
云南腾冲县新分离乙型脑炎和盖塔病毒的分子生物学特征
2014-04-09 03:18李胜国张海林付士红康显虎寸待启章域震杨卫红王环宇梁国栋
中国人兽共患病学报 2014年4期
冯 云,李胜国,张海林,付士红,康显虎,寸待启,郭 超,章域震,杨卫红,王环宇,梁国栋
云南西部腾冲县自古以来就是中国与缅甸的重要通商口岸,边境贸易及人员往来频繁。近几年来,中国在“史迪威公路”基础上建造了“腾-密公路”(中国腾冲至缅甸密支那),该公路的建成使得中缅边境贸易得到迅速发展,也增加了各种传染病传播的风险。因此,我们对腾冲县蚊虫中新分离的20株流行性乙型脑炎病毒(Janpanese encephalitis virus, JEV)和1株盖塔病毒(Getah virus, GETV)进行研究。
1 材料和方法
1.1蚊虫采集和病毒分离 在腾冲县人房和畜圈用玻璃吸蚊管和诱蚊灯(“功夫小帅”牌,光催化诱蚊器,武汉市吉星环保科技有限责任公司产品)采集蚊虫标本。诱蚊灯每天采集时间段为19:00至次日7:00共12 h。吸蚊管人工捕蚊法每天采集时间段为19:00至22:30。捕获蚊虫经分类鉴定,分批放入2 mL冻存管,液氮保存运输回实验室,蚊虫标本经研磨后,取上清液接种C6/36和BHK-21细胞进行病毒分离,连续3代出现规律细胞病变判为阳性。
1.2病毒鉴定 阳性分离物上清液用QIAamp Viral RNA Mini Kit(美国QIAGEN公司)提取病毒RNA,Amersham Bioscience Ready-to-GoTM You First-Strand Beads(美国Amersham Pharmacia Biotech公司)制备cDNA。取2 μL cDNA做反应模板。用JEV的PreM、E基因和GETV的E2、Capsid基因特异性引物(表1)[1-2],TaKaRa LA Taq 试剂盒完成RT-PCR扩增。反应结束后,取2 μL产物进行1%琼脂糖凝胶电泳检测扩增效果。
所有测序在北京博迈德科技发展有限公司完成。使用DNAstar软件包中的SeqMan对序列片段进行拼接、编辑、校正。Clustal X(1.8)、MEGA3.1和DNAStar软件包中的MegAlign及GeneDoc等生物学软件进行系统进化分析和核苷酸、氨基酸序列分析。
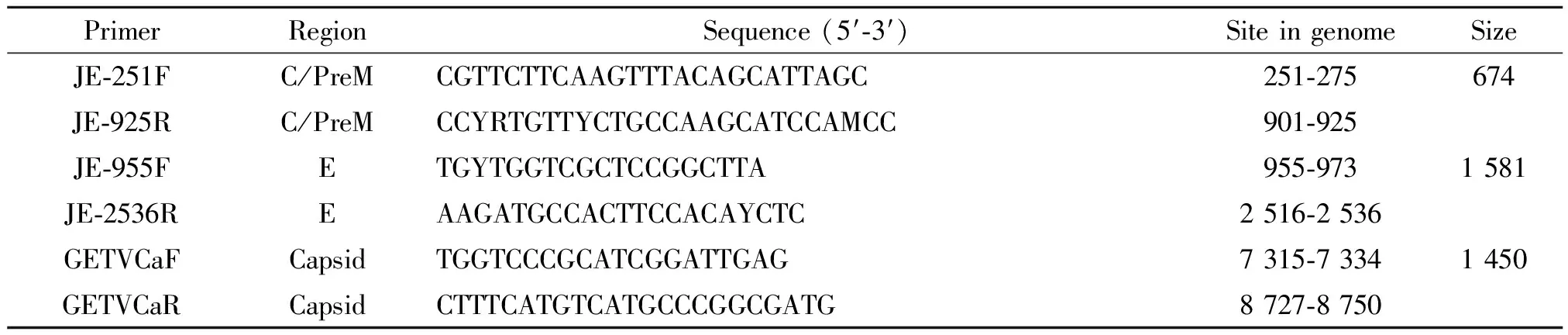
表1 本研究中病毒鉴定使用的引物信息
Note: F means forward primer; R means reverse primer. Y, C/T; M, C/A.
2 结 果
2.1病毒分离 在腾冲县共捕获蚊虫4属25种26 702只,分为310批进行研磨。其中有21批蚊虫标本能引起BHK-21细胞产生规律病变。经JEV PreM、E基因和GETV E2、Capsid基因特异性引物用RT-PCR方法对阳性分离物进行扩增,结果20株为JEV阳性,其中15株分离自三带喙库蚊(Culextritaeniorhynchus),3株分离自中华按蚊(Anophelessinensis),2株分离自骚扰阿蚊(Armigeressubalbatus);另1株为GETV阳性,分离自伪杂鳞库蚊(Culexpseudovishnui)(表2)。
2.2JEV系统进化分析 新分离的20株JEV与来源于GenBank中不同国家、不同年代的4个基因型的56株乙脑病毒PreM基因的240个核苷酸序列构建PreM基因核苷酸进化树提示,其中14株(YNTC07008、YNTC07012、YNTC07020、YNTC07028、YNTC07046、YNTC07099、YNTC07109、YNTC07172、YNTC07255、YNTC07257、YNTC07259、YNTC07273、YNTC07290、YNTC07295)与2004年四川省分离株SC04-25、河南省分离株HN04-21的亲缘关系最近;6株(YNTC07011、YNTC07018、YNTC07101、YNTC07111、YNTC07177、YNTC07292)与2001年上海的分离株SH-53处于同一进化簇中,亲缘关系更接近(图1)。新分离的20株JEV与既往5个基因型别的58株乙脑病毒E基因的1500个核苷酸序列进行系统进化分析,进化树提示15株(YNTC07008、YNTC07012、YNTC07020、YNTC07028、YNTC07046、YNTC07099、YNTC07101、YNTC07109、YNTC07172、YNTC07255、YNTC07257、YNTC07259、YNTC07273、YNTC07290、YNTC07295)与2004年四川省分离株SC04-25处于同一进化簇中;5株(YNTC07011、YNTC07018、YNTC07111、YNTC07177、YNTC07292)与2001年上海的分离株SH-53处于同一进化簇中(图2)。新分离JEV与日本、韩国和越南的JEV在E基因的进化关系上较近,但与泰国分离株的进化关系稍远(图2)。经过对新分离的20株JEV的PreM和E基因的系统树分析表明,本次分离的20株JEV都属于基因Ⅰ型,与我国其它地区近年来分离到的Ⅰ型JEV的进化关系很近,20株新分离的JEV又分别处于基因Ⅰ型乙脑病毒的两个次级分支中,说明它们之间还是存在一定的差异。

表2 蚊虫标本阳性分离物的背景资料
2.3JEV核苷酸同源性和氨基酸位点分析 新分离20株JEV 的PreM基因核苷酸差异度为0.0%~9.2%,其中YNTC07028株与其它分离株的差异度最大,表明当地存在不同株系的JEV流行株。这20株JEV与减毒活疫苗(SA14-14-2株)在PreM区段的共同差异位点位于第64位氨基酸,由T→A。此外,YNTC07011、YNTC07018、YNTC07101、YNTC07111、YNTC07177、YNTC07292株与SA14-14-2株在PreM区段的共同差异位点位于第2位氨基酸,由I→T;YNTC07046株与SA14-14-2株在PreM区段的差异位点位于第45位氨基酸,由A→T;YNTC07028株与SA14-14-2株在PreM区段的差异位点有10个,分别位于第3、4、7、44、45、47、55、57、64、71位氨基酸上。
新分离的20株JEV之间的E基因核苷酸同源性为96.5%~100%,氨基酸的同源性为97.2%~100%,存在一定差异;这20株JEV与SA14-14-2株的核苷酸同源性为87.5%~88.3%,氨基酸同源性为96.4%~97.2%。E蛋白是决定JEV毒力的重要组成[3],其中有8个位点非常重要[4],20株新分离株在这8个位点的差异与其它基因Ⅰ型JEV一致,而E138是影响JEV毒力最重要的位点,如果该位点由谷氨酸(E)替换成赖氨酸(K),则病毒的神经毒力就会明显降低[5]。通过测定新分离株的E138位点都是谷氨酸,说明毒力较疫苗株强。通过对比本研究得到的结果从分子生物学的角度证实了减毒活疫苗SA-14-14-2株能够有效保护当地新分离的JEV流行株的感染。
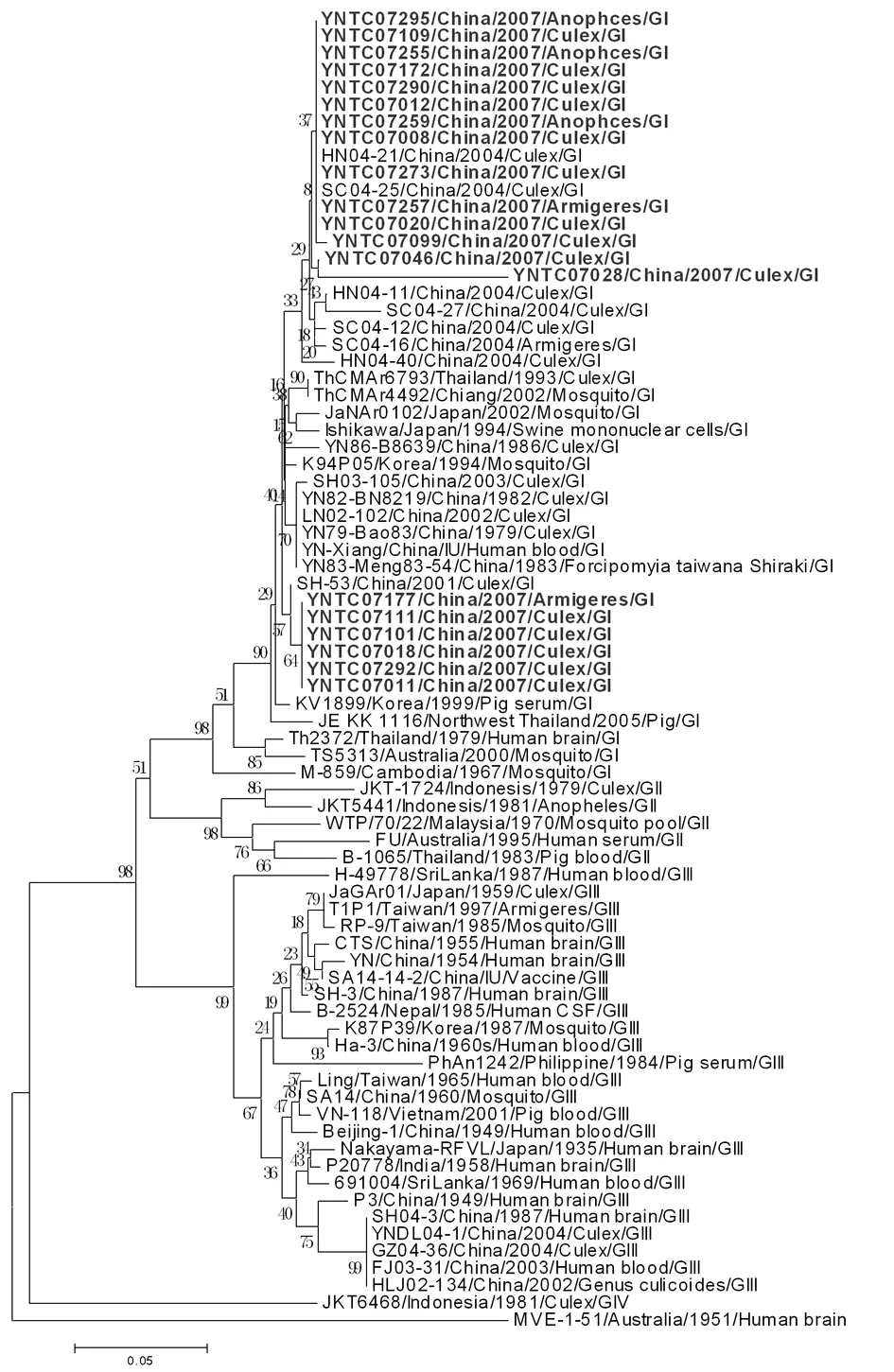
图1以PreM基因为基础的系统进化分析
Fig.1PhylogeneticanalysisofJEVisolatesbasedonPreMgenesequence
2.4GETV系统进化分析 用新分离的YNTC07180与GenBank中其它地区和年代的11株GETV的Capsid基因核苷酸序列构建系统进化树, YNTC07180株与以往云南分离株YN0540位于同一进化分支,亲缘关系最近(图3),而1955年分离自马来西亚的M2021株位于GETV进化分支的根部,与YNTC07180株的亲缘性较远。
2.5GETV核苷酸同源性和氨基酸位点分析 YNTC07180株的Capsid基因与以往云南分离株YN0540的同源性最高,核苷酸和氨基酸序列同源性分别为99.2%和99.3%,表明具有明显地域特征。YNTC07180株与1955年分离自马来西亚的M2021株的核苷酸序列同源性最低。依据GETV代表株(M1)的Capsid基因全长为801nt,编码长267个氨基酸的结构衣壳蛋白进行氨基酸位点分析,将YNTC07180株与来自日本(Sagiyama virus, SAGV)、马来西亚(M2021株)、韩国(GETV SouthKorea)、俄罗斯(LEIV/16275/Mag)、蒙古(LEIV/17741/MPR)以及中国河北分离株(HB0215-3、HB0234)、云南分离株(YN0540)、海南分离株(Alphavirus M1)等的Capsid基因序列进行比对,Capsid基因编码的267个氨基酸中,YNTC07180与M1有2个氨基酸差异位点,第211位(Q→R)和第231位(L→F)。GETV的E2基因有1 266个核苷酸,编码422个氨基酸的病毒包膜糖蛋白,E2蛋白是GETV与宿主细胞受体相互作用的最重要的表面蛋白,也是病毒诱导中和抗体产生的主要结构蛋白。新分离株与其它的17株GETV的E2基因的核苷酸同源性在94.8%~99.9%之间,氨基酸同源性在97.6%~100%之间。YNTC07180分离株与M1的氨基酸差异位点是E2-5(H→D)、E2-109(G→D)、E2-368(V→A)和E2-374(C→G)。

图2以E基因为基础的系统进化分析
Fig.2PhylogeneticanalysisofJEVisolatesbasedonEgenesequence
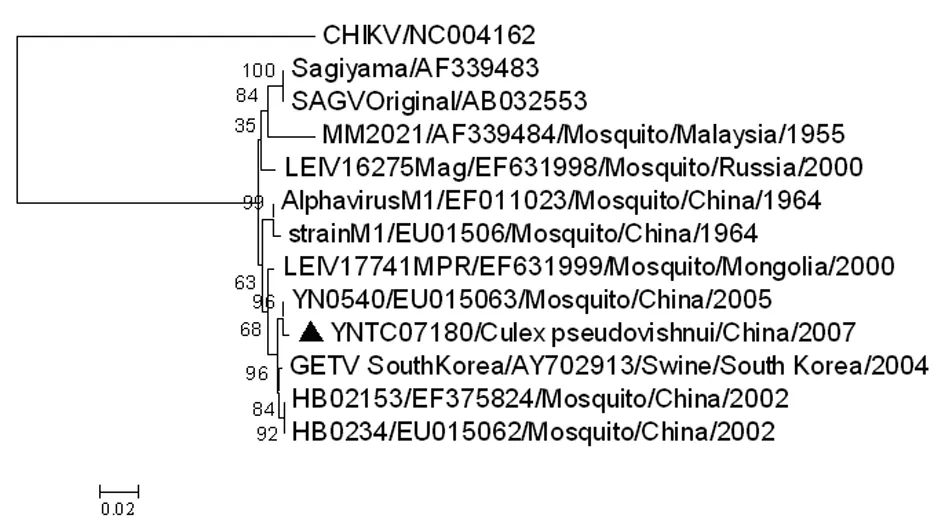
图3云南省GETV分离株Capsid基因的系统进化分析
Fig.3PhylogeneticanalysisofGETVisolatesbasedonCapsidgenesequence
3 讨 论
近10多年,先后在云南省分离到10余种虫媒病毒,其中流行性乙型脑炎是云南省分布最广,发病最多,危害最大的一种虫媒病毒病,并证实云南省JEV宿主动物和传播媒介种类多,分布广[6-8]。本次序列分析表明分离自腾冲县的20株JEV均为基因Ⅰ型。20世纪80和90年代,我们在云南省分离到的JEV几乎都是基因Ⅲ型[9-10],近几年相继在云南各地分离到基因Ⅰ型JEV[11-12]。本研究为首次证实云南省腾冲县存在基因Ⅰ型JEV流行,提示基因Ⅰ型已逐步取代基因Ⅲ型成为云南省JEV主要流行型,具有重要流行病学意义。
对新分离的20株JEV的PreM和E基因核苷酸和氨基酸同源性分析表明,它们之间仍然存在一定差异,尤其是PreM基因核苷酸差异度范围较大,如YNTC07028株与其它病毒株差异度高达9.2%,提示当地存在不同株系的JEV。根据图1和图2系统进化树分析,无论PreM或E基因均可将本次分离的20株JEV区分为两个进化分支,表明当地存在两个株系的JEV流行。这20株JEV与我国和东南亚国家近几年分离的基因Ⅰ型JEV相比,虽然它们间的同源性较高,但仍存在一定差异,而这些差异可能与不同地域JEV流行株为了适应当地的环境和媒介生物,导致病毒在进化中发生一些细微的改变。综合以上分析,我们认为,云南JEV与其它省份和国家流行株间虽有一些差异,但它们的抗原和毒力关键位点未发生明显变异,表明自然界JEV具有较好的遗传稳定性。
GETV是披膜病毒科(Togaviridae)甲病毒属(Alphavirus)塞姆利基森林病毒(Semliki forest virus, SFV)复合组中的一个成员,于1955年首次在马来西亚蚊虫中分离到,主要分布在亚洲及澳大利亚北部地区[13-14]。我国于1964年在海南首次分离到GETV(M1株),时隔38年后又在我国北方的河北省分离到两株,随后相继在我国其它省份分离到该病毒。目前在我国河北、上海、云南、辽宁和甘肃等省市共分离到10余株GETV[2]。提示该病毒在我国分布较广。GETV在人群中的感染和致病情况尚不清楚,但国外研究已证实,GETV是马和猪等家畜疾病的重要病原体[13-14]。因此,我国应加强GETV的分布及其与动物疾病关系的研究,深入开展我国GETV分离株与东南亚国家分离株的分子遗传研究,评价它们的公共卫生意义是当前面临的重要任务。
参考文献:
[1]Wang HY, Takasaki, T, Fu SH, et al. Molecular epidemiological analysis of Japanese encephalitis virus in China[J]. J Gen Virol, 2007, 88(3): 885-894.
[2]Zhai YG, Wang HY, Sun XH, et al. Complete sequence characterization of isolates of Getah virus (genus Alphavirus, family Togaviridae) from China[J]. J Gen Virol, 2008, 89(6): 1446-1456.
[3]Nga PT, Del Carmen Parquet M, Cuong VD, et al . Shift in Japanese encephalitis virus (JEV) genotype circulating in northern Vietnam: implications for frequent introductions of JEV from Southeast Asia to East Asia[J]. J Gen Virol, 2004, 85(6): 1625-1631.
[4]Ma SP, Yoshida Y, Makino Y, et al. Short report: a major genotype of Japanese encephalitis virus currently circulating in Japan[J]. Am J Trop Med Hyg, 2003, 69(2): 151-154.
[5]Jia LL, Yu YX, Huang Y, et al. Stability of phenotype and E protein gene of primary virus seed and final product of live attenuated Japanese encephalitis vaccine[J]. Chin J Biologicals, 2004, 17(1): 12-15. (in Chinese)
贾丽丽, 俞永新, 黄莺, 等. 流行性乙型脑炎减毒活疫苗生产毒种(SA14-14-2株)及其疫苗的表型和E基因稳定性研究[J]. 中国生物制品学杂志,2004, 17(1): 12-15.
[6]Zhang HL, Zi DY, Gong ZD, et al. Host and vector of Japanese encephalitis virus in Yunnan Province, China[J]. Chin J Prev Vet Med, 2000, 22(2): 81-83. (in Chinese)
张海林,自登云,龚正达,等. 云南省乙型脑炎病毒宿主和媒介研究[J]. 中国预防兽医学报,2000, 22(2): 81-83.
[7]Zhang HL, Shi HF, Mi ZQ, et al. The character of mosquitoes distribution and relation with the Arbovirus in Xishuangbanna Prefecture, Yunnan Province[J]. Endem Dis Bull, 2000, 15(4): 35-38. (in Chinese)
张海林,施华芳,米竹青,等. 云南省西双版纳州蚊虫分布特点及与虫媒病毒的关系[J]. 地方病通报,2000, 15(4):35-38.
[8]Zhang HL, Zi DY, Shi HF, et al. The natural infection rate of mosquitoes by Japanese encephalitis B virus in Yunnan Province[J]. Chin J Prev Med, 1990, 24(5): 265-267. (in Chinese)
张海林,自登云,施华芳,等. 云南省自然蚊体乙型脑炎病毒带毒率研究[J]. 中华预防医学杂志,1990, 24(5): 265-267.
[9]Wang JL, Zhang HL, Zhou JH, et al. Genotyping of Japanese encephalitis virus isolated in Yunnan[J]. Chin J Exper Clin Virol, 2008, 22(2): 87-90. (in Chinese)
王静林,张海林,周济华,等. 云南省乙型脑炎病毒基因分型研究[J].中华实验临床病毒学杂志,2008, 22(2):87-90.
[10]Sun YJ, Zhang HL. Distribution and gene-typing of Japanese encephalitis virus in China[J]. Chin J Vector Biol Ctrl, 2012, 23(5): 436-439. (in Chinese)
孙玉杰,张海林. 中国流行性乙型脑炎病毒基因型及分布[J]. 中国媒介生物学及控制杂志,2012, 23(5):436-439.
[11]Feng Y, Zhang HL, Fu SH, et al. Full-genome sequencing analysis of two stains of Japanese encephalitis virus isolated from Yunnan, China[J]. Chin J Viral Dis, 2012, 2(1): 9-15. (in Chinese)
冯云,张海林,付士红,等. 云南新分离2株基因I型流行性乙型脑炎病毒的全基因组序列分析[J]. 中国病毒病杂志,2012, 2(1):9-15.
[12]Yang DJ, Li MH, Fu SH, et al. Molecular characteristics of the full-length genomes of Japanese encephalitis virus strains newly isolated in 2009, China[J]. Chin J Virol, 2011, 27(6): 571-579. (in Chinese)
杨杜鹃,李铭华,付士红,等. 我国2009年新分离乙脑病毒全基因组序列特征研究[J]. 病毒学报,2011, 27(6):571-579.
[13]Fukunaqa Y, Kumanomido T, Kamada M. Getah virus as equine pathogen[J]. Vet Clin North Am Equine Pract, 2000, 16(3):605-617.
[14]Sun J, Zhang HL. The situation of research on Getah virus[J]. Chin J Zoonoses, 2012, 28(1): 77-79. (in Chinese)
孙洁,张海林. 盖塔病毒研究状况[J]. 中国人兽共患病学报,2012, 28(1):77-79.
猜你喜欢
小学生学习指导(爆笑校园)(2022年9期)2022-09-30
传染病信息(2022年3期)2022-07-15
保健医苑(2022年6期)2022-07-08
少男少女·小作家(2022年4期)2022-04-23
今日农业(2021年12期)2021-11-28
世界科学技术-中医药现代化(2021年5期)2021-11-05
杂文月刊(2019年14期)2019-08-03
祝您健康·文摘版(2018年4期)2018-05-14
广东第二课堂·小学(2016年9期)2016-05-14
中国当代医药(2015年31期)2015-03-01
