
温州潮间带大型底栖动物资源调查与分析
2014-06-13 07:54王凤丽韩志强胡成业水柏年
浙江海洋大学学报(自然科学版) 2014年2期
王凤丽,韩志强,徐 衡,杜 肖,郑 伟,胡成业,李 良,水柏年
(1.浙江海洋学院水产学院,浙江舟山 316022;2.乐清市水产科学研究所,浙江乐清 325600)
潮间带是湿地生态系统的重要类型,素有地球之肾之称,而潮间带大型底栖动物又是海洋生物资源的重要组成部分,其资源及生物多样性状况既关系到海洋生态系统平衡,也关系海洋生物产业的健康持续发展。关于潮间带大型底栖动物资源及生物多样性,国内外有一些研究及其报道。例如,在国外,BAKRI等[1-2]对科威特潮间带大型底栖动物群落与环境因子关系的研究;DAVIDSON[3]探讨了物种迁移对滩涂大型底栖动物分布、多样性、密度和生物量在小尺度上的影响机制;OTANI等[4]对日本濑户内海滩涂区大型底栖动物群落结构与生境的关系进行了研究等。在国内,梁超愉等[5]研究了大亚湾潮间带生物种类组成、数量分布及生物多样性;安传光等[6]对崇明岛潮间带夏季大型底栖动物多样性的研究;黄雅琴等[7]研究了湄洲湾潮间带底栖生物多样性;鲍毅新等[8]对温州灵昆岛潮间带大型底栖动物及其季节性变化的研究;彭欣等[9]对南麂列岛底栖动物群落时空变化的研究;赵永强等[10]对椒江口潮间带大型底栖动物的生态位研究等。有关温州潮间带大型底栖动物的研究,主要有吕永林等[11]对浙江洞头大竹屿岛潮间带大型底栖生物多样性的研究;仇建标等[12]对洞头列岛潮间带大型底栖生物的时空分布及其多样性的研究;郑荣泉等[13]研究了乐清湾潮间带大型底栖动物群落结构分布格局及其对人类活动的响应;高爱根等[14]对鳌江口南侧苍南县泥相潮间带春末底栖动物生态特征的研究;高爱根等[15]对南麂列岛的潮间带贝类资源时空分布的研究等。但研究基本针对局部海域,对温州潮间带大型底栖动物的生物多样性及群落稳定性的全面系统的研究尚未见报道。本文试图利用2012年秋季的调查资料,结合已有资料分析温州潮间带大型底栖动物资源,揭示其现状及存在问题,提出生态修复与保护建议。
1 材料与方法
1.1 材料
2012年秋季在温州潮间带6条断面调查采样所得的大型底栖动物样本,以及查询所得的有关调查研究资料。
1.2 方法
按照《海洋调查规范第6部分:海洋生物调查》(GB12763.6-2007)于2012年10月进行潮间带大型底栖动物采样与实验。
1.2.1 采样方法
在乐清、平阳和苍南滩涂区各设一个调查断面,另在苍南霞关岩礁区设一条调查断面,洞头岩礁区设迎风与背风两个调查断面,每个断面布设三站,即高潮、中潮与低潮带,每站随机取4~6个样方,采样断面设置如图1。采样时用GPS定位,样方为50 cm×50 cm,滩涂区软相底质采样时先取框内表面的大型底栖动物,再挖取样框内底泥至硬底,用孔径1.0 mm的筛网冲洗去泥沙等。岩礁区采样时,先将框内的易碎生物计数,再采集样框内所有大型底栖动物。获得的样品用无水乙醇固定保存。
1.2.2 实验方法
在实验室,利用有关文献资料对所采集的样本进行鉴定、分类并测定。所用天平为HENGPING MP1100B型,精度0.01 g。
1.2.3 数据处理
生物多样性指数及优势度分析采用以下公式计算:

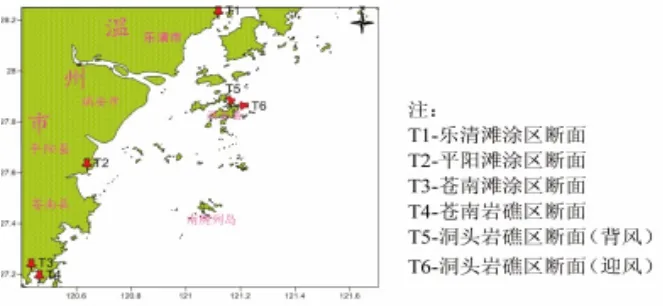
图1 温州潮间带大型底栖动物调查断面Fig.1 Sampling sections for the intertidal macrobenthos in Wenzhou

式中,S:样品中的物种总数;N:群落中所有物种的总丰度;Pi:第i种的丰度(ni)与总丰度(N)之比值;N%:某一物种尾数占总尾数的百分比;W%:某一物种重量占总重量的百分比;F%:某一物种在调查中出现的样方数占总样方数的百分比(即出现频率)。
IRI包含了大型底栖生物的个体数、生物量和出现频率这3个重要信息,能充分反映它们在群落中所处的地位,文中规定IRI值大于1000的为优势种,100~1000之间的为重要种,10~100为常见种,1~10为一般种,小于1为稀有种。
用PRIMER5.0软件绘制ABC曲线,评价潮间带大型底栖动物群落结构受干扰程度。ABC曲线对于任何物理性、生物性以及污染扰动引起的大型底栖生物群落变化都能灵敏地作出反应,在稳定的环境中,生物量曲线位于丰度曲线上方;当群落受到中度干扰时,两条曲线相互交叉或重叠在一起;严重干扰时,丰度曲线位于生物量曲线之上[17-19]。
2 结果与分析
2.1 种类组成与数量分布
2.1.1 种类组成
2012年秋季,共采集到潮间带大型底栖动物16目31科45种,其中,甲壳类最多,有18种,占总种数的40.00%,隶属于3目10科;其次为软体动物,有13种,占28.89%,隶属于13目13科;环节动物3种,占6.67%;纽形动物和星虫动物各1种,均占2.22%;棘皮动物2种,占4.44%;鱼类7种,占15.56%。其中,在滩涂区采到甲壳类13种、软体动物3种,而在岩礁区甲壳类6种、软体动物9种。各断面大型底栖动物的分布特征见表1。

表1 温州潮间带大型底栖动物种类组成与分布Tab.1 Species composition and distribution of the intertidal macrobenthos in Wenzhou

续表1
2.1.2 数量分布
滩涂区生物量T1断面最高,为40.84 g/m2;T3次之,为23.54 g/m2;T2最低,仅为10.12 g/m2。丰度T2最高,为89.33 ind/m2;T1 次之,为 48.27 ind/m2;T3 最低,为 26.40 ind/m2。生物量垂直分布T1断面中潮区最高,低潮区最低;T2中潮区最高,高潮区最低;T3低潮区最高,中潮区最低。丰度垂直分布T1断面高潮区最高,低潮区最低;T2、T3均为中潮区最高,高潮区最低(图 2)。
岩礁区生物量T4断面最高,为361.16 g/m2;T6次之,为110.71 g/m2;T5 最低,为 58.90 g/m2。丰度 T4 最高,为 242.93 ind/m2;T6 次之,为 148.80 ind/m2;T5 最低,为 105.20 ind/m2。生物量垂直分布T4断面低潮区最高,高潮区最低;T5、T6高潮区高于中潮区;丰度垂直分布T4断面低潮区最高,高潮区最低;T5、T6高潮区高于中潮区(其中T5、T6断面由于风浪太大,低潮区未采样)(图2)。

图2 温州潮间带各断面大型底栖动物生物量与丰度分布Fig.2 Distribution of biomass and abundance of the intertidal macrobenthos in each section in Wenzhou

图3 温州潮间带各断面大型底栖动物分类群生物量与丰度分布Fig.3 Distribution of biomass and abundance of different intertidal macrobenthos groups in each section in Wenzhou
由图3可知,3个滩涂区断面均以甲壳类为主,生物量和丰度都占各自断面50%以上;软体动物在T1和T2所占比例高于T3;星虫动物在T3占7.21%;其它4个类群所占比例都较小。岩礁区只采到软体动物、甲壳类和少量环节动物,各断面以软体动物为主;T4断面甲壳类生物量占50%以上,而软体动物丰度占50%以上;T5、T6两个断面软体动物生物量和丰度均占65%以上。各断面分类群生物量与丰度水平分布详见表2。

表2 温州潮间带各断面大型底栖动物分类群生物量与丰度分布Tab.2 DistributionofbiomassandabundanceofdifferentintertidalmacrobenthosgroupsineachsectioninWenzhou
2.2 优势种及相对重要性指数
根据IRI值优势种、重要种和稀有种判定标准可知,在滩涂区,T1断面优势种为长足长方蟹、珠带拟蟹守螺和日本大眼蟹3种,重要种为弓形革囊星虫、缢蛏等4种;T2断面优势种为台湾泥蟹、长足长方蟹和微黄镰玉螺3种,重要种为日本大眼蟹等3种;T3断面优势种为长足长方蟹、日本大眼蟹和弓形革囊星虫3种,重要种为鲜明鼓虾。在岩礁区,T4断面优势种为鳞笠藤壶和疣荔枝螺2种,重要种为青蚶;T5断面优势种为单齿螺、平背蜞和疣荔枝螺3种,重要种为嫁和青蚶2种;T6断面疣荔枝螺占了绝对的优势。各断面均未发现稀有种。各断面优势种与重要种及其IRI值见表3。

表3 温州潮间带各断面大型底栖动物优势种与重要种Tab.3 Dominant and important species of the intertidal macrobenthos in each section in Wenzhou
2.3 生物多样性指数
滩涂区生物多样性水平最高的是T3断面,多样性指数(H′)、均匀度指数(J)、丰度指数(D)都最大,依次为 2.98、0.70、2.72,C 值最小,为 0.22;T2 生物多样性水平最差,C 值最大,为 0.26,H′、J、D 都最小,依次为2.27、0.63、1.31。岩礁区生物多样性水平最高的是 T5 断面,H′、J、D 都最大,依次为 2.12、0.64、1.12,C 值最小,为 0.30;T6 生物多样性水平最差,C 值最大,为 0.93,H′、J、D 都最小,依次为 0.32、0.12、0.59。滩涂区各断面生物多样性水平普遍高于岩礁区断面,且T3生物多样性水平最高,T6生物多样性水平最低(表4)。

表4 温州潮间带各断面大型底栖动物生物多样性指数Tab.4 The biodiversity index of the intertidal macrobenthos in each section in Wenzhou
2.4 ABC曲线分析
由图4分析可知,T1断面生物量曲线与丰度曲线有两次相交,两条曲线相距很近,表明群落结构受到中等程度的干扰;T2丰度曲线大部分位于生物量曲线之上,表明群落结构受到的干扰较严重;T3两条曲线有一次相交,但生物量曲线大部分位于丰度曲线之上,表明群落结构受到的干扰较轻;T4两条曲线有两次相交,且相距颇近,几近重叠,表明群落结构受到中等程度的干扰;T5生物量曲线位于丰度曲线之上,但两条曲线相距较近,表明群落结构受到的干扰较小;T6丰度曲线位于生物量曲线之上,表明群落结构受到了较严重的干扰。
2.5 邻近海域历史研究结果
彭欣等[13]在2006年秋季和2007年春季对乐清湾潮间带滩涂区进行了大型底栖动物调查,其中与T1断面相距最近的南塘断面调查结果:秋季和春季生物量依次为38.82 g/m2、91.97 g/m2,丰度依次为271 ind/m2、1476 ind·/m2;秋季主要优势种为短拟沼螺,春季为焦河蓝蛤;秋季多样性指数(H′)、丰度指数(D)、均匀度指数(J)依次为 2.34、5.89、0.46,春季依次为 2.81、4.19、0.58。
国家海洋局第二海洋研究所于2010年春季和2011年秋季对鳌江口潮间带进行了大型底栖动物调查,平阳县邻近的两个滩涂区断面调查结果为:春、秋季生物量平均值依次为17.95 g/m2、41.15 g/m2,丰度平均值依次为232 ind/m2、112 ind/m2,软体动物生物量和丰度均最高,其次为甲壳类;春季多样性指数(H′)、丰度指数(D)、均匀度指数(J)均值依次为 2.27、0.73、0.76,秋季依次为 1.92、1.17、0.89。
国家海洋局第二海洋研究所于2011年春季和秋季对苍南县大渔湾潮间带进行了大型底栖动物调查,春、秋季的底栖动物生物量依次为 70.52/m2、41.60/m2,丰度依次为 121.5 ind/m2、75.2 ind/m2,春、秋两季生物量与丰度都是软体动物最高,生物量和丰度的垂直分布都是中潮区最高,高潮区最低;春季多样性指数(H′)、均匀度指数(J)依次为 1.70、0.71,秋季依次为 0.84、0.84。
仇建标等[12]在2006年秋季和2007年春季对洞头列岛潮间带(岩礁相)开展了大型底栖动物调查,洞头本岛断面调查结果为:秋季、春季大型底栖动物生物量依次为1058.87 g/m2、2105.17 g/m2,丰度依次为474 ind/m2、1154 ind/m2,生物量和丰度都是中潮区最高,高潮区最低;秋季主要优势种为日本笠藤壶、条纹隔贻贝,春季为日本笠藤壶、隔贻贝;秋季多样性指数(H′)、丰度指数(D)、均匀度指数(J)依次为1.90、2.05、0.68,春季依次为 1.53、2.95、0.47。

图4 温州潮间带各断面大型底栖动物ABC曲线Fig.4 ABC curves of intertidal macrobenthos in each section in Wenzhou
3 讨论
3.1 种类组成与数量分布
在滩涂区采到甲壳类13种、软体动物3种,其它几个类群也采到了样本,但种类数较少;在岩礁区采到甲壳类6种、软体动物9种,还有少量的环节动物,其它类群均未采到;相似底质的断面其种类组成也有一定的差异。这可能是潮间带生物的生活空间既受到潮汐、波浪作用的限制,又受到阳光、气温和盐度等的影响[24-25]所致。
岩礁区断面的生物量与丰度均远高于滩涂区断面,这可能与不同底质生境中的生物种类存在差异有关,岩石底质适合营固着生活的种类分布[26,27],在小范围的空间内聚集了大量的大型底栖动物,导致单位面积内生物量和丰度达到相当高的水平,这与赵永强的研究结果[28]一致。
各断面秋季的生物量与丰度较邻近海域同季结果有较大幅度的下降。T1断面与乐清湾南塘断面相比,生物量相当,但丰度低了将近5倍;T2断面与平阳两断面平均值相比,生物量低3倍,丰度低0.25倍;T3断面与苍南大渔湾平均值相比,生物量低将近1倍,丰度低将近2倍;而洞头两个断面T5、T6与洞头本岛相比,生物量平均值低了11倍,丰度低2倍多。这可能与近几年来温州海域环境污染严重,滩涂围垦工程的相继实施以及潮间带大型底栖动物资源遭到滥采乱挖,资源受到严重的破坏而快速衰退有关。各断面生物量与丰度普遍较低,也可能与采样季节有关。邻近海域的研究资料(2006年秋季、2007年春季,乐清湾;2010年春季、2011年秋季,平阳;2011年春、秋季,苍南;2006年秋季、2007年春季,洞头)表明,生物量与丰度有较明显的季节变化,且秋季往往明显低于春季[12,13],但总体的季节变化情况还有待更进一步的研究证实。
3.2 优势种分析
岩礁区T6断面疣荔枝螺(IRI=18 760)占了绝对的优势,T4、T5优势种除疣荔枝螺(IRI值依次为7 717和1 865)外还有鳞笠藤壶、平背蜞、单齿螺(其IRI>4 500),岩礁区优势种组成有差异可能是受海岸开敞度的影响,研究表明从隐蔽海岸到开敞海岸占优势的软体动物逐渐被甲壳动物所取代[29,30],而T6断面与T4、T5断面相比海岸开敞度较小。同时,本研究得出,滩涂区大型底栖动物分布受海岸开敞度影响与岩礁区并不一致,T3断面较另外两个滩涂区断面(T1、T2)开敞度小,但从生物量与丰度以及IRI值来看,软体动物并不占优势。相反,T1、T2断面软体动物(红树拟蟹守螺、微黄镰玉螺的IRI>150)也是优势种。这可能是生活在滩涂区的大型底栖动物主要营埋栖生活,海岸开敞度对其影响较小所致。
T5、T6同为洞头岩礁区断面,但优势种组成有一定的差异,即T6断面疣荔枝螺占绝对优势(IRI=18 760),而T5断面优势种除疣荔枝螺(IRI=1 865)外还有单齿螺(IRI=9 476)和平背蜞(IRI=4 795),这除了与海岸开敞度有关外,还可能与风浪大小有关,T5断面位于背风面,风浪相对较小,适于生存的种类相对较多,而T6断面位于迎风面,风浪较大,海浪冲刷较强,其生境难以适于一些种类分布。仇建标等对洞头列岛的研究[12]中作为洞头本岛秋季主要优势种的条纹隔贻贝在T5、T6断面并不是优势种,这可能是由其生活习性决定的(条纹隔贻贝宜附着生活在中、低潮区的岩石等物体上),但也不能排除人为因素影响,条纹隔贻贝是重要的经济贝类,可能遭到人为的滥采乱挖更严重,导致其数量显著减少。
3.3 生物多样性与群落结构稳定性分析
根据生物多样性与ABC曲线结果可知,滩涂区T3断面生物多样性水平较高,群落组成较丰富,群落稳定性较好;而T2断面生物多样性水平较低,群落组成较单一,群落稳定性较差;尽管T1断面生物多样性不是最差的,但与彭欣等的研究结果[13]相比,其生物多样性水平有较大幅度的降低。这可能与海域污染程度有关,由2007-2012年温州市海洋环境质量公报可知,T3断面所在海域属于较清洁海域,T1与T2断面所在海域均属于严重污染海域,这也进一步佐证了污染程度对大型底栖动物的生物多样性与群落结构的影响。T2断面生物多样性水平较低、群落稳定性差,也可能是由于滩涂围垦工程等海洋工程改变了潮间带生物的生境状况,使群落结构单一化程度越来越高,稳定性降低[9,28]。岩礁区T5断面生物多样性水平较高,群落稳定性较好;T6断面生物多样性水平较低,物种组成较单一,群落稳定性差,可能是由于T5断面位于背风面,风浪较小,其生境适于生存的种类较多;而T6断面处于风浪较大的迎风面,适于分布的种类较少。
4 结语
根据本次调查研究结果并结合历史资料分析可知,温州潮间带大型底栖动物生物量与丰度都较低,且呈下降趋势,生物多样性水平较低,群落组成较单一,物种分布不均匀,群落结构受到了不可忽视的干扰,稳定性较差,资源现状不容乐观。其原因多种多样,如生境污染、过度采捕、滩涂围垦工程的相继实施等,而潮间带作为陆海交互作用的系统单元,其稳定性不高,易受人类活动的影响。建议合理开发利用海洋生物资源,禁止对大型底栖动物的过度采捕;积极开展沿岸生态修复与保护工作,在适宜海域培育红树林,在严重污染的海域放养适宜的藻类以及滤食性贝类,净化海域环境;申请设立更多国家级与省级自然保护区;科学合理地规划滩涂围垦等海洋工程,尽量减少对当地环境的不良影响,以期实现海洋经济的可持续发展。
[1]BAKRI A D,BEHBEHANI M,KHURAIBET A.Quantitative assessment of the intertidal environment of Kuwait I:Integrated environment classification[J].Journal of Environmental Mannagement,1997,51:321-332.
[2]BAKRI A D,BEHBEHANI M,KHURAIBET A.Quantitative assessment of the intertidal environment of Kuwait II:Controlling factors[J].Journal of Environmental Mannagement,1997,51:333-341.
[3]DAVIDSON I C,CROOK A C,BARNES D K A.Quantifying spatial patterns of intertidal biodiversity:is movement important[J].Marine Ecology,2004,25(1):15-34.
[4]OTANI S,KOZUKI Y,MURAKAMI H,et al.Relationship between macrobenthos and physical habitat characters in tidal flat in eastern Seto Inland Sea,Japan[J].Marine Pollution Bulletin,2008,57:142-148.
[5]梁超愉,张汉华,吴进峰.大亚湾潮间带生物种类组成、数量分布及生物多样性研究[J].南方水产,2005,1(3):42-48.
[6]安传光,赵云龙,林 凌,等.崇明岛潮间带夏季大型底栖动物多样性[J].生态学报,2008,28(2):577-586.
[7]黄雅琴,李荣冠,王建军,等.湄洲湾潮间带底栖生物多样性[J].生物多样性,2010,18(2):161-167.
[8]鲍毅新,葛宝明,郑 祥,等.温州湾灵昆岛东滩潮间带大型底栖动物群落的季节动态[J].水生生物学报,2007,31(3):437-444.
[9]彭 欣,谢起浪,陈少波,等.南麂列岛潮间带底栖生物时空分布及其对人类活动的响应[J].海洋与湖沼,2009,40(5):585-589.
[10]赵永强,曾江宁,黄逸君,等.椒江口潮间带大型底栖动物的生态位[J].应用生态学报,2009,30(5):1 176-1 183.
[11]吕永林,张永普,李 凯,等.浙江洞头大竹屿岛潮间带大型底栖生物多样性[J].生态学杂志,2011,30(4):707-716.
[12]仇建标,彭 欣,谢起浪,等.洞头列岛潮间带大型底栖生物的时空分布及其多样性[J].水产学报,2012,36(4):608-613.
[13]彭 欣,谢起浪,陈少波,等.乐清湾潮间带大型底栖动物群落分布格局及其对人类活动的响应[J].生态学报,2011,31(4):954-963.
[14]高爱根,陈全震,杨俊毅,等.浙江苍南泥相潮间带春末底栖动物生态特征[J].海洋科学,2006,30(5):92-96.
[15]高爱根,曾江宁,陈全震,等.南麂列岛海洋自然保护区潮间带贝类资源时空分布[J].海洋学报,2007,29(2):105-111.
[16]RORERT E U.Information theory in ecology[J].Computers&Chemistry,2001,25(4):393-399.
[17]SHANNON C E,WEAVER W.The mathematical theory of communication[M].Urbana:University of Illinois Press,1948.
[18]PIELOU E C.Mathematical Ecology[M].New York:Wiley,1977.
[19]徐兆礼.长江口北支水域浮游动物的研究[J].应用生态学报,2005,16(7):1 341-1 345.
[20]PINKAS L,OLIPHANT M S,IVEROSON I L K.Food habits of albacore,bluefin tuna,and bonito in California waters[J].Fishery Bulletin,1971,152:1-105.
[21]WARWICK R M.A new method for detecting pollution effects on marine macrobenthic communities[J].Marine Biology,1986,92:557-562.
[22]魏永杰,张海波,王晓波,等.象山港潮间带大型底栖动物群落受干扰状况研究[J].海洋环境科学,2009,28(增刊1):增46-增49.
[23]顾晓英,陶 磊,尤仲杰,等.象山港大型底栖动物群落特征[J].海洋与湖沼,2010,41(2):208-213.
[24]郑成兴,李荣冠,江锦祥,等.泉州湾岩相潮间带底栖生物生态研究Ⅰ.物种多样性与分布特征[J].生物多样性,2004,12(6):594-610.
[25]杨俊毅,高爱根,陈全震,等.拟建宁海电厂附近潮间带底栖生物群落生态[J].东海海洋,2004,22(3):48-55.
[26]张永普,应雪萍,吴海龙,等.北麂列岛岩相潮间带底栖生物群落的组成特征[J].海洋湖沼通报,2000(4):26-33.
[27]李宝泉,于海燕,徐凤山,等.胶州湾底栖软体动物与环境因子的关系[J].海洋与湖沼,2005,36(3):194-197.
[28]赵永强,曾江宁,陈全震,等.宁波大榭开发区北岸潮间带春季大型底栖动物群落格局[J].动物学杂志,2009,44(2):78-83.
[29]杨万喜,陈永寿.嵊泗列岛潮间带群落生态学研究Ⅰ.岩相潮间带底栖生物群落组成及季节变化[J].应用生态学报,1996,7(3):305-309.
[30]高爱根,杨俊毅,曾江宁,等.玉环坎门排污口邻近岩相潮间带生物分布特征[J].东海海洋,2004,22(4):24-30.
猜你喜欢
儿童故事画报·自然探秘(2022年3期)2022-04-27
中国水产(2021年1期)2021-03-01
海洋信息技术与应用(2020年4期)2021-01-18
青年文学家(2020年28期)2020-11-02
海洋通报(2020年3期)2020-10-20
科学导报(2020年61期)2020-09-29
少年文艺(2019年5期)2019-05-28
领导决策信息(2018年32期)2018-10-25
消费导刊(2018年10期)2018-08-20
安徽农业科学(2018年21期)2018-05-14
