入含100mL/L 血清的DMEM 1.0mL,再培养72h,观察病变,吹落细胞后与上清一起收集,以除HA 之外的七质粒共转为阴性对照。
1.2.7 重组病毒的鉴定 收集转染细胞和上清,经尿囊腔接种于10dSPF鸡胚,每胚0.2mL,每个样品接种3个胚,置37 ℃孵化箱培养72h,观察鸡胚死亡情况,分别收集尿囊液,用OIE 推荐的标准进行血凝(HA)试验,HA 阳性者用H9亚型流感病毒的阳性血清进行血凝抑制(HI)试验。第1 代阴性的取尿囊液再盲传1代。
HI阳性尿囊液,取其鸡胚传代的第4代尿囊液提取RNA,用RT-PCR方法(同1.2.2和1.2.3)扩增获救病毒NA 和HA 基因,送至上海生工生物工程技术服务有限公司进行测序;未经反转录的RNA 用通用引物进行PCR 扩增对照,以排除尿囊液中转染质粒DNA 的存在。阴性对照的尿囊液同样进行验证。
将含获救H9N2 亚型流感病毒的第4 代鸡胚尿囊液进行10倍递次稀释,0.2mL/胚接种于10d SPF鸡胚,每个稀释梯度接种4个鸡胚,37 ℃培养,弃掉24h之内的死胚,24h之后死亡的鸡胚置4℃保存。通过测定感染鸡胚尿囊液的血凝活性来判断其是否感染,用Reed Muench法计算EID50。
2 结果
2.1 目的基因的获得
以病毒cDNA 为模板,经PCR 扩增后,取1μL PCR 产物,进行10g/L琼脂糖凝胶电泳鉴定,结果表明成功获得目的基因(图1)。
2.2 转录/表达载体的构建
2.2.1 目的基因与18-T 载体的连接 经过蓝白斑筛选挑取白色克隆,经鉴定后阳性克隆扩增摇菌12h左右,提取质粒酶切后进行PCR鉴定重组载体,结果如图2。将上述阳性的菌液送上海生工生物工程技术服务有限公司进行测序,测序结果与相应基因序列完全一致。
2.2.2 目的基因与pHW2000载体的连接转化将HA、NA 两个目的片段与载体经过连接转化后,挑单克隆,用菌落PCR 的方法鉴定阳性克隆,琼脂糖凝胶电泳结果见图3。将通过菌落PCR鉴定为阳性的菌液送上海生工生物工程技术服务有限公司进行测序,并通过DNA Star软件进行序列比对,结果H9N2亚型流感重组病毒片段序列正确无误,均符合病毒拯救要求,转染重组质粒构建成功。
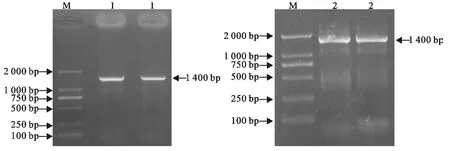
图1 禽流感H9N2亚型山东分离株HA、NA 基因的RT-PCR扩增结果Fig.1 RT-PCR Products of HA and NA genes of AIV H9N2Shandong strain
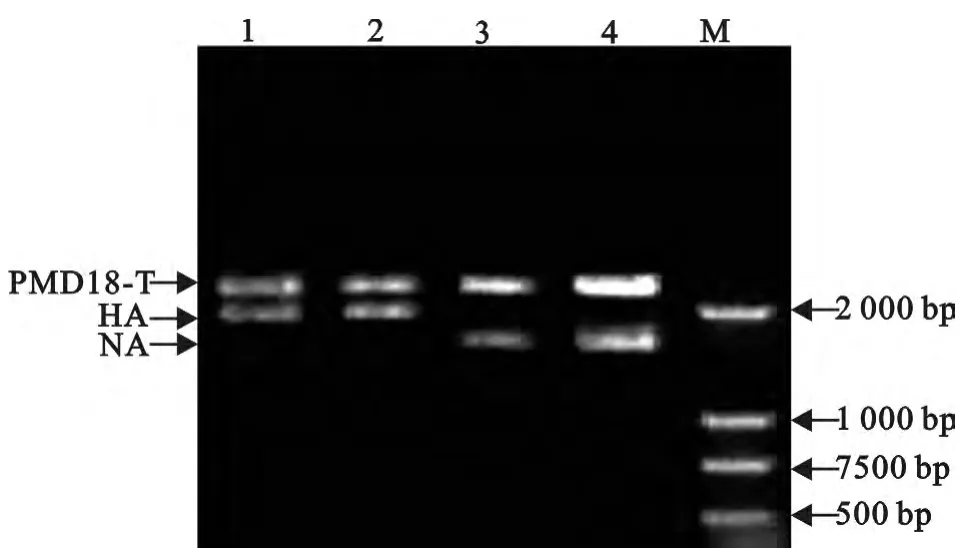
图2 HA 基因和NA 基因连接T 载体扩增产物的酶切结果Fig.2 The enzyme digestion results of amplification products of HA and NA genes connected with T vector gene
图3 HA 和NA 基因PCR鉴定结果Fig.3 RT-PCR products of HA and NA genes
2.2.3 转录/表达载体阳性质粒浓度和纯度测定测序鉴定正确的阳性克隆进行扩大培养后提取质粒,取2μL质粒200倍稀释后,紫外分光光度计测定质粒的浓度和纯度,测定结果见表1。
2.3 重组病毒的拯救
8个重组质粒共转染293T/Vero细胞48h后,收集病毒和细胞,接种第1代10dSPF 鸡胚,72h后收集尿囊液测定血凝效价达29。用H9亚型阳性血清进行重组病毒的血凝抑制试验,结果表明,病毒可被H9阳性血清抑制,血凝抑制效价为29。阴性对照转染孔的上清液和293T/Vero细胞接种鸡胚,连续传2代,鸡胚尿囊液中未检测到血凝效价。
表1 紫外分光光度计检测结果Table 1 The detection results of ultraviolet spectrophotometer
2.4 获救H9N2亚型流感病毒的RT-PCR及测序鉴定
HI阳性尿囊液,取其鸡胚传代的第4代尿囊液提取RNA,用RT-PCR 方法(同1.2.2和1.2.3)扩增获救H9N2病毒NA 和HA 基因,送至上海生工生物工程技术服务有限公司进行测序;鉴定结果表明,获救病毒的HA 和NA 基因与H9N2山东分离株完全一致。
2.5 获救病毒鸡胚半数感染剂量(EID50)的测定
参照文献的方法,将传至第4代的H9N2亚型重组病毒尿囊液接种于鸡胚,观察鸡胚死亡情况,Reed-Muench法可得获救H9N2 亚型流感病毒鸡胚半数感染剂量(EID50)为10-7.75/0.2mL(表2)。
表2 EID50测定鸡胚感染和死亡数Table 2 Infection(I)and death(D)numbers of chicken embryos in EID50test
3 讨论
病毒拯救技术是在了解病毒复制特点等基础上,利用分子生物学技术建立和完善起来的,是通过人工操作基因,用病毒核酸的适当形式在一定条件下转染细胞,产生有感染性的病毒粒子[13]。流感病毒的反向遗传操作技术虽然已经广泛应用于探讨流感病毒基因结构与功能、揭示病毒的致病、传播和免疫机理、构建疫苗候选株等方面的研究,但全病毒拯救还是比较困难,拯救成功的大多是那些高致病性毒株[14]。
在本试验建立的8质粒拯救系统中,所用双向转录/表达载体pHW2000构建和克隆虽然步骤较为繁琐,但具有比单向载体更高的效率,每个片段连接的效率高达80%以上。用转染48h后的上清接种鸡胚,在第1代均可上升至高效价,经4次传代,和分离株一样,病毒增殖能稳定在一定滴度范围之内。同时通过改变质粒浓度来控制起始病毒蛋白的量,从而到达提高转染效率的目的,以保证转染成功。本试验中经过多次测定,同时结合文献报道,采用的质粒浓度为200μg/mL,纯度(260/280)为1.75~2.0。另外,在转染过程中发现,细胞长满80%~90%时,共转效果最好。本试验以H1N1亚型流感病毒的6 个内部基因片段为骨架与H9N2 亚型流感病毒的HA 和NA 基因进行重组,成功拯救出了与H9N2亚型禽流感病毒具有同样基因组合方式的重组病毒,经测定获得的重组H9N2亚型流感病毒的血凝效价为29,高于亲本株(27)。这可能是因为重组病毒中含有鸡胚高适应性的H1N1 亚型流感病毒的基因,其作为骨架拯救出的重组病毒可以达到很高的血凝效价,这保证了足够的抗原量并降低了成本,重组病毒在鸡胚上连续传4代,其增殖能与亲本株一样稳定在一想的滴度范围之内。具有较高的免疫效果并可以在细胞内稳定传代生长,使其可以作为新型疫苗候选株使用,为禽流感的其他基础性研究及疫苗的研制奠定了基础。病毒拯救是否成功,关键在于基因的准确克隆,本试验在进行H9亚型禽流感病毒NA 基因的克隆过程中,非编码区的序列突变导致多次克隆失败,经过多次的筛选测定才得到准确序列,使得下一步的病毒拯救可以顺利进行。本试验还对获救H9N2 亚型流感病毒进行了传代,传至第4代的获救H9N2亚型流感病毒的HA 和NA 基因序列仍与分离株一致,说明获救的病毒稳定,保持了其抗原特性。
流感病毒亚型众多,变异很快,特别2013 年H7N9亚型禽流感病毒的发现,说明流感病毒已经突破了种间屏障[15]。疫苗作为防控流感流行的有效手段,得到了大家的公认,但是传统的疫苗生产工艺需要的时间长且步骤繁琐。反向遗传学的发展一方面简化了流感疫苗株产生的流程,缩短了疫苗制备所需的时间,减少了疫苗生产的成本,另一方面,反向遗传学技术的发展也为流感病毒的基础研究提供了便利[16]。因此,我们有理由相信,反向遗传学在流感病毒这个领域中将会得到越来越多的应用。
[1]沈 佳,章振华,姜北宇,等.2株H9N2亚型禽流感病毒全基因组序列测定及遗传性分析[J].安徽农业科学,2011(9):5241-5244.
[2]Ha Y,Steven D J,John J S,et al.H5avian and H9swine influenza virus haemagglutinin structures:possible origin of influenza subtypes[J].EMBO J,2002,21:865-875.
[3]Chen B L,Zhang Z J,Chen W B,et al.Isolation and preliminary serological characterization of type A influenza viruses from chickens[J].Chin J Vet Med(Chinese),1944,22:3-5.
[4]Liu H Q,Liu X F,Cheng J,et al.Phylogenetic analysis of the hemagglutinin genes of 26avian influenza viruses of subtype H9N2isolated from chickens in China during 1996-2001[J].Avian Dis,2003,47:116-127.
[5]唐秀英,付朝阳,冯菊艳.H9亚型禽流感的流行与防制//中国畜牧兽医学会禽病学分会.中国畜牧兽医学会禽病学会分会第十次学术研讨会论文集[C].中国畜牧兽医学会禽病学分会,2000:4.
[6]刘志刚,刘立峰,陈伦勇,等.禽流感新型疫苗研究进展[J].动物医学进展,2013,34(2):99-103.
[7]严悌昆,王延树,向 华.RNA 病毒反向遗传的应用前景[J].广东畜牧兽医科技,2007(6):3-4,8.
[8]刘大飞,刘 明,刘春国,等.H1N1亚型猪流感病毒反向遗传操作系统的建立及高产细胞型疫苗株的拯救[J].中国农业科学,2009(5):1797-1804.
[9]Neumann G,Kawaoka Y.Reverse genetics of influenza virus[J].Virology,2001,287:243-250.
[10]Hoffmann E,Neumann G,Kawaoka Y,et al.A DNA t ransfection system for generation of influenza A virus from eight plasmids[J].Proc Natl Acad Sci,2000,97:6108-6113.
[11]Hoffmann E,Stech J,Guan Y,et al.Universal primer set for the full-length amplification of all influenza A viruses[J].Arch Virol,2001,146(12):2275-2289.
[12]Hoffmann E,Krauss S,Perez D,et al.Eight-plasmid system for rapid generation of influenza virus vaccines[J].Vaccine,2002,20(25-26):3165-3170.
[13]叶柱德,胡江锋.H9N2亚型禽流感病毒疫苗研究进展[J].动物医学进展,2008,29(2):89-92.
[14]靳 冬,吴志明,张志凌,等.反向遗传学操作技术在禽流感研究中的应用[J].中国动物检疫,2011(3):75-77.
[15]Zhuang Q Y,Wang S C,Wu M L,et al.Epidemiological and risk analysis of the H7N9subtype influenza outbreak in China at its early stage[J].Chin Sci Bulle,2013,26:7-8.
[16]张鹏飞,杜 山,李云章.反向遗传操作技术在流感病毒研究中的应用[J].畜牧与饲料科学,2008(2):69-72.
猜你喜欢
广东医科大学学报(2020年6期)2020-02-06
启蒙(3-7岁)(2017年4期)2017-06-15
中国介入影像与治疗学(2017年12期)2017-01-14
现代检验医学杂志(2016年3期)2016-11-15
现代检验医学杂志(2016年1期)2016-11-12
华南农业大学学报(2015年5期)2015-12-04
现代泌尿外科杂志(2015年9期)2015-02-20
当代畜禽养殖业(2014年7期)2014-02-27
当代畜禽养殖业(2014年6期)2014-02-27
当代畜禽养殖业(2014年4期)2014-02-27
